
Paclitaxel and Caffeine–Taurine, New Colchicine Alternatives for Chromosomes Doubling in Maize Haploid Breeding
 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
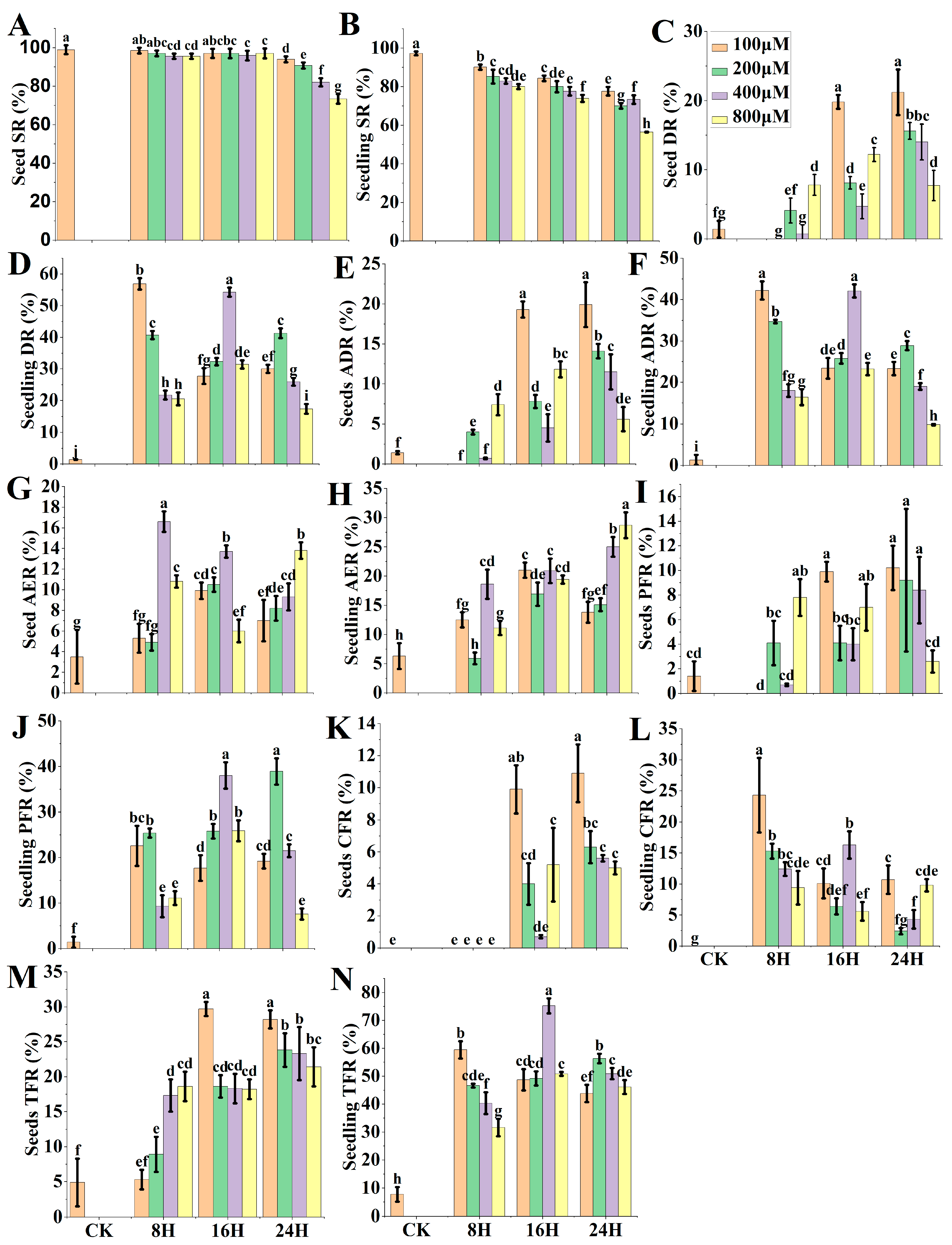
2.1. Experiment 1
Field Assessment-Based Comparative Efficiencies in Different Treatments of PTX Application
2.2. Experiment 2
2.2.1. Field Assessment-Based Comparative Efficiencies in Different Treatments of CAF-T Application
2.2.2. Field Assessment-Based Comparative Efficiencies in Different Treatments of Colchicine Application
2.3. Comparative DH Seed Quantity Produced by PTX, CAF-T versus Colchicine
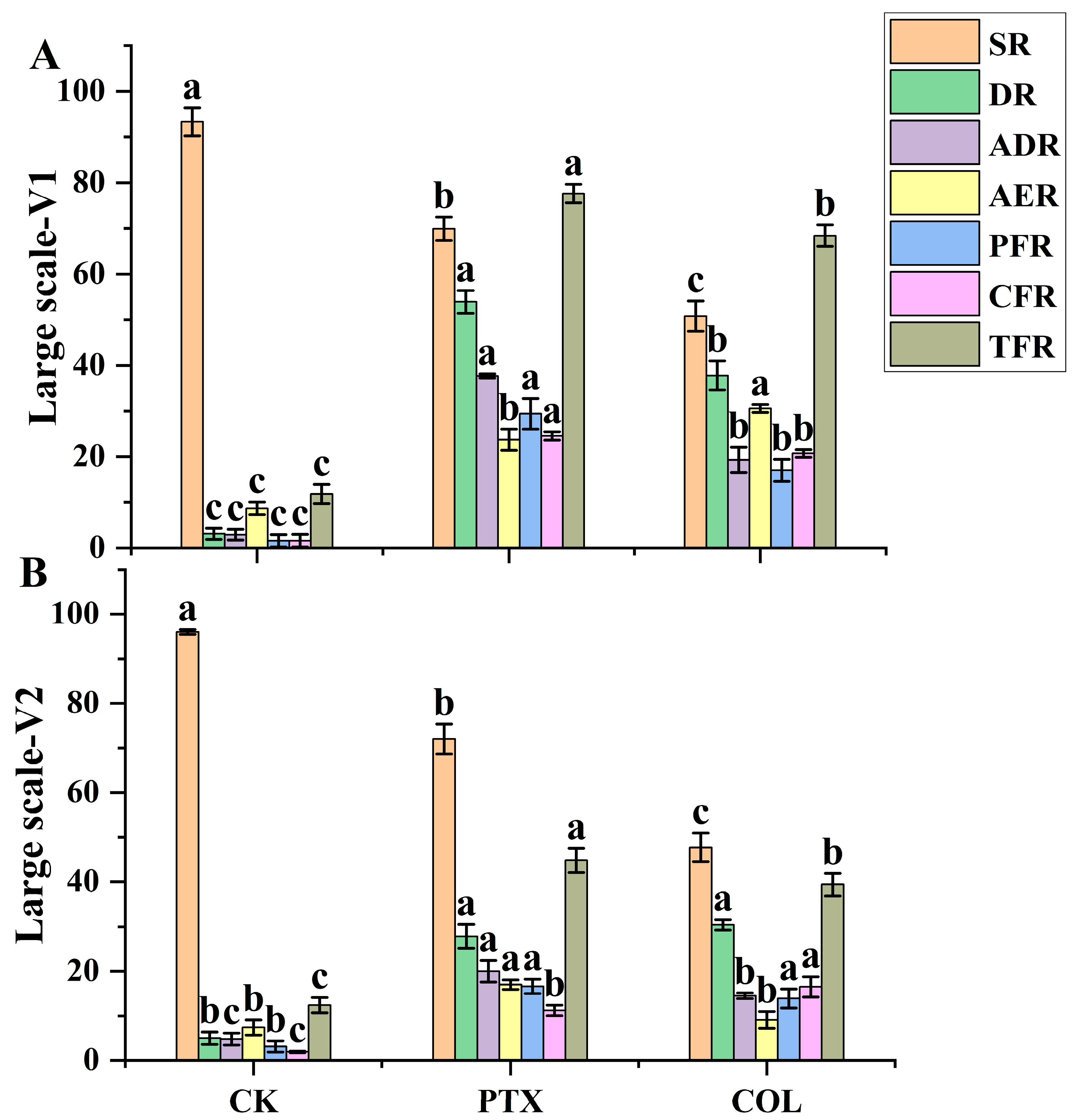
2.4. Experiment 4
Large-Scale (LS) Field Efficacy-Based Comparative Studies for Validation of the Best Treatment of PTX for DH Production Pipeline
2.5. Experiment 5
2.5.1. Morphological Studies
Comparative Plant and Ear Height of Treated Maize Plants by PTX versus Colchicine and CK
Comparative Silks Number/Ear Treated by PTX versus Colchicine and CK
Comparative Plant Weight and Root Growth Treated by PTX versus Colchicine and CK
2.5.2. Physiological Studies
Comparative Photosynthetic Pigments of Treated Seedlings by PTX versus Colchicine and CK
Comparative Germination (%) Impact on Treated Seeds by PTX, CAF–T versus Colchicine
- Germination (%) Impact on Treated Seeds by PTX
- 2.
- Germination (%) Impact on Treated Seeds by CAF–T
- 3.
- Germination (%) Impact on Treated Seeds by Colchicine
2.6. Experiment 6
2.6.1. PTX Induced Chromosome Doubling Signals Detected by Fluorescence in Situ Hybridization (FISH) Using Knob-2 as Probe
PTX Induced Chromosome Doubling by Seed-Soaking Method (M1)
PTX-Induced Chromosome Doubling by Root-Immersion Method (M3)
2.7. Experiment 7
2.7.1. CAF–T-Induced Chromosome-Doubling Signals Detected by FISH Using Knob-2 as Probe
CAF–T-Induced Chromosome Doubling by Seed-Soaking Method (M1)
CAF–T-Induced Chromosome Doubling by Root-Immersion Method (M3)
3. Discussion
4. Materials and Methods
4.1. Germplasm, Chemicals, Instruments and Experimental Locations
4.2. Treatments & Concentrations
4.2.1. Experiment 1
4.2.2. Experiment 2
4.2.3. Experiment 3
4.2.4. Experiments 4 and 5
4.2.5. Experiments 6 and 7
4.3. Methods
4.3.1. Seed-Soaking (Method 1/M1)
4.3.2. Seedling-Immersion (Method 2/M2)
4.3.3. Root-Immersion (Method 3/M3)
4.3.4. Acclimatization of Seedlings in Greenhouse and Transplanting in Field
4.3.5. Experiments 6 and 7
4.4. Assessment Methodology and Data Collection Procedure
4.4.1. Experiments 1–4
- Survival rate (SR) = count of survived plants at pollination/number of seedlings treated × 100 [46];
- Doubling rate (DR) = sum of plants showed partial and/or complete fertility/count of haploid survived plants at pollination × 100;
- Actual doubling rate (ADR) = SR × DR/100 [24];
- Partial fertility rate (PFR) = number of plants that exhibited partial fertility/number of haploid plants that survived at pollination × 100;
- Complete fertility rate (CFR) = number of plants presented complete fertility/number of haploid plants that survived at pollination ×100;
- Total fertility rate (TFR) = AER + PFR + CFR.
- Percentage of DH lines with 1–5 seeds = number of D0 plants/D1 ears produced 1–5 seeds/number of haploids survived plants at pollination × 100;
- Percentage of DH lines containing 6–25 seeds = number of D0 plants/D1 ears produced 6–25 seeds/number of haploid survived plants at pollination × 100;
- Percentage of DH lines containing > 25 seeds = number of D0 plants/D1 ears produced > 25 seeds/number of haploid survived plants at pollination × 100;
- RR = number of D0 plants produced seeds/number of D0 survived haploid plants at pollination × 100;
- OSR = number of D0 plants produced seeds/number of putative seeds/seedlings treated with chemical agent × 100.
4.4.2. Experiment 5
Morphological Studies
- Plant and Ear Height, Plant Weight, and Number of Silks/Ear
Physiological Studies
- Measurement of Photosynthetic Pigments (Chla, Chlb, ChlT, and Cx)
- 2.
- Germination Percentages
4.4.3. Experiments 6 and 7
4.5. Statistical Analysis and Graphics Improvement
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hallauer, A.R.; Carena, M.J.; Miranda Filho, J.B.D. Quantitative Genetics in Maize Breeding; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010; Volume 6. [Google Scholar]
- Jacquier, N.M.A.; Gilles, L.M.; Martinant, J.-P.; Rogowsky, P.M.; Widiez, T. Maize in planta haploid inducer lines: A cornerstone for doubled haploid technology. In Doubled Haploid Technol.; Humana: New York, NY, USA, 2021; pp. 25–48. [Google Scholar]
- Chaikam, V.; Prasanna, B.M. Doubled Haploid Technology for Rapid and Efficient Maize Breeding. In Accelerated Plant Breeding, Volume 1: Cereal Crops; Springer: Berlin/Heidelberg, Germany, 2020; pp. 257–292. [Google Scholar]
- Yan, G.; Liu, H.; Wang, H.; Lu, Z.; Wang, Y.; Mullan, D.; Hamblin, J.; Liu, C. Accelerated generation of selfed pure line plants for gene identification and crop breeding. Front. Plant Sci. 2017, 8, 1786. [Google Scholar] [CrossRef]
- Chaikam, V.; Gowda, M.; Martinez, L.; Ochieng, J.; Omar, H.A.; Prasanna, B.M. Improving the efficiency of colchicine-based chromosomal doubling of maize haploids. Plants 2020, 9, 459. [Google Scholar] [CrossRef]
- Böhm, J.; Schipprack, W.; Utz, H.F.; Melchinger, A.E. Tapping the genetic diversity of landraces in allogamous crops with doubled haploid lines: A case study from European flint maize. Theor. Appl. Genet. 2017, 130, 861–873. [Google Scholar] [CrossRef]
- Brauner, P.C.; Schipprack, W.; Utz, H.F.; Bauer, E.; Mayer, M.; Schön, C.-C.; Melchinger, A.E. Testcross performance of doubled haploid lines from European flint maize landraces is promising for broadening the genetic base of elite germplasm. Theor. Appl. Genet. 2019, 132, 1897–1908. [Google Scholar] [CrossRef]
- Melchinger, A.E.; Böhm, J.; Utz, H.F.; Müller, J.; Munder, S.; Mauch, F.J. High-throughput precision phenotyping of the oil content of single seeds of various oilseed crops. Crop Sci. 2018, 58, 670–678. [Google Scholar] [CrossRef]
- Kanbar, O.Z.; Lantos, C.; Pauk, J. In vitro anther culture as efficiently applied technique for doubled haploid production of wheat (Triticum aestivum L.). Ratar. I Povrt. 2021, 58, 31–45. [Google Scholar] [CrossRef]
- Ravi, M.; Chan, S.W.L. Haploid plants produced by centromere-mediated genome elimination. Nature 2010, 464, 615–618. [Google Scholar] [CrossRef]
- Sorrells, M.E.; Gustafson, J.P.; Somers, D.; Chao, S.; Benscher, D.; Guedira-Brown, G.; Huttner, E.; Kilian, A.; McGuire, P.E.; Ross, K. Reconstruction of the Synthetic W7984× Opata M85 wheat reference population. Genome 2011, 54, 875–882. [Google Scholar] [CrossRef]
- Murovec, J.; Bohanec, B. Haploids and doubled haploids in plant breeding. In Plant Breeding; Abdurakhmonov, I., Ed.; InTech Europe: Rijeka, Croatia, 2012; pp. 87–106. ISBN 978-953-307-932-5. [Google Scholar]
- Hao, M.; Chen, J.; Zhang, L.; Luo, J.; Yuan, Z.; Yan, Z.; Zhang, B.; Chen, W.; Wei, Y.; Zhang, H. The genetic study utility of a hexaploid wheat DH population with non-recombinant A-and B-genomes. SpringerPlus 2013, 2, 131. [Google Scholar] [CrossRef]
- Shi, Y.G.; Lian, Y.; Shi, H.W.; Wang, S.G.; Fan, H.; Sun, D.Z.; Jing, R.L. Dynamic analysis of QTLs for green leaf area duration and green leaf number of main stem in wheat. Cereal Res. Commun. 2019, 47, 250–263. [Google Scholar] [CrossRef]
- Dunwell, J.M. Haploids in flowering plants: Origins and exploitation. Plant Biotechnol. J. 2010, 8, 377–424. [Google Scholar] [CrossRef]
- Chaikam, V.; Mahuku, G.; Prasanna, B.M. Chromosome Doubling of Maternal Haploids; CIMMYT: Texcoco, Mexico, 2012. [Google Scholar]
- Dhooghe, E.; Van Laere, K.; Eeckhaut, T.; Leus, L.; Van Huylenbroeck, J. Mitotic chromosome doubling of plant tissues in vitro. Plant Cell Tissue Organ Cult. (PCTOC) 2011, 104, 359–373. [Google Scholar] [CrossRef]
- Chase, S.S. Monoploids and monoploid-derivatives of maize (Zea mays L.). Bot. Rev. 1969, 35, 117–168. [Google Scholar] [CrossRef]
- Deimling, S.; Röber, F.K.; Geiger, H.H. Methodology and genetics of in vivo haploid induction in maize. Vortr. Pflanzenzüchtg 1997, 38, 203–224. [Google Scholar]
- Wan, Y.; Duncan, D.R.; Rayburn, A.L.; Petolino, J.F.; Widholm, J.M. The use of antimicrotubule herbicides for the production of doubled haploid plants from anther-derived maize callus. Theor. Appl. Genet. 1991, 81, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Melchinger, A.E.; Brauner, P.C.; Böhm, J.; Schipprack, W. In vivo haploid induction in maize: Comparison of different testing regimes for measuring haploid induction rates. Crop Sci. 2016, 56, 1127–1135. [Google Scholar] [CrossRef]
- Chaikam, V.; Molenaar, W.; Melchinger, A.E.; Boddupalli, P.M. Doubled haploid technology for line development in maize: Technical advances and prospects. Theor. Appl. Genet. 2019, 132, 3227–3243. [Google Scholar] [CrossRef]
- Duncan, D.R.; Widholm, J.M. Differential response to potassium permanganate of regenerable and of non-regenerable tissue cell walls from maize callus cultures. Plant Sci. 1989, 61, 91–103. [Google Scholar] [CrossRef]
- Ren, X. Doubling effect of anti-microtubule herbicides on the maize haploid. Emir. J. Food Agric. 2018, 30, 903–908. [Google Scholar] [CrossRef]
- Molenaar, W.S.; Schipprack, W.; Melchinger, A.E. Nitrous Oxide-Induced Chromosome Doubling of Maize Haploids. Crop Sci. 2018, 58, 650–659. [Google Scholar] [CrossRef]
- Kato, A.; Geiger, H.H. Chromosome doubling of haploid maize seedlings using nitrous oxide gas at the flower primordial stage. Plant Breed. 2002, 121, 370–377. [Google Scholar] [CrossRef]
- Morejohn, L.C.; Bureau, T.E.; Tocchi, L.P.; Fosket, D.E. Resistance of Rosa microtubule polymerization to colchicine results from a low-affinity interaction of colchicine and tubulin. Planta 1987, 170, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Morejohn, L.C.; Bureau, T.E.; Tocchi, L.P.; Fosket, D.E. Tubulins from different higher plant species are immunologically nonidentical and bind colchicine differentially. Proc. Natl. Acad. Sci. USA 1984, 81, 1440–1444. [Google Scholar] [CrossRef] [PubMed]
- Khanna, C.; Rosenberg, M.; Vail, D.M. A review of paclitaxel and novel formulations including those suitable for use in dogs. J. Vet. Intern. Med. 2015, 29, 1006–1012. [Google Scholar] [CrossRef] [PubMed]
- Fong, K.-W.; Leung, J.W.-C.; Li, Y.; Wang, W.; Feng, L.; Ma, W.; Liu, D.; Songyang, Z.; Chen, J. MTR120/KIAA1383, a novel microtubule-associated protein, promotes microtubule stability and ensures cytokinesis. J. Cell Sci. 2013, 126, 825–837. [Google Scholar] [CrossRef]
- Snyder, J.P.; Nettles, J.H.; Cornett, B.; Downing, K.H.; Nogales, E. The binding conformation of Taxol in β-tubulin: A model based on electron crystallographic density. Proc. Natl. Acad. Sci. USA 2001, 98, 5312–5316. [Google Scholar] [CrossRef]
- Yan-Hua, Y.; Jia-Wang, M.A.O.; Xiao-Li, T.A.N. Research progress on the source, production, and anti-cancer mechanisms of paclitaxel. Chin. J. Nat. Med. 2020, 18, 890–897. [Google Scholar]
- Lourenco, R.; Camilo, M.E. Taurine: A conditionally essential amino acid in humans? An overview in health and disease. Nutr Hosp 2002, 17, 262–270. [Google Scholar]
- Mates, J.M.; Segura, J.A.; Alonso, F.J.; Marquez, J. Sulphur-containing non enzymatic antioxidants: Therapeutic tools against cancer. Front. Biosci.-Sch. 2012, 4, 722–748. [Google Scholar] [CrossRef]
- Kilb, W.; Fukuda, A. Taurine as an essential neuromodulator during perinatal cortical development. Front. Cell. Neurosci. 2017, 11, 328. [Google Scholar] [CrossRef]
- Morales-Borge, R.H.; González, M.J.; Gupta, R.C.; Ayeotan, O. Taurine as Anticancer and Antiviral: Case Report and Prospective Update. Cancer Case Rep. 2020, 1, 1–14. [Google Scholar] [CrossRef]
- He, F.; Ma, N.; Midorikawa, K.; Hiraku, Y.; Oikawa, S.; Mo, Y.; Zhang, Z.; Takeuchi, K.; Murata, M. Anti-cancer mechanisms of taurine in human nasopharyngeal carcinoma cells. In Taurine 11; Springer: Berlin/Heidelberg, Germany, 2019; pp. 533–541. [Google Scholar]
- Yousuf, M.; Shamsi, A.; Mohammad, T.; Azum, N.; Alfaifi, S.Y.M.; Asiri, A.M.; Mohamed Elasbali, A.; Islam, A.; Hassan, M.I.; Haque, Q.M.R. Inhibiting Cyclin-Dependent Kinase 6 by Taurine: Implications in Anticancer Therapeutics. ACS Omega 2022, 7, 25844–25852. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Lu, R.; Huang, C.; Lin, D. Taurine protects C2C12 myoblasts from impaired cell proliferation and myotube differentiation under cisplatin-induced ROS exposure. Front. Mol. Biosci. 2021, 8, 685362. [Google Scholar] [CrossRef]
- Broughton, S.; Castello, M.; Liu, L.; Killen, J.; Hepworth, A.; O’Leary, R. The effect of caffeine and trifluralin on chromosome doubling in wheat anther culture. Plants 2020, 9, 105. [Google Scholar] [CrossRef]
- Yasuhara, H. Caffeine inhibits callose deposition in the cell plate and the depolymerization of microtubules in the central region of the phragmoplast. Plant Cell Physiol. 2005, 46, 1083–1092. [Google Scholar] [CrossRef] [PubMed]
- Prigge, V.; Melchinger, A.E. Production of haploids and doubled haploids in maize. In Plant cell culture protocols; Springer: Berlin/Heidelberg, Germany, 2012; pp. 161–172. [Google Scholar]
- Gayen, P.; Madan, J.K.; Kumar, R.; Sarkar, K.R. Chromosome doubling in haploids through colchicine. Maize Genet. Coop. Newsl. 1994, 68, 65. [Google Scholar]
- Eder, J.; Chalyk, S. In vivo haploid induction in maize. Theor. Appl. Genet. 2002, 104, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Chalyk, S.T. Obtaining fertile pollen in maize maternal haploids. Maize Genet. Coop. Newsl. 2000, 74, 17–18. [Google Scholar]
- Melchinger, A.E.; Molenaar, W.S.; Mirdita, V.; Schipprack, W. Colchicine alternatives for chromosome doubling in maize haploids for doubled-haploid production. Crop Sci. 2016, 56, 559–569. [Google Scholar] [CrossRef]
- Mahuku, G. Putative DH Seedlings: From the Lab to the Field; CIMMYT: Texcoco, Mexico, 2012. [Google Scholar]
- Kleiber, D.; Prigge, V.; Melchinger, A.E.; Burkard, F.; San Vicente, F.; Palomino, G.; Gordillo, G.A. Haploid fertility in temperate and tropical maize germplasm. Crop Sci. 2012, 52, 623–630. [Google Scholar] [CrossRef]
- Ren, J.; Wu, P.; Tian, X.; Lübberstedt, T.; Chen, S. QTL mapping for haploid male fertility by a segregation distortion method and fine mapping of a key QTL qhmf4 in maize. Theor. Appl. Genet. 2017, 130, 1349–1359. [Google Scholar] [CrossRef]
- Chaikam, V.; Gowda, M.; Nair, S.K.; Melchinger, A.E.; Boddupalli, P.M. Genome-wide association study to identify genomic regions influencing spontaneous fertility in maize haploids. Euphytica 2019, 215, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Ren, J.; Tian, X.; Lübberstedt, T.; Li, W.; Li, G.; Li, X.; Chen, S. New insights into the genetics of haploid male fertility in maize. Crop Sci. 2017, 57, 637–647. [Google Scholar] [CrossRef]
- Ma, H.; Li, G.; Würschum, T.; Zhang, Y.; Zheng, D.; Yang, X.; Li, J.; Liu, W.; Yan, J.; Chen, S. Genome-wide association study of haploid male fertility in maize (Zea mays L.). Front. Plant Sci. 2018, 9, 974. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, B.M.; Chaikam, V.; Mahuku, G. Doubled Haploid Technology in Maize Breeding: Theory and Practice; CIMMYT: Texcoco, Mexico, 2012. [Google Scholar]
- Ghotbi Ravandi, E.; Rezanejad, F.; Zolala, J.; Dehghan, E. The effects of chromosome-doubling on selected morphological and phytochemical characteristics of Cichorium intybus L. J. Hortic. Sci. Biotechnol. 2013, 88, 701–709. [Google Scholar] [CrossRef]
- Jensen, C. Chromosome Doubling Techniques in Haploids. In Haploids in Higher Plants: Advances and Potential; The University of Guelph: Guelph, ON, Canada, 1974; pp. 151–190. [Google Scholar]
- Inagaki, M.N. Doubled haploid production in wheat through wide hybridization. In Doubled Haploid Production in Crop Plants; Springer: Berlin/Heidelberg, Germany, 2003; pp. 53–58. [Google Scholar]
- Davarnejad, R.; Moraveji, M.K.; Havaie, M. Integral technique for evaluation and optimization of Ni (II) ions adsorption onto regenerated cellulose using response surface methodology. Arab. J. Chem. 2018, 11, 370–379. [Google Scholar] [CrossRef]
- Jiang, M.; Wu, H.; Li, Z.; Ji, D.; Li, W.; Liu, Y.; Yuan, D.; Wang, B.; Zhang, Z. Highly selective photoelectrochemical conversion of carbon dioxide to formic acid. ACS Sustain. Chem. Eng. 2018, 6, 82–87. [Google Scholar] [CrossRef]
- Khaleel, C.; Tabanca, N.; Buchbauer, G. α-Terpineol, a natural monoterpene: A review of its biological properties. Open Chem. 2018, 16, 349–361. [Google Scholar] [CrossRef]
- Sankarganesh, M.; Rajesh, J.; Kumar, G.G.V.; Vadivel, M.; Mitu, L.; Kumar, R.S.; Raja, J.D. Synthesis, spectral characterization, theoretical, antimicrobial, DNA interaction and in vitro anticancer studies of Cu(II) and Zn(II) complexes with pyrimidine-morpholine based Schiff base ligand. J. Saudi Chem. Soc. 2018, 22, 416–426. [Google Scholar] [CrossRef]
- Thomas, J.; Chen, Q.; Howes, N. Chromosome doubling of haploids of common wheat with caffeine. Genome 1997, 40, 552–558. [Google Scholar] [CrossRef]
- Chaudhary, H.K.; Badiyala, A.; Jamwal, N.S. New frontiers in doubled haploidy breeding in wheat. Agric. Res. J. 2015, 52, 1–12. [Google Scholar] [CrossRef]
- Tiwari, A.K.; Mishra, S.K. Effect of colchicine on mitotic polyploidization and morphological characteristics of Phlox drummondi. Afr. J. Biotechnol. 2012, 11, 9336–9342. [Google Scholar] [CrossRef]
- Kwon, S.-J.; Roy, S.K.; Cho, K.-Y.; Moon, Y.-J.; Woo, S.-H.; Kim, H.-H. Effect of Colchicine on the Induction of Prunella vulgaris for. albiflora Nakai. Korean J. Crop Sci. 2015, 60, 107–113. [Google Scholar] [CrossRef]
- Lv, Z.; Zhu, F.; Jin, D.; Wu, Y.; Wang, S. Seed Germination and Seedling Growth of Dendrocalumus brandisii in vitro, and the Inhibitory Mechanism of Colchicine. Front. Plant Sci. 2021, 12, 784581. [Google Scholar] [CrossRef] [PubMed]
- Megbo, B.C. The physiological effects of colchicine in Okra, Hibiscus esculentus L, plant growth and development. Int. J. Sci. Eng. Res. 2010, 1, 29–33. [Google Scholar]
- Amiri, S.; Kazemitabaar, S.K.; Ranjbar, G.; Azadbakht, M. The effect of trifluralin and colchicine treatments on morphological characteristics of jimsonweed (Datura stramonium L.). Trakia J. Sci. 2010, 8, 47–61. [Google Scholar]
- Li, Y.; He, N.; Hou, J.; Xu, L.; Liu, C.; Zhang, J.; Wang, Q.; Zhang, X.; Wu, X. Factors influencing leaf chlorophyll content in natural forests at the biome scale. Front. Ecol. Evol. 2018, 6, 64. [Google Scholar] [CrossRef]
- Cogdell, R.J. Carotenoids in photosynthesis. Philos. Trans. R. Soc. London. B Biol. Sci. 1978, 284, 569–579. [Google Scholar] [CrossRef]
- Zhu, M.; Du, P.; Zhuang, L.; Chu, C.; Zhao, H.; Qi, Z. A simple and efficient non-denaturing FISH method for maize chromosome differentiation using single-strand oligonucleotide probes. Genome 2017, 60, 657–664. [Google Scholar] [CrossRef]
- Han, Y.; Zhang, T.; Thammapichai, P.; Weng, Y.; Jiang, J. Chromosome-specific painting in Cucumis species using bulked oligonucleotides. Genetics 2015, 200, 771–779. [Google Scholar] [CrossRef]
- Surapaneni, M.S.; Das, S.K.; Das, N.G. Designing Paclitaxel drug delivery systems aimed at improved patient outcomes: Current status and challenges. Int. Sch. Res. Not. 2012, 2012, 623139. [Google Scholar] [CrossRef]
- Shanmugam, S.; Im, H.T.; Sohn, Y.T.; Kim, Y.-I.; Park, J.-H.; Park, E.-S.; Woo, J.S. Enhanced oral bioavailability of paclitaxel by solid dispersion granulation. Drug Dev. Ind. Pharm. 2015, 41, 1864–1876. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Z. Development and Characterization of Small Segmental Translocations of Thinopyrum Bessarabicum and Cytological Mapping of Interesting Genes. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2013. [Google Scholar]
- Su, S.; Zhou, Y.; Qin, J.G.; Yao, W.; Ma, Z. Optimization of the method for chlorophyll extraction in aquatic plants. J. Freshw. Ecol. 2010, 25, 531–538. [Google Scholar] [CrossRef]
- Webb, D.J.; Burnison, B.K.; Trimbee, A.M.; Prepas, E.E. Comparison of chlorophyll a extractions with ethanol and dimethyl sulfoxide/acetone, and a concern about spectrophotometric phaeopigment correction. Can. J. Fish. Aquat. Sci. 1992, 49, 2331–2336. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arshad, S.; Wei, M.; Ali, Q.; Mustafa, G.; Ma, Z.; Yan, Y. Paclitaxel and Caffeine–Taurine, New Colchicine Alternatives for Chromosomes Doubling in Maize Haploid Breeding. Int. J. Mol. Sci. 2023, 24, 14659. https://doi.org/10.3390/ijms241914659
Arshad S, Wei M, Ali Q, Mustafa G, Ma Z, Yan Y. Paclitaxel and Caffeine–Taurine, New Colchicine Alternatives for Chromosomes Doubling in Maize Haploid Breeding. International Journal of Molecular Sciences. 2023; 24(19):14659. https://doi.org/10.3390/ijms241914659
Chicago/Turabian StyleArshad, Saeed, Mengli Wei, Qurban Ali, Ghulam Mustafa, Zhengqiang Ma, and Yuanxin Yan. 2023. "Paclitaxel and Caffeine–Taurine, New Colchicine Alternatives for Chromosomes Doubling in Maize Haploid Breeding" International Journal of Molecular Sciences 24, no. 19: 14659. https://doi.org/10.3390/ijms241914659