
Integrated Secondary Metabolomic and Antioxidant Ability Analysis Reveals the Accumulation Patterns of Metabolites in Momordica charantia L. of Different Cultivars
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Comparison of Characters and Nutrients of Different Cultivars
2.2. Organic Acid Content
2.3. Total Phenolic, Flavonoid, and Triterpenoids Contents
2.4. Antioxidant Activity
2.5. Correlation of TPC, TFC, and TTC with Antioxidative Capacity
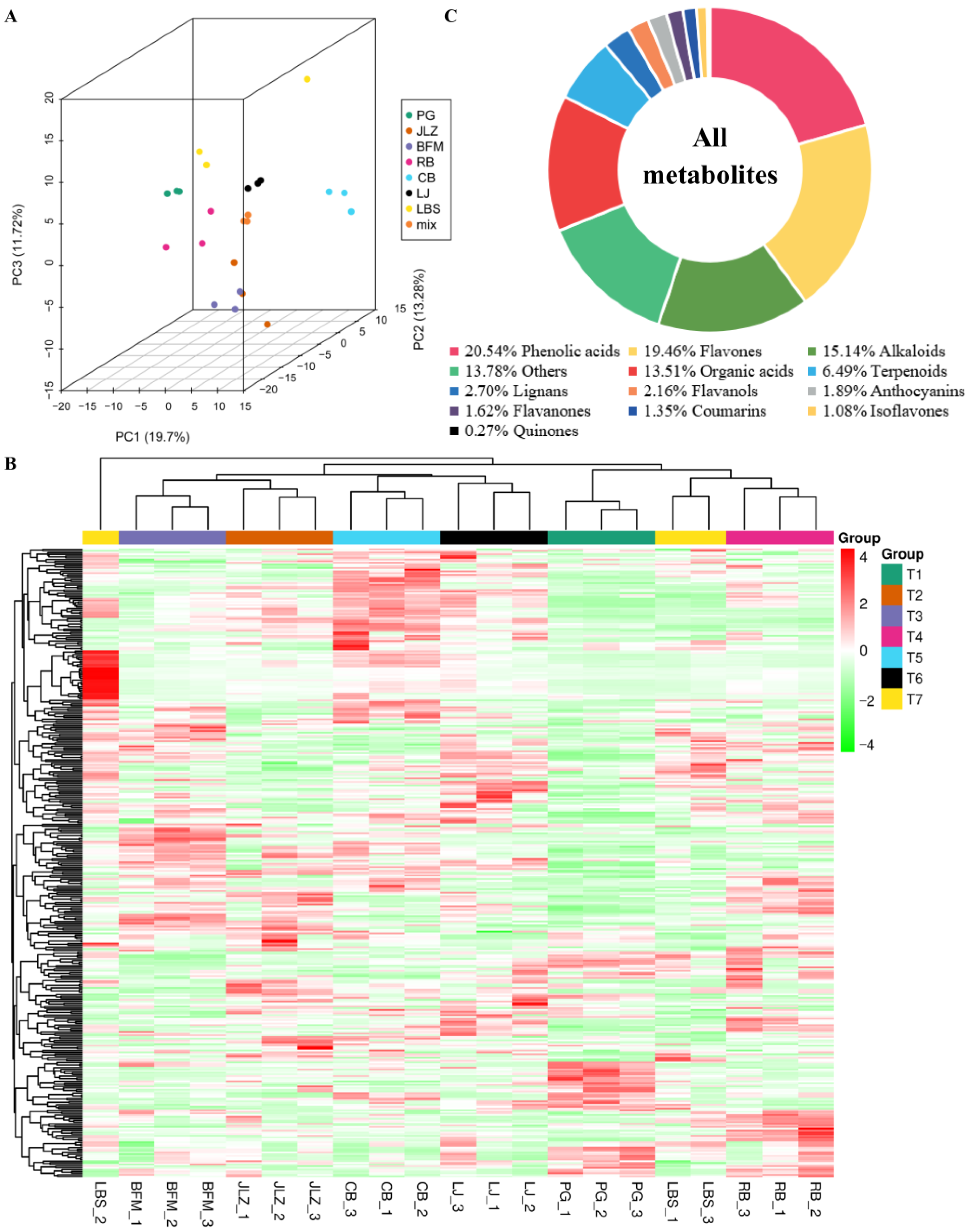
2.6. Profiles of the Secondary Metabolome in Seven Cultivars of Bitter Gourd
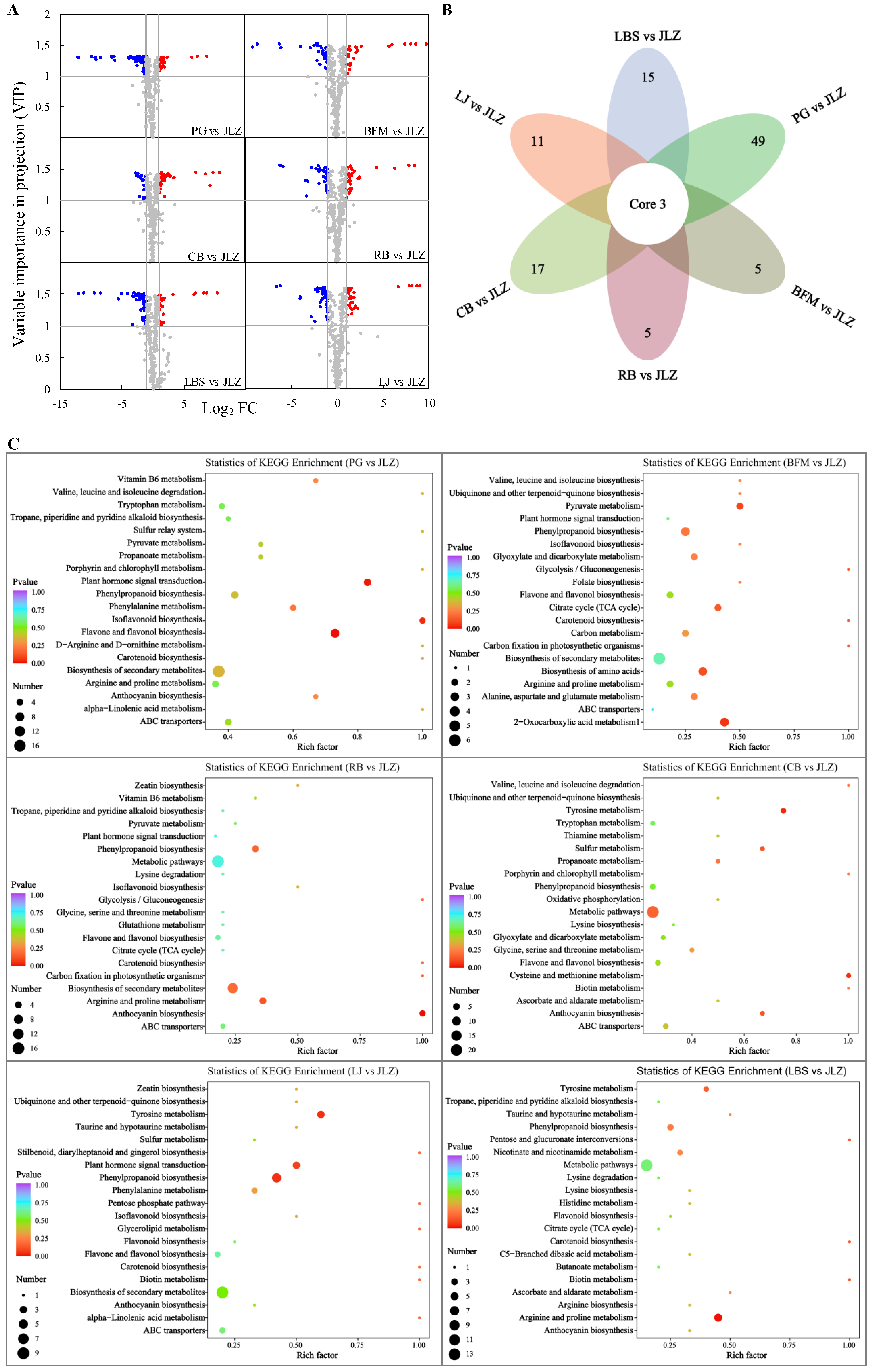
2.7. Differential Metabolite Analysis of Seven Cultivars of Bitter Gourd
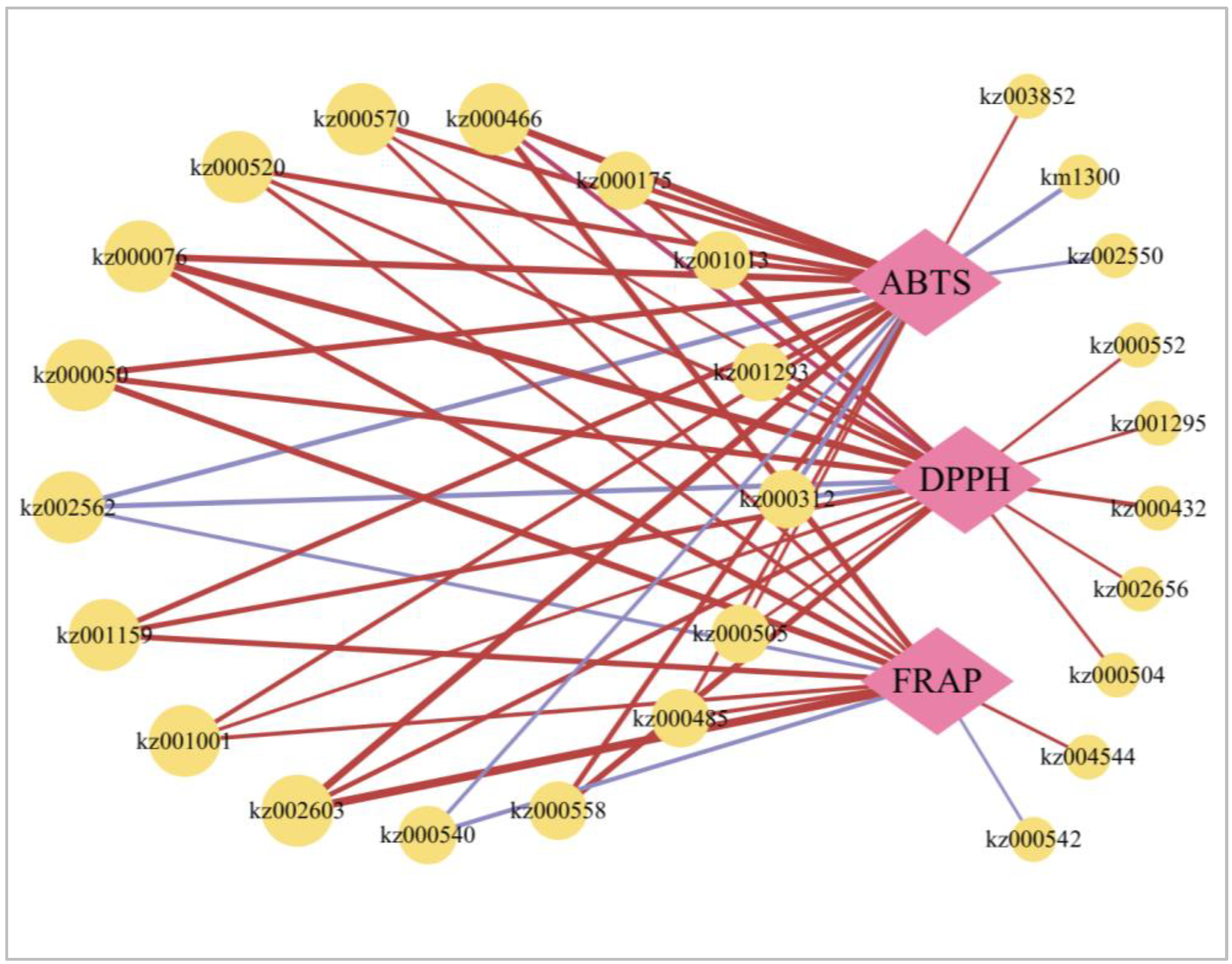
2.8. Network Diagram between Antioxidative Capacity and Metabolites
3. Discussion
3.1. Comparison of Antioxidant Activity in Different Cultivars of Bitter Gourd
3.2. Phenolic Acids and Flavonoid Components
3.3. Terpenoids Components
3.4. Organic Acids Components
3.5. Other Components
4. Materials and Methods
4.1. Plant Cultivars
4.2. Determination of Nutrient Composition Content
4.3. Quantification of Organic Acid Content
4.4. Total Flavonoid, Triterpenoid, and Phenolic Content Quantification
4.5. Determination of Antioxidative Activity
4.6. UPLC-MS/MS and Secondary Metabolomics Analysis
4.7. KEGG Annotation and Enrichment Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cortez-Navarrete, M.; Méndez-Del Villar, M.; Ramos-González, E.J.; Pérez-Rubio, K.G. Momordica charantia: A review of its effects on metabolic diseases and mechanisms of action. J. Med. Food 2021, 24, 1017–1027. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Kim, E.K.; Choi, Y.J.; Tang, Y.; Moon, S.H. The role of Momordica charantia in resisting obesity. Int. J. Environ. Res. Public Health 2019, 16, 3251. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, Q.; Zeng, T.; Zhan, J.; Zhao, H.; Ho, C.T.; Xiao, Y.; Li, S. Immunomodulatory effects and associated mechanisms of Momordica charantia and its phytochemicals. Food Funct. 2022, 13, 11986–11998. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Du, X.; Yu, J.; Jin, H.; Liu, N. Comparative metabolomics study of flavonoids in the pericarp of different coloured bitter gourds (Momordica charantia L.). Physiol. Mol. Biol. Plants Int. J. Funct. Plant Biol. 2022, 28, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.P.; Kha, T.C.; Parks, S.E.; Roach, P.D. Bitter melon (Momordica charantia L.) bioactive composition and health benefits: A review. Plants 2016, 32, 181–202. [Google Scholar]
- Aminah, A.; Anna, P.K. Influence of ripening stages on physicochemical characteristics and antioxidant properties of bitter gourd (Monodic charlatan). Int. Food Res. J. 2011, 18, 895–900. [Google Scholar]
- Habicht, S.D.; Kind, V.; Rudloff, S.; Borsch, C.; Mueller, A.S.; Pallauf, J.; Yang, R.Y.; Krawinkel, M.B. Quantification of antidiabetic extracts and compounds in bitter gourd varieties. Food Chem. 2011, 126, 172–176. [Google Scholar] [CrossRef]
- Bora, A.F.M.; Kouame, K.J.E.; Li, X.; Liu, L.; Pan, Y. New insights into the bioactive polysaccharides, proteins, and triterpenoids isolated from bitter melon (Momordica charantia) and their relevance for nutraceutical and food application: A review. Int. J. Biol. Macromol. 2023, 231, 123173. [Google Scholar] [CrossRef]
- Wang, S.; Li, Z.; Yang, G.; Ho, C.T.; Li, S. Momordica charantia: A popular health-promoting vegetable with multifunctionality. Food Funct. 2017, 8, 1749–1762. [Google Scholar] [CrossRef]
- Krawinkel, M.B.; Keding, G.B. Bitter gourd (Momordica charantia): A dietary approach to hyperglycemia. Nutr. Rev. 2006, 64 Pt 1, 331–337. [Google Scholar] [CrossRef]
- Fernie, A.R.; Pichersky, E. Focus issue on metabolism: Metabolites, metabolites everywhere. Plant Physiol. 2015, 169, 1421–1423. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Kliebenstein, D.J. Plant secondary metabolites as defenses, regulators, and primary metabolites: The blurred functional trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Abiri, R.; Abdul-Hamid, H.; Sytar, O.; Abiri, R.; Bezerra de Almeida, E., Jr.; Sharma, S.K.; Bulgakov, V.P.; Arroo, R.R.J.; Malik, S. A brief overview of potential treatments for viral diseases using natural plant compounds: The case of SARS-CoV. Molecules 2021, 26, 3868. [Google Scholar] [CrossRef]
- Chou, M.C.; Lee, Y.J.; Wang, Y.T.; Cheng, S.Y.; Cheng, H.L. Cytotoxic and anti-inflammatory triterpenoids in the vines and leaves of Momordica charantia. Int. J. Mol. Sci. 2022, 23, 1071. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, R.P.; Koprivova, A.; Kopriva, S. Pinpointing secondary metabolites that shape the composition and function of the plant microbiome. J. Exp. Bot. 2021, 72, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Ankit; Sharma, R.; Gogna, N.; Dorai, K. NMR-based metabolomic profiling of the differential concentration of phytomedicinal compounds in pericarp, skin and seeds of Momordica charantia (bitter melon). Nat. Prod. Res. 2022, 36, 390–395. [Google Scholar] [CrossRef]
- Niu, Y.; Liu, Z.; He, H.; Han, X.; Qi, Z.; Yang, Y. Gene expression and metabolic changes of Momordica charantia L. seedlings in response to low temperature stress. PLoS ONE 2020, 15, e0233130. [Google Scholar] [CrossRef]
- Wang, S.Y.; Ballington, J. Free radical scavenging capacity and antioxidant enzyme activity in deerberry (Vaccinium stamineum L.): Food science + technology. Sci. Technol. Aliment. 2007, 40, 1352–1361. [Google Scholar]
- Pérez-Torres, I.; Castrejón-Téllez, V.; Soto, M.E.; Rubio-Ruiz, M.E.; Manzano-Pech, L.; Guarner-Lans, V. Oxidative stress, plant natural antioxidants, and obesity. Int. J. Mol. Sci. 2021, 22, 1786. [Google Scholar] [CrossRef]
- Matsumura, H.; Hsiao, M.C.; Lin, Y.P.; Toyoda, A.; Taniai, N.; Tarora, K.; Urasaki, N.; Anand, S.S.; Dhillon, N.P.S.; Schafleitner, R.; et al. Long-read bitter gourd (Momordica charantia) genome and the genomic architecture of nonclassic domestication. Proc. Natl. Acad. Sci. USA 2020, 117, 14543–14551. [Google Scholar] [CrossRef]
- Chen, F.; Huang, G.; Yang, Z.; Hou, Y. Antioxidant activity of Momordica charantia polysaccharide and its derivatives. Int. J. Biol. Macromol. 2019, 138, 673–680. [Google Scholar] [CrossRef]
- Kumar, R.; Balaji, S.; Sripriya, R.; Nithya, N.; Uma, T.S.; Sehgal, P.K. In vitro evaluation of antioxidants of fruit extract of Momordica charantia L. on fibroblasts and keratinocytes. J. Agric. Food Chem. 2010, 58, 1518–1522. [Google Scholar] [CrossRef]
- Wang, R.S.; Dong, P.H.; Shuai, X.X.; Chen, M.S. Evaluation of different black mulberry fruits (Morus nigra L.) based on phenolic compounds and antioxidant activity. Foods 2022, 11, 1252. [Google Scholar] [CrossRef] [PubMed]
- Kubola, J.; Siriamornpun, S. Phenolic contents and antioxidant activities of bitter gourd (Momordica charantia L.) leaf, stem and fruit fraction extracts in vitro. Food Chem. 2008, 110, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.C.; Liu, Y.J.; He, G.R.; Cao, Y.W.; Bi, M.M.; Song, M.; Yang, P.P.; Xu, L.F.; Ming, J. Comprehensive analysis of secondary metabolites in the extracts from different Lily Bulbs and their antioxidant ability. Antioxidants 2021, 10, 1634. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.P.; Stathopoulos, C.; Parks, S.; Roach, P. An optimised aqueous extract of phenolic compounds from bitter melon with high antioxidant capacity. Antioxidants 2014, 3, 814–829. [Google Scholar] [CrossRef]
- Horax, R.; Hettiarachchy, N.; Islam, S. Total phenolic contents and phenolic acid constituents in 4 varieties of bitter melons (Momordica charantia) and antioxidant activities of their Extracts. J. Food Sci. 2010, 70, C275–C280. [Google Scholar] [CrossRef]
- Horax, R.; Hettiarachchy, N.; Chen, P. Extraction, quantification, and antioxidant activities of phenolics from pericarp and seeds of bitter melons (Momordica charantia) harvested at three maturity stages (immature, mature, and ripe). J. Agric. Food Chem. 2010, 58, 4428–4433. [Google Scholar] [CrossRef]
- Budrat, P.; Shotipruk, A.J.S.; Technology, P. Enhanced recovery of phenolic compounds from bitter melon (Momordica charantia) by subcritical water extraction. Sep. Purif. Technol. 2009, 66, 125–129. [Google Scholar] [CrossRef]
- Jia, S.; Shen, M.; Zhang, F.; Xie, J. Recent advances in Momordica charantia: Functional components and biological activities. Int. J. Mol. Sci. 2017, 18, 2555. [Google Scholar] [CrossRef]
- Gepdiremen, A.; Mshvildadze, V.; Süleyman, H.; Elias, R. Acute anti-inflammatory activity of four saponins isolated from ivy: Alpha-hederin, hederasaponin-C, hederacolchiside-E and hederacolchiside-F in carrageenan-induced rat paw edema. Phytomed. Int. J. Phytother. Phytopharm. 2005, 12, 440–444. [Google Scholar] [CrossRef]
- Fawole, O.A.; Amoo, S.O.; Ndhlala, A.R.; Light, M.E.; Finnie, J.F.; Van Staden, J. Anti-inflammatory, anticholinesterase, antioxidant and phytochemical properties of medicinal plants used for pain-related ailments in South Africa. J. Ethnopharmacol. 2010, 127, 235–241. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef] [PubMed]
- Irmisch, S.; Ruebsam, H.; Jancsik, S.; Man Saint Yuen, M.; Madilao, L.L.; Bohlmann, J. Flavonol biosynthesis genes and their use in engineering the plant antidiabetic metabolite Montbretin A. Plant Physiol. 2019, 180, 1277–1290. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, L.J.; Wang, Y.; Liu, S.; Geng, Z.; Song, A.; Jiang, J.; Chen, S.; Chen, F. Functional identification of a flavone synthase and a flavonol synthase genes affecting flower color formation in Chrysanthemum morifolium. Plant Physiol. Biochem. PPB 2021, 166, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Hua, F.; Li, J.Y.; Zhang, M.; Zhou, P.; Wang, L.; Ling, T.J.; Bao, G.H. Kaempferol-3-O-rutinoside exerts cardioprotective effects through NF-κB/NLRP3/Caspase-1 pathway in ventricular remodeling after acute myocardial infarction. J. Food Biochem. 2022, 46, e14305. [Google Scholar] [CrossRef] [PubMed]
- Borges, P.H.O.; Pedreiro, S.; Baptista, S.J.; Geraldes, C.; Batista, M.T.; Silva, M.M.C.; Figueirinha, A. Inhibition of α-glucosidase by flavonoids of Cymbopogon citratus (DC) Stapf. J. Ethnopharmacol. 2021, 280, 114470. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, S.; Yang, S.; Chen, C.; Yang, Y.; Lin, M.; Liu, C.; Wang, W.; Zhou, X.; Ai, Q.; et al. Mechanism of dihydromyricetin on inflammatory diseases. Front. Pharmacol. 2021, 12, 794563. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wang, F.; Tang, B.; Han, J.; Li, X.; Lian, G.; Li, X.; Hao, S. Anti-inflammatory effects of kaempferol-3-O-rhamnoside on HSV-1 encephalitis in vivo and in vitro. Neurosci. Lett. 2021, 765, 136172. [Google Scholar] [CrossRef]
- Tuli, H.S.; Aggarwal, V.; Kaur, J.; Aggarwal, D.; Parashar, G.; Parashar, N.C.; Tuorkey, M.; Kaur, G.; Savla, R.; Sak, K.; et al. Baicalein: A metabolite with promising antineoplastic activity. Life Sci. 2020, 259, 118183. [Google Scholar] [CrossRef]
- Wang, W.; Yue, R.F.; Jin, Z.; He, L.M.; Shen, R.; Du, D.; Tang, Y.Z. Efficiency comparison of apigenin-7-O-glucoside and trolox in antioxidative stress and anti-inflammatory properties. J. Pharm. Pharmacol. 2020, 72, 1645–1656. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, K.; Sezai Cicek, S.; Zidorn, C.; Granica, S. Phenolic constituents of the aerial parts of Impatiens glandulifera Royle (Balsaminaceae) and their antioxidant activities. Nat. Prod. Res. 2019, 33, 2851–2855. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.; Fang, Y.; De Mukhopdhyay, K.; Wargovich, M.J. Natural agents used in chemoprevention of aerodigestive and GI cancers. Curr. Pharmacol. Rep. 2016, 2, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Gulluce, M.; Orhan, F.; Adiguzel, A.; Bal, T.; Guvenalp, Z.; Dermirezer, L.O. Determination of antimutagenic properties of apigenin-7-O-rutinoside, a flavonoid isolated from Mentha longifolia (L.) Huds. ssp. longifolia with yeast DEL assay. Toxicol. Ind. Health 2013, 29, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Helferich, W.G.; Andrade, J.E.; Hoagland, M.S. Phytoestrogens and breast cancer: A complex story. Inflammopharmacology 2008, 16, 219–226. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Quispe, C.; Imran, M.; Rauf, A.; Nadeem, M.; Gondal, T.A.; Ahmad, B.; Atif, M.; Mubarak, M.S.; Sytar, O.; et al. Genistein: An integrative overview of its mode of action, pharmacological properties, and health benefits. Oxidative Med. Cell. Longev. 2021, 2021, 3268136. [Google Scholar] [CrossRef]
- Sharma, E.; Anand, G.; Kapoor, R. Terpenoids in plant and arbuscular mycorrhiza-reinforced defence against herbivorous insects. Ann. Bot. 2017, 119, 791–801. [Google Scholar] [CrossRef]
- Tholl, D. Biosynthesis and biological functions of terpenoids in plants. Adv. Biochem. Eng. Biotechnol. 2015, 148, 63–106. [Google Scholar]
- El Hazzam, K.; Hafsa, J.; Sobeh, M.; Mhada, M.; Taourirte, M.; El Kacimi, K.; Yasri, A. An insight into saponins from Quinoa (Chenopodium quinoa Willd): A Review. Molecules 2020, 25, 1059. [Google Scholar] [CrossRef]
- Liu, Y.; Mu, S.; Chen, W.; Liu, S.; Cong, Y.; Liu, J.; Jia, N. Saponins of Momordica charantia increase insulin secretion in INS-1 pancreatic β-cells via the PI3K/Akt/FoxO1 signaling pathway. Endocrinol. Diabetes Y Nutr. 2021, 68, 329–337. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Yi, P.F.; Guo, X.; Wu, S.C.; Fu, Y.X.; Zhang, C.; Fu, B.D.; Shen, H.Q.; Wei, X.B. Astragaloside IV inhibits the inflammatory injury of chicken type II pneumocytes induced by avian pathogenic Escherichia coli. Inflammation 2016, 39, 1660–1669. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.; Dong, L.; Thorsen, T.H.; Raadam, M.H.; Khakimov, B.; Carreno-Quintero, N.; Kampranis, S.C.; Bak, S. Metabolic engineering of cucurbitacins in Cucurbita pepo hairy roots. Front. Plant Sci. 2022, 13, 1021907. [Google Scholar] [CrossRef] [PubMed]
- Attard, E.; Martinoli, M.G. Cucurbitacin E, An experimental lead triterpenoid with anticancer, immunomodulatory and novel Effects against degenerative diseases. A Mini-Review. Curr. Top. Med. Chem. 2015, 15, 1708–1713. [Google Scholar] [CrossRef] [PubMed]
- Yuan, R.; Zhao, W.; Wang, Q.Q.; He, J.; Han, S.; Gao, H.; Feng, Y.; Yang, S. Cucurbitacin B inhibits non-small cell lung cancer in vivo and in vitro by triggering TLR4/NLRP3/GSDMD-dependent pyroptosis. Pharmacol. Res. 2021, 170, 105748. [Google Scholar] [CrossRef]
- Cuc, N.T.; Cuong, N.T.; Anh, L.T.; Yen, D.T.H.; Tai, B.H.; Thu Trang, D.; Yen, P.H.; Kiem, P.V.; Nam, N.H.; Minh, C.V.; et al. Dihydrostilbene glycosides from Camellia sasanqua and their α-glucosidase and α-amylase inhibitory activities. Nat. Prod. Res. 2021, 35, 4025–4031. [Google Scholar] [CrossRef]
- Tang, Y.; Li, W.; Cao, J.; Li, W.; Zhao, Y. Bioassay-guided isolation and identification of cytotoxic compounds from Bolbostemma paniculatum. J. Ethnopharmacol. 2015, 169, 18–23. [Google Scholar] [CrossRef]
- Qiu, X.; Zhang, Y.; Zhou, Y.; Li, G.H.; Feng, X.S. Progress in pretreatment and analysis of organic Acids: An update since 2010. Food Chem. 2021, 360, 129977. [Google Scholar] [CrossRef]
- Hasnaoui, N.; Jbir, R.; Mars, M.; Trifi, M.; Kamal-Eldin, A.; Melgarejo, P.; Hernandez, F. Organic acids, sugars, and anthocyanins contents in juices of tunisian pomegranate fruits. Int. J. Food Prop. 2011, 14, 741–757. [Google Scholar] [CrossRef]
- Mattey, M. The production of organic acids. Crit. Rev. Biotechnol. 1992, 12, 87–132. [Google Scholar] [CrossRef]
- Chini, A.; Monte, I.; Zamarreño, A.M.; Hamberg, M.; Lassueur, S.; Reymond, P.; Weiss, S.; Stintzi, A.; Schaller, A.; Porzel, A.; et al. An OPR3-independent pathway uses 4,5-didehydrojasmonate for jasmonate synthesis. Nat. Chem. Biol. 2018, 14, 171–178. [Google Scholar] [CrossRef]
- Schulze, A.; Zimmer, M.; Mielke, S.; Stellmach, H.; Melnyk, C.W.; Hause, B.; Gasperini, D. Wound-induced shoot-to-root relocation of JA-Ile precursors coordinates Arabidopsis growth. Mol. Plant 2019, 12, 1383–1394. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Zhou, Y.; Zhou, M.; Yan, J.; Khurshid, M.; Weng, W.; Cheng, J.; Zhang, K. Jasmonic acid signaling pathway in plants. Int. J. Mol. Sci. 2019, 20, 2479. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Choi, D.; Son, B.; Park, S.; Park, T. Activation of OR10A3 by suberic acid promotes collagen synthesis in UVB-irradiated dermal fibroblasts via the cAMP-Akt pathway. Cells 2022, 11, 3961. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Choi, D.; Park, T. Dietary suberic acid protects against UVB-induced skin photoaging in hairless mice. Nutrients 2019, 11, 2948. [Google Scholar] [CrossRef]
- Lederer, A.K.; Hannibal, L.; Hettich, M.; Behringer, S.; Spiekerkoetter, U.; Steinborn, C.; Gründemann, C.; Zimmermann-Klemd, A.M.; Müller, A.; Simmet, T.; et al. Vitamin B12 status upon short-term intervention with a vegan diet—A randomized controlled trial in healthy participants. Nutrients 2019, 11, 2815. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, H.; Zhang, X.; Chen, Q.; Xia, L. Role of succinic acid in the regulation of sepsis. Int. Immunopharmacol. 2022, 110, 109065. [Google Scholar] [CrossRef]
- Tahjib-Ul-Arif, M.; Zahan, M.I.; Karim, M.M.; Imran, S.; Hunter, C.T.; Islam, M.S.; Mia, M.A.; Hannan, M.A.; Rhaman, M.S.; Hossain, M.A.; et al. Citric acid-mediated abiotic stress tolerance in plants. Int. J. Mol. Sci. 2021, 22, 7235. [Google Scholar] [CrossRef]
- Bhambhani, S.; Kondhare, K.R.; Giri, A.P. Diversity in chemical structures and biological properties of plant alkaloids. Molecules 2021, 26, 3374. [Google Scholar] [CrossRef]
- Day, C.R.; Kempson, S.A. Betaine chemistry, roles, and potential use in liver disease. Biochim. Biophys. Acta 2016, 1860, 1098–1106. [Google Scholar] [CrossRef]
- Saxena, V.K.; De Deyn, P.P. Ergotamine: Its use in the treatment of migraine and its complications. Acta Neurol. 1992, 14, 140–146. [Google Scholar]
- Zhang, W.; Shen, Y.; Li, Z.; Xie, X.; Gong, E.S.; Tian, J.; Si, X.; Wang, Y.; Gao, N.; Shu, C.; et al. Effects of high hydrostatic pressure and thermal processing on anthocyanin content, polyphenol oxidase and β-glucosidase activities, color, and antioxidant activities of blueberry (Vaccinium spp.) puree. Food Chem. 2021, 342, 128564. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Lv, L.; Zeng, X.; Zhang, F.; Chen, Y.; Tian, W.; Li, J.; Li, X.; Li, Y. Integrative analysis of metabolomics and transcriptomics reveals molecular mechanisms of anthocyanin metabolism in the zikui tea plant (Camellia sinensis cv. Zikui). Int. J. Mol. Sci. 2022, 23, 4780. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, R.; Francioso, A.; Mosca, L.; Silva, P. Anthocyanins: A comprehensive review of their chemical properties and health effects on cardiovascular and neurodegenerative diseases. Molecules 2020, 25, 3809. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Xiao, C.; Zhang, T.; Zheng, Y.; Cosgrove, D.J.; Anderson, C.T. Xyloglucan deficiency disrupts microtubule stability and cellulose biosynthesis in arabidopsis, altering cell growth and morphogenesis. Plant Physiol. 2016, 170, 234–249. [Google Scholar] [CrossRef]
- Chen, Q.; Ding, J.; Cai, J.; Sun, Z.; Zhao, J. Simultaneous measurement of total acid content and soluble salt-free solids content in Chinese vinegar using near-infrared spectroscopy. J. Food Sci. 2012, 77, C222–C227. [Google Scholar] [CrossRef]
- Tomar, O.; Akarca, G.; GÖk, V.; Çağlar, M.Y. The effects of packaging materials on the fatty acid composition, organic acid content, and texture profiles of Tulum cheese. J. Food Sci. 2020, 85, 3134–3140. [Google Scholar] [CrossRef]
- Matić, P.; Sabljić, M.; Jakobek, L. Validation of spectrophotometric methods for the determination of total polyphenol and total flavonoid content. J. AOAC Int. 2017, 100, 1795–1803. [Google Scholar] [CrossRef]
- He, X.; Wang, Y.; Hu, H.; Zhang, Z. In vitro and in vivo antimammary tumor activities and mechanisms of the apple total triterpenoids. J. Agric. Food Chem. 2012, 60, 9430–9436. [Google Scholar] [CrossRef]
- Alzagameem, A.; Khaldi-Hansen, B.E.; Büchner, D.; Larkins, M.; Kamm, B.; Witzleben, S.; Schulze, M. Lignocellulosic biomass as source for lignin-based environmentally benign antioxidants. Molecules 2018, 23, 2664. [Google Scholar] [CrossRef]
- Yang, L.; Wang, D.; Zhou, D.; Zhang, Y. Effect of different isolation methods on structure and properties of lignin from valonea of Quercus variabilis. Int. J. Biol. Macromol. 2016, 85, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Rumpf, J.; Burger, R.; Schulze, M. Statistical evaluation of DPPH, ABTS, FRAP, and Folin-Ciocalteu assays to assess the antioxidant capacity of lignins. Int. J. Biol. Macromol. 2023, 233, 123470. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Luo, J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: Application in the study of rice metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res. 2023, 51, D587–D592. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Vc Content (μg/g FW) | Soluble Protein Content (mg/g FW) | Amino Acid Content (mg/g FW) | Cellulose Content (mg/g DW) | Total Acid Content (%) |
|---|---|---|---|---|---|
| PG | 117.28 ± 3.58 a | 4.70 ± 0.34 d | 1.05 ± 0.04 e | 298.79 ± 15.28 bc | 0.41 ± 0.01 cd |
| JLZ | 121.55 ± 11.88 a | 12.83 ± 0.51 a | 1.64 ± 0.02 c | 284.81 ± 7.45 c | 0.28 ± 0.00 e |
| BFM | 56.65 ± 1.59 b | 5.13 ± 0.06 d | 2.03 ± 0.03 a | 399.46 ± 5.52 a | 0.40 ± 0.01 d |
| RB | 125.83 ± 6.08 a | 6.07 ± 0.34 c | 0.52 ± 0.03 f | 274.94 ± 2.83 c | 0.47 ± 0.00 b |
| CB | 126.57 ± 9.16 a | 11.52 ± 0.17 b | 1.16 ± 0.02 d | 372.33 ± 2.96 a | 0.40 ± 0.00 cd |
| LJ | 65.28 ± 5.09 b | 6.20 ± 0.02 c | 1.86 ± 0.05 b | 318.35 ± 21.41 b | 0.52 ± 0.02 a |
| LBS | 63.07 ± 0.78 b | 13.30 ± 0.57 a | 2.03 ± 0.05 a | 287.31 ± 19.39 c | 0.42 ± 0.00 c |
| Cultivar | Oxalic Acid (mg/kg) | Succinic Acid (mg/kg) | Malic Acid (mg/kg) | Citric Acids (mg/kg) |
|---|---|---|---|---|
| PG | 206.05 ± 13.34 de | 44.19 ± 2.12 b | 853.54 ± 50.04 d | 41.02 ± 2.07 d |
| JLZ | 341.86 ± 31.26 ab | 26.68 ± 0.73 c | 2214.07 ± 123.42 a | 186.20 ± 18.59 a |
| BFM | 221.07 ± 14.99 de | 26.50 ± 1.05 c | 1092.73 ± 81.41 c | 107.19 ± 11.44 b |
| RB | 255.47 ± 25.31 cd | 30.30 ± 3.28 c | 800.29 ± 81.34 d | 43.84 ± 2.27 d |
| CB | 302.29 ± 25.27 bc | 63.10 ± 2.18 a | 2121.76 ± 56.72 a | 108.38 ± 7.18 b |
| LJ | 365.75 ± 67.11 a | 20.47 ± 2.11 d | 1343.16 ± 85.76 b | 68.90 ± 11.12 c |
| LBS | 164.22 ± 6.11 e | 16.79 ± 2.36 d | 594.00 ± 76.59 e | 37.78 ± 3.65 d |
| Cultivar | TPC (μg/g) | TFC (mg/100 g) | TTC (mg/g) | DPPH (%) | ABTS (μmol/g FW) | FRAP (μmol/g FW) |
|---|---|---|---|---|---|---|
| PG | 17.02 ± 0.64 d | 130.33 ± 9.58 e | 1.23 ± 0.06 e | 0.58 ± 0.02 e | 10.11 ± 0.19 g | 9.64 ± 0.62 d |
| JLZ | 17.60 ± 1.14 d | 135.85 ± 7.26 de | 1.24 ± 0.11 e | 0.62 ± 0.02 e | 10.50 ± 0.30 f | 9.83 ± 0.66 cd |
| BFM | 20.11 ± 1.02 c | 142.58 ± 16.26 cde | 1.33 ± 0.02 de | 0.68 ± 0.01 d | 11.11 ± 0.18 e | 10.14 ± 0.39 cd |
| RB | 21.04 ± 0.75 c | 150.94 ± 8.33 bcd | 1.45 ± 0.08 cd | 0.75 ± 0.03 c | 11.80 ± 0.25 d | 10.84 ± 0.48 c |
| CB | 24.04 ± 0.99 b | 160.24 ± 7.45 bc | 1.72 ± 0.06 a | 0.84 ± 0.02 a | 12.60 ± 0.10 c | 12.84 ± 0.63 b |
| LJ | 29.05 ± 1.23 a | 180.54 ± 10.46 a | 1.59 ± 0.08 ab | 0.81 ± 0.05 ab | 13.48 ± 0.03 a | 13.93 ± 0.53 a |
| LBS | 25.26 ± 1.20 b | 170.59 ± 12.73 ab | 1.49 ± 0.06 bc | 0.77 ± 0.03 bc | 13.00 ± 0.12 b | 13.33 ± 0.63 ab |
| TPC | TFC | TTC | DPPH | ABTS | FRAP | TPC | TFC | TTC | DPPH | ABTS | FRAP | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TPC | PG | 1 | −0.997 ** | −0.997 ** | −0.507 ** | −0.983 ** | 0.999 ** | JLZ | 1 | −0.989 ** | −0.551 ** | −0.925 ** | −0.991 ** | −0.976 ** |
| TFC | 1 | 1 | 0.573 ** | 0.995 ** | −0.995 ** | 1 | 0.671 ** | 0.972 ** | 1 ** | 0.998 ** | ||||
| TTC | 1 | 0.572 ** | 0.994 ** | −0.996 ** | 1 | 0.826 ** | 0.657 ** | 0.720 ** | ||||||
| DPPH | 1 | 0.655 ** | −0.492 ** | 1 | 0.968 ** | 0.986 ** | ||||||||
| ABTS | 1 | −0.980 | 1 | 0.996 | ||||||||||
| FRAP | 1 | 1 | ||||||||||||
| TPC | BFM | 1 | −0.976 ** | 0.133 ** | 0.908 ** | −0.459 ** | 0.581 ** | RB | 1 | 0.181 ** | 0.597 ** | −0.508 ** | 0.418 ** | 0.410 ** |
| TFC | 1 | −0.345 ** | −0.796 ** | 0.640 ** | −0.744 ** | 1 | −0.681 ** | 0.755 ** | 0.969 ** | −0.823 ** | ||||
| TTC | 1 | −0.294 ** | −0.942 ** | 0.884 ** | 1 | −0.994 ** | −0.479 ** | 0.976 ** | ||||||
| DPPH | 1 | −0.045 ** | 0.187 ** | 1 | 0.570 ** | −0.994 ** | ||||||||
| ABTS | 1 | −0.990 * | 1 | −0.657 * | ||||||||||
| FRAP | 1 | 1 | ||||||||||||
| TPC | CB | 1 | −0.467 ** | −0.558 ** | −0.960 ** | 1 ** | 0.959 ** | LJ | 1 | −0.993 ** | 0.340 ** | −0.516 ** | 0.878 ** | −0.960 ** |
| TFC | 1 | −0.473 ** | 0.695 ** | −0.469 ** | −0.698 ** | 1 | −0.446 ** | 0.414 ** | −0.817 ** | 0.986 ** | ||||
| TTC | 1 | 0.304 ** | −0.556 ** | −0.301 ** | 1 | 0.630 ** | −0.152 ** | −0.591 ** | ||||||
| DPPH | 1 | −0.961 ** | −1 ** | 1 | −0.863 ** | 0.254 ** | ||||||||
| ABTS | 1 | 0.960 | 1 | −0.708 | ||||||||||
| FRAP | 1 | 1 | ||||||||||||
| TPC | LBS | 1 | −0.200 ** | 0.975 ** | −0.382 ** | 0.407 ** | 0.302 ** | |||||||
| TFC | 1 | −0.413 ** | −0.829 ** | −0.976 ** | −0.994 ** | |||||||||
| TTC | 1 | −0.167 ** | 0.600 ** | 0.506 ** | ||||||||||
| DPPH | 1 | 0.689 ** | 0.765 ** | |||||||||||
| ABTS | 1 | 0.994 | ||||||||||||
| FRAP | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Lu, P.; Jin, H.; Cui, J.; Miao, C.; He, L.; Yu, J.; Ding, X.; Zhang, H. Integrated Secondary Metabolomic and Antioxidant Ability Analysis Reveals the Accumulation Patterns of Metabolites in Momordica charantia L. of Different Cultivars. Int. J. Mol. Sci. 2023, 24, 14495. https://doi.org/10.3390/ijms241914495
Zhang Y, Lu P, Jin H, Cui J, Miao C, He L, Yu J, Ding X, Zhang H. Integrated Secondary Metabolomic and Antioxidant Ability Analysis Reveals the Accumulation Patterns of Metabolites in Momordica charantia L. of Different Cultivars. International Journal of Molecular Sciences. 2023; 24(19):14495. https://doi.org/10.3390/ijms241914495
Chicago/Turabian StyleZhang, Yongxue, Panling Lu, Haijun Jin, Jiawei Cui, Chen Miao, Lizhong He, Jizhu Yu, Xiaotao Ding, and Hongmei Zhang. 2023. "Integrated Secondary Metabolomic and Antioxidant Ability Analysis Reveals the Accumulation Patterns of Metabolites in Momordica charantia L. of Different Cultivars" International Journal of Molecular Sciences 24, no. 19: 14495. https://doi.org/10.3390/ijms241914495