

Chronic Administration of Ion Channel Blockers Impact Microglia Morphology and Function in a Murine Model of Alzheimer’s Disease

 , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Verapamil Induces Increased Short-Term Memory and Exploration Behavior in APP Mice
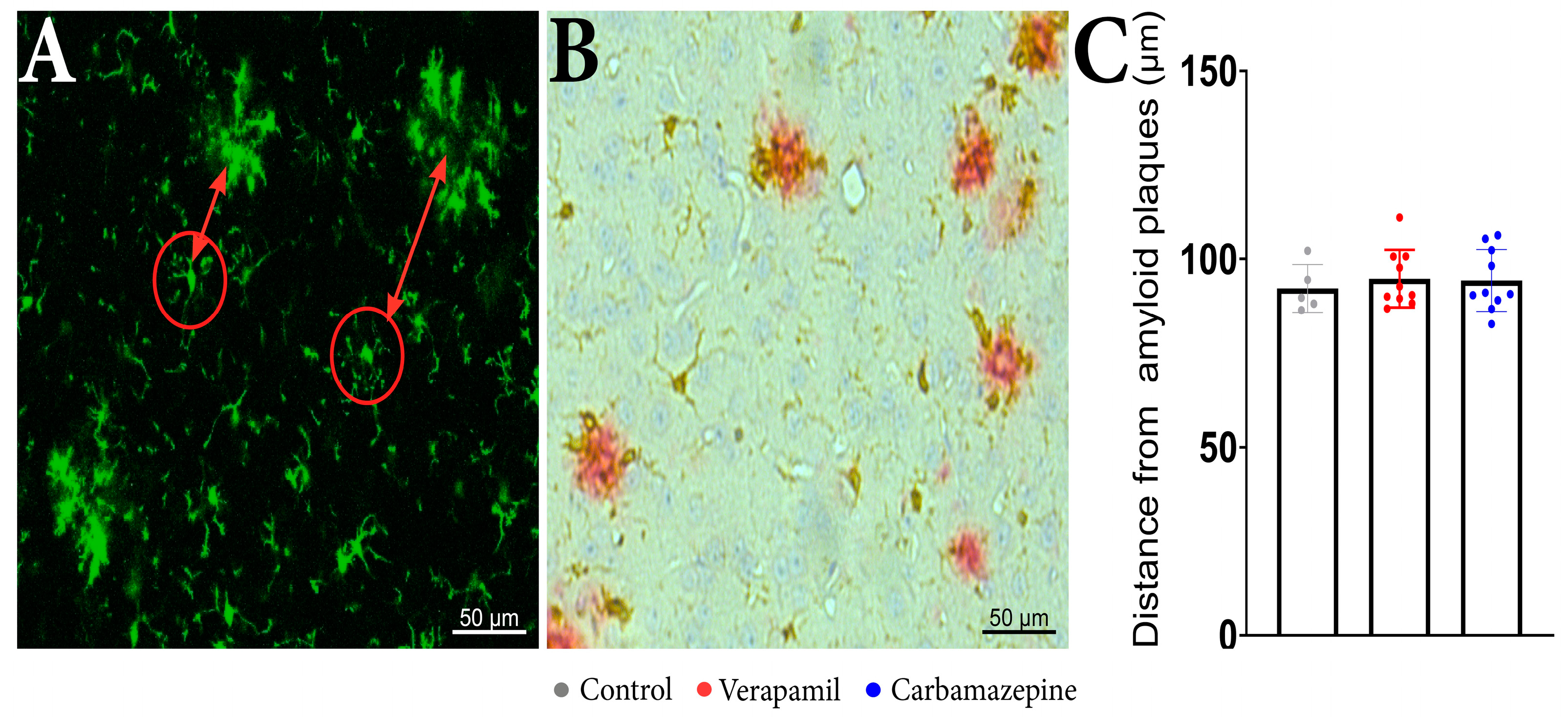
2.2. Microglia Motility and Morphology Are Sensitive to Systemic Ion Channel Blockers
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Behavioral Studies
4.3. Pharmacological Treatment
4.4. Images Acquisition and Processing
4.5. Distance Measurements
4.6. Statistical Analysis
5. Conclusions
Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garin, N.; Koyanagi, A.; Chatterji, S.; Tyrovolas, S.; Olaya, B.; Leonardi, M.; Lara, E.; Koskinen, S.; Tobiasz-Adamczyk, B.; Ayuso-Mateos, J.L.; et al. Global Multimorbidity Patterns: A Cross-Sectional, Population-Based, Multi-Country Study. J. Gerontol. A Biol. Sci. Med. Sci. 2016, 71, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Diesveld, M.M.E.; de Klerk, S.; Cornu, P.; Strobach, D.; Taxis, K.; Borgsteede, S.D. Management of Drug-Disease Interactions: A Best Practice from the Netherlands. Int. J. Clin. Pharm. 2021, 43, 1437–1450. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Wu, Y.J.; Tee, B.L.; Lo, R.Y. Medical Comorbidity in Alzheimer’s Disease: A Nested Case-Control Study. J. Alzheimers Dis. 2018, 63, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Demuro, A.; Mina, E.; Kayed, R.; Milton, S.C.; Parker, I.; Glabe, C.G. Calcium Dysregulation and Membrane Disruption as a Ubiquitous Neurotoxic Mechanism of Soluble Amyloid Oligomers. J. Biol. Chem. 2005, 280, 17294–17300. [Google Scholar] [CrossRef]
- Diaz, J.C.; Simakova, O.; Jacobson, K.A.; Arispe, N.; Pollard, H.B. Small Molecule Blockers of the Alzheimer Abeta Calcium Channel Potently Protect Neurons from Abeta Cytotoxicity. Proc. Natl. Acad. Sci. USA 2009, 106, 3348–3353. [Google Scholar] [CrossRef]
- Cojocaru, A.; Zavaleanu, A.D.; Calina, D.C.; Osiac, E.; Boboc, I.K.S.; Mitran, S.I. Different Age Related Neurological and Cardiac Effects of Verapamil on a Transgenic Mouse Model of Alzheimer’s Disease. Curr. Health Sci. J. 2021, 47, 263–269. [Google Scholar]
- Liu, Y.; Lo, Y.C.; Qian, L.; Crews, F.T.; Wilson, B.; Chen, H.L.; Wu, H.M.; Chen, S.H.; Wei, K.; Lu, R.B.; et al. Verapamil Protects Dopaminergic Neuron Damage through a Novel Anti-Inflammatory Mechanism by Inhibition of Microglial Activation. Neuropharmacology 2011, 60, 373–380. [Google Scholar] [CrossRef]
- Mosalam, E.M.; Elberri, A.I.; Sallam, A.S.; Salem, H.R.; Metwally, E.M.; Abdallah, M.S.; Shaldam, M.A.; Mansour, H.E.A. Chronotherapeutic Neuroprotective Effect of Verapamil against Lipopolysaccharide-Induced Neuroinflammation in Mice through Modulation of Calcium-Dependent Genes. Mol. Med. 2022, 28, 139. [Google Scholar] [CrossRef]
- Guo, S.; Wang, H.; Yin, Y. Microglia Polarization From M1 to M2 in Neurodegenerative Diseases. Front. Aging Neurosci. 2022, 14, 815347. [Google Scholar] [CrossRef]
- Jiang, X.; Wu, Q.; Zhang, C.; Wang, M. Homoharringtonine Inhibits Alzheimer’s Disease Progression by Reducing Neuroinflammation via STAT3 Signaling in APP/PS1 Mice. Neurodegener. Dis. 2022, 21, 93–102. [Google Scholar] [CrossRef]
- Mccarthy, L.H.; Bigal, M.E.; Katz, M.; Derby, C.; Lipton, R.B. Chronic Pain and Obesity in Elderly People: Results from the Einstein Aging Study. J. Am. Geriatr. Soc. 2009, 57, 115–119. [Google Scholar] [CrossRef]
- Nakai, Y.; Makizako, H.; Kiyama, R.; Tomioka, K.; Taniguchi, Y.; Kubozono, T.; Takenaka, T.; Ohishi, M. Association between Chronic Pain and Physical Frailty in Community-Dwelling Older Adults. Int. J. Environ. Res. Public Health 2019, 16, 1330. [Google Scholar] [CrossRef]
- Zhou, B.; Bentham, J.; Di Cesare, M.; Bixby, H.; Danaei, G.; Cowan, M.J.; Paciorek, C.J.; Singh, G.; Hajifathalian, K.; Bennett, J.E.; et al. Worldwide Trends in Blood Pressure from 1975 to 2015: A Pooled Analysis of 1479 Population-Based Measurement Studies with 19·1 Million Participants. Lancet 2017, 389, 37–55. [Google Scholar] [CrossRef] [PubMed]
- Ezzati, M.; Zhou, B.; Bentham, J.; Di Cesare, M.; Bixby, H.; Danaei, G.; Hajifathalian, K.; Taddei, C.; Carrillo-Larco, R.M.; Djalalinia, S.; et al. Contributions of Mean and Shape of Blood Pressure Distribution to Worldwide Trends and Variations in Raised Blood Pressure: A Pooled Analysis of 1018 Population-Based Measurement Studies with 88.6 Million Participants. Int. J. Epidemiol. 2018, 47, 872–883i. [Google Scholar] [CrossRef]
- Cojocaru, A.; Burada, E.; Bălșeanu, A.T.; Deftu, A.F.; Cătălin, B.; Popa-Wagner, A.; Osiac, E. Roles of Microglial Ion Channel in Neurodegenerative Diseases. J. Clin. Med. 2021, 10, 1239. [Google Scholar] [CrossRef] [PubMed]
- Leng, F.; Edison, P. Neuroinflammation and Microglial Activation in Alzheimer Disease: Where Do We Go from Here? Nat. Rev. Neurol. 2020, 17, 157–172. [Google Scholar] [CrossRef]
- Boboc, I.K.S.; Rotaru-Zavaleanu, A.D.; Calina, D.; Albu, C.V.; Catalin, B.; Turcu-Stiolica, A. A Preclinical Systematic Review and Meta-Analysis of Behavior Testing in Mice Models of Ischemic Stroke. Life 2023, 13, 567. [Google Scholar] [CrossRef]
- Nimmerjahn, A.; Kirchhoff, F.; Helmchen, F. Neuroscience: Resting Microglial Cells Are Highly Dynamic Surveillants of Brain Parenchyma in Vivo. Science (80-) 2005, 308, 1314–1318. [Google Scholar] [CrossRef]
- Schürmann, B.; Bermingham, D.P.; Kopeikina, K.J.; Myczek, K.; Yoon, S.; Horan, K.E.; Kelly, C.J.; Martin-de-Saavedra, M.D.; Forrest, M.P.; Fawcett-Patel, J.M.; et al. A Novel Role for the Late-Onset Alzheimer’s Disease (LOAD)-Associated Protein Bin1 in Regulating Postsynaptic Trafficking and Glutamatergic Signaling. Mol. Psychiatry 2020, 25, 2000–2016. [Google Scholar] [CrossRef]
- Stopper, L.; Bălşeanu, T.A.; Cătălin, B.; Rogoveanu, O.C.; Mogoantă, L.; Scheller, A. Microglia Morphology in the Physiological and Diseased Brain—From Fixed Tissue to in Vivo Conditions. Rom. J. Morphol. Embryol. 2018, 59, 7–12. [Google Scholar]
- Morrison, H.; Young, K.; Qureshi, M.; Rowe, R.K.; Lifshitz, J. Quantitative Microglia Analyses Reveal Diverse Morphologic Responses in the Rat Cortex after Diffuse Brain Injury. Sci. Rep. 2017, 7, 13211. [Google Scholar] [CrossRef]
- Godeanu, S.; Clarke, D.; Stopper, L.; Deftu, A.F.; Popa-Wagner, A.; Bălșeanu, A.T.; Scheller, A.; Catalin, B. Microglial Morphology in the Somatosensory Cortex across Lifespan. A Quantitative Study. Dev. Dyn. 2023, 252, 1113–1129. [Google Scholar] [CrossRef]
- Clark, M.B.; Wrzesinski, T.; Garcia, A.B.; Hall, N.A.L.; Kleinman, J.E.; Hyde, T.; Weinberger, D.R.; Harrison, P.J.; Haerty, W.; Tunbridge, E.M. Long-Read Sequencing Reveals the Complex Splicing Profile of the Psychiatric Risk Gene CACNA1C in Human Brain. Mol. Psychiatry 2020, 25, 37–47. [Google Scholar] [CrossRef]
- Kotturi, M.F.; Jefferies, W.A. Molecular Characterization of L-Type Calcium Channel Splice Variants Expressed in Human T Lymphocytes. Mol. Immunol. 2005, 42, 1461–1474. [Google Scholar] [CrossRef] [PubMed]
- Hopp, S.C. Targeting Microglia L-Type Voltage-Dependent Calcium Channels for the Treatment of Central Nervous System Disorders. J. Neurosci. Res. 2021, 99, 141. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, L.; Zhou, C.-N.; Jiang, L.; Tang, J.; Liang, X.; Fan, J.-H.; Dou, X.-Y.; Tang, Y. Oligodendrocyte Generation and Maturation in the Hippocampus May Be a Target of Fluoxetine for Delaying Cognitive Dysfunction during Early Alzheimer’s Disease. arXiv 2020. [Google Scholar] [CrossRef]
- Hoyt, C.T.; Domingo-Fernández, D.; Balzer, N.; Güldenpfennig, A.; Hofmann-Apitius, M. A Systematic Approach for Identifying Shared Mechanisms in Epilepsy and Its Comorbidities. Database 2018, 2018, bay050. [Google Scholar] [CrossRef]
- Wang, C.C.; Kong, J.Y.; Li, X.Y.; Yang, J.Y.; Xue, C.H.; Yanagita, T.; Wang, Y.M. Antarctic Krill Oil Exhibited Synergistic Effects with Nobiletin and Theanine in Ameliorating Memory and Cognitive Deficiency in SAMP8 Mice: Applying the Perspective of the Sea–Land Combination to Retard Brain Aging. Front. Aging Neurosci. 2022, 14, 964077. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Huang, W.; Lu, J.; Chen, H.; Yu, Z. TRPV1-Mediated Microglial Autophagy Attenuates Alzheimer’s Disease-Associated Pathology and Cognitive Decline. Front. Pharmacol. 2022, 12, 763866. [Google Scholar] [CrossRef]
- Radde, R.; Bolmont, T.; Kaeser, S.A.; Coomaraswamy, J.; Lindau, D.; Stoltze, L.; Calhoun, M.E.; Jäggi, F.; Wolburg, H.; Gengler, S.; et al. Aβ42-Driven Cerebral Amyloidosis in Transgenic Mice Reveals Early and Robust Pathology. EMBO Rep. 2006, 7, 940. [Google Scholar] [CrossRef]
- Verapamil Appears to Reduce Anxiety and Panic Attacks. InPharma 1988, 637, 12. [CrossRef]
- Uhde, T.W.; Ballenger, J.C.; Post, R.M. Carbamazepine: Treatment of Affective Illness and Anxiety Syndromes. Psychiatry State Art 1985, 17, 479–484. [Google Scholar] [CrossRef]
- Kuryshev, Y.A.; Brown, A.M.; Duzic, E.; Kirsch, G.E. Evaluating State Dependence and Subtype Selectivity of Calcium Channel Modulators in Automated Electrophysiology Assays. Assay Drug Dev. Technol. 2014, 12, 110. [Google Scholar] [CrossRef] [PubMed]
- Rice, R.A.; Berchtold, N.C.; Cotman, C.W.; Green, K.N. Age-Related Downregulation of the CaV3.1 T-Type Calcium Channel as a Mediator of Amyloid Beta Production. Neurobiol. Aging 2014, 35, 1002–1011. [Google Scholar] [CrossRef]
- Ismail, F.S.; Corvace, F.; Faustmann, P.M.; Faustmann, T.J. Pharmacological Investigations in Glia Culture Model of Inflammation. Front. Cell. Neurosci. 2021, 15, 805755. [Google Scholar] [CrossRef]
- Franklin, R.J.M.; Ffrench-Constant, C.; Edgar, J.M.; Smith, K.J. Neuroprotection and Repair in Multiple Sclerosis. Nat. Rev. Neurol. 2012, 8, 624–634. [Google Scholar] [CrossRef]
- Cruz, M.P. Aripiprazole Lauroxil (Aristada): An Extended-Release, Long-Acting Injection For the Treatment of Schizophrenia. Pharm. Ther. 2016, 41, 556. [Google Scholar]
- Zhou, S.-F. Drugs Behave as Substrates, Inhibitors and Inducers of Human Cytochrome P450 3A4. Curr. Drug Metab. 2008, 9, 310–322. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.L.; Hyeon, M.R.; Myung, H.B.; Yong, S.K.; Ju, H.L.; Yongwhi, P.; Heo, J.H.; Young, S.L.; Dong, H.Y.; Hun, S.P.; et al. Prognosis and Natural History of Drug-Related Bradycardia. Korean Circ. J. 2009, 39, 367–371. [Google Scholar] [CrossRef]
- FDA 2002 Highlights of Prescribing Information Abilify® (Aripiprazole). Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2014/021436s038,021713s030,021729s022,021866s023lbl.pdf (accessed on 10 February 2023).
- Ahmed, H.; Ishrat, T. Repurposing Verapamil for Prevention of Cognitive Decline in Sporadic Alzheimer’s Disease. Neural. Regen. Res. 2022, 17, 1018. [Google Scholar] [CrossRef]
- Jung, S.; Aliberti, J.; Graemmel, P.; Sunshine, M.J.; Kreutzberg, G.W.; Sher, A.; Littman, D.R. Analysis of Fractalkine Receptor CX(3)CR1 Function by Targeted Deletion and Green Fluorescent Protein Reporter Gene Insertion. Mol. Cell. Biol. 2000, 20, 4106–4114. [Google Scholar] [CrossRef] [PubMed]
- Lueptow, L.M. Novel Object Recognition Test for the Investigation of Learning and Memory in Mice. J. Vis. Exp. 2017, 2017, 55718. [Google Scholar] [CrossRef]
- Cătălin, B.; Mitran, S.; Albu, C.; Iancău, M. Comparative Aspects of Microglia Reaction in White and Gray Matter. Curr. Health Sci. J. 2013, 39, 151. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, A.; Kirchhoff, F.; Kerr, J.N.D.; Helmchen, F. Sulforhodamine 101 as a Specific Marker of Astroglia in the Neocortex in Vivo. Nat. Methods 2004, 1, 31–37. [Google Scholar] [CrossRef]
- Lowe, D.G. Distinctive Image Features from Scale-Invariant Keypoints. Int. J. Comput. Vis. 2004, 60, 91–110. [Google Scholar] [CrossRef]
- Registration. Available online: https://imagej.net/imaging/registration (accessed on 10 August 2023).
- Cătălin, B.; Alexandru, D.; Albu, C.; Iancău, M. Original Paper Microglia Branching Using a Sholl Analysis Method. Curr. Health Sci. J. 2013, 39, 1–5. [Google Scholar]
- Sholl Analysis. Available online: https://imagej.net/plugins/sholl-analysis (accessed on 10 August 2023).
- Jung, C.K.E.; Keppler, K.; Steinbach, S.; Blazquez-Llorca, L.; Herms, J. Fibrillar Amyloid Plaque Formation Precedes Microglial Activation. PLoS ONE 2015, 10, e0119768. [Google Scholar] [CrossRef]
- Huang, Y.; Happonen, K.E.; Burrola, P.G.; O’Connor, C.; Hah, N.; Huang, L.; Nimmerjahn, A.; Lemke, G. Microglia Use TAM Receptors to Detect and Engulf Amyloid β Plaques. Nat. Immunol. 2021, 22, 586–594. [Google Scholar] [CrossRef]
- Hierro-Bujalance, C.; Bacskai, B.J.; Garcia-Alloza, M. In Vivo Imaging of Microglia with Multiphoton Microscopy. Front. Aging Neurosci. 2018, 10, 379872. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Distance (µm) | Verapamil | Carbamazepine | Control | WT | Statistical Difference | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |||
| 1 | 10.3725 | −5.7831 | 0.139557 | −5.7272 | 0.322582 | −4.97413 | 0.671435 | −4.663 | 0.143934 | ## &&&& * xxx |
| 2 | 12.447 | −6.05621 | 0.133852 | −6.07918 | 0.216208 | −5.26317 | 0.662206 | −4.954 | 0.14545 | ## &&& ** xxx |
| 3 | 14.5215 | −6.2927 | 0.122836 | −6.2817 | 0.318961 | −5.5213 | 0.653246 | −5.214 | 0.14804 | ## &&& * xxx |
| 4 | 16.596 | −6.51083 | 0.11892 | −6.51614 | 0.320295 | −5.75362 | 0.645137 | −5.448 | 0.15125 | |
| 5 | 18.6705 | −6.70983 | 0.116308 | −6.72914 | 0.322799 | −5.96736 | 0.636441 | −5.665 | 0.15497 | |
| 6 | 20.745 | −6.89286 | 0.114638 | −6.9243 | 0.326087 | −6.16294 | 0.628662 | −5.863 | 0.15898 | |
| 7 | 22.8195 | −7.06227 | 0.113641 | −7.1044 | 0.329918 | −6.34392 | 0.6213 | −6.046 | 0.16307 | |
| 8 | 24.894 | −7.21997 | 0.113138 | −7.27158 | 0.334097 | −6.51256 | 0.614668 | −6.217 | 0.16727 | |
| 9 | 26.9685 | −7.36821 | 0.112439 | −7.43002 | 0.340929 | −6.67269 | 0.609749 | −6.380 | 0.17706 | # &&& * xx |
| 10 | 29.043 | −7.50673 | 0.112679 | −7.57732 | 0.346555 | −6.82148 | 0.6052 | −6.529 | 0.18101 | |
| 11 | 31.1175 | −7.63621 | 0.114154 | −7.7148 | 0.351021 | −6.96232 | 0.599641 | −6.672 | 0.17927 | # && * xx |
| 12 | 33.192 | −7.75639 | 0.1175 | −7.8475 | 0.35843 | −7.09622 | 0.595547 | −6.807 | 0.18833 | |
| 13 | 35.2665 | −7.8771 | 0.113572 | −7.98082 | 0.373796 | −7.22297 | 0.591105 | −6.934 | 0.19231 | |
| 14 | 37.341 | −7.99016 | 0.114175 | −8.08488 | 0.365638 | −7.34511 | 0.586263 | −7.059 | 0.19820 | |
| 15 | 39.4155 | −8.1003 | 0.114734 | −8.16646 | 0.339129 | −7.47236 | 0.577831 | −7.196 | 0.21528 | |
| 16 | 41.49 | −8.18716 | 0.122873 | −8.27062 | 0.263343 | −7.58284 | 0.573399 | −7.306 | 0.21771 | |
| 17 | 43.5645 | −8.26976 | 0.142779 | −8.41186 | 0.206976 | −7.66377 | 0.596221 | −7.367 | 0.28199 | |
| 18 | 45.639 | −8.3628 | 0.144326 | −8.50794 | 0.187582 | −7.76163 | 0.594916 | −7.465 | 0.28714 | |
| 19 | 47.7135 | −8.45017 | 0.14861 | −8.49078 | 0.371688 | −7.84599 | 0.588008 | −7.541 | 0.24247 | |
| 20 | 49.788 | −8.53619 | 0.150288 | −8.58276 | 0.360576 | −7.91597 | 0.606144 | −7.586 | 0.24205 | |
| 21 | 51.8625 | −8.61907 | 0.151902 | −8.81828 | 0.186521 | −7.98178 | 0.668145 | −7.619 | 0.35071 | # &&& ** xxx |
| 22 | 53.937 | −8.69929 | 0.15317 | −8.86 | 0.170743 | −8.05414 | 0.637683 | −7.701 | 0.35160 | ## &&& * xxx |
| 23 | 56.0115 | −8.77759 | 0.154672 | −8.9484 | 0.15765 | −8.13477 | 0.642158 | −7.777 | 0.35751 | |
| 24 | 58.086 | −8.85951 | 0.147872 | −9.0011 | 0.232461 | −8.2165 | 0.642421 | −7.851 | 0.36161 | |
| 25 | 60.1605 | −8.93364 | 0.149098 | −9.06893 | 0.185161 | −8.3032 | 0.64599 | −7.925 | 0.36179 | |
| 26 | 62.235 | −8.99816 | 0.13772 | −9.09815 | 0.265973 | −8.36863 | 0.660695 | −7.993 | 0.38311 | # &&& * xx |
| 27 | 64.3095 | −9.0684 | 0.144437 | −9.10358 | 0.401635 | −8.4352 | 0.647236 | −8.073 | 0.39104 | # &&& xx |
| 28 | 66.384 | −9.1613 | 0.13027 | −9.1714 | 0.404112 | −8.53611 | 0.689189 | −8.142 | 0.39530 | |
| 29 | 68.4585 | −9.20471 | 0.151935 | −9.23785 | 0.406829 | −8.64203 | 0.770144 | −8.203 | 0.39999 | &&& xx |
| 30 | 70.533 | −9.24374 | 0.166534 | −9.30785 | 0.411824 | −8.68916 | 0.685098 | −8.288 | 0.42385 | && xx |
| 31 | 72.6075 | −9.29834 | 0.165424 | −9.361 | 0.415674 | −8.75859 | 0.684737 | −8.356 | 0.43166 | |
| 32 | 74.682 | −9.34884 | 0.165499 | −9.62873 | 0.148646 | −8.8059 | 0.685321 | −8.417 | 0.43390 | && * xx |
| 33 | 76.7565 | −9.396 | 0.143285 | −9.7088 | 0.159811 | −9.00829 | 0.580387 | −8.690 | 0.25807 | x |
| 34 | 78.831 | −9.45221 | 0.144371 | −9.7904 | 0.167395 | −9.06555 | 0.589108 | −8.726 | 0.29280 | & xx |
| 35 | 80.9055 | −9.51339 | 0.153587 | −9.89917 | 0.240864 | −9.13456 | 0.813707 | −8.661 | 0.50328 | |
| 36 | 82.98 | −9.56846 | 0.172276 | −9.93613 | 0.204159 | −9.19769 | 0.824955 | −8.729 | 0.54609 | |
| 37 | 85.0545 | −9.5962 | 0.181902 | −9.91117 | 0.185086 | −9.24154 | 0.829573 | −8.783 | 0.55095 | |
| 38 | 87.129 | −9.64963 | 0.18594 | −9.9695 | 0.181791 | −9.17946 | 0.770714 | −8.803 | 0.57303 | & * xx |
| 39 | 89.2035 | −9.7008 | 0.167524 | −10.0297 | 0.176193 | −9.24964 | 0.775958 | −8.891 | 0.62076 | |
| 40 | 91.278 | −9.75289 | 0.171649 | −10.0795 | 0.176856 | −9.06428 | 0.524778 | −8.953 | 0.63885 | # & ** xx |
| 41 | 93.3525 | −9.78566 | 0.159271 | −10.1013 | 0.277616 | −9.10405 | 0.458513 | −8.994 | 0.54934 | |
| 42 | 95.427 | −9.87427 | 0.18834 | −10.1585 | 0.281657 | −9.09462 | 0.450958 | −9.052 | 0.54198 | |
| 43 | 97.5015 | −9.88828 | 0.192042 | −10.1612 | 0.222306 | −9.2539 | 0.378871 | −9.265 | 0.47044 | * |
| 44 | 99.576 | −9.95285 | 0.307011 | −10.1703 | 0.229126 | −9.34927 | 0.285649 | −9.478 | 0.23121 | No Diff |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boboc, I.K.S.; Cojocaru, A.; Nedelea, G.; Catalin, B.; Bogdan, M.; Calina, D. Chronic Administration of Ion Channel Blockers Impact Microglia Morphology and Function in a Murine Model of Alzheimer’s Disease. Int. J. Mol. Sci. 2023, 24, 14474. https://doi.org/10.3390/ijms241914474
Boboc IKS, Cojocaru A, Nedelea G, Catalin B, Bogdan M, Calina D. Chronic Administration of Ion Channel Blockers Impact Microglia Morphology and Function in a Murine Model of Alzheimer’s Disease. International Journal of Molecular Sciences. 2023; 24(19):14474. https://doi.org/10.3390/ijms241914474
Chicago/Turabian StyleBoboc, Ianis Kevyn Stefan, Alexandru Cojocaru, Gabriel Nedelea, Bogdan Catalin, Maria Bogdan, and Daniela Calina. 2023. "Chronic Administration of Ion Channel Blockers Impact Microglia Morphology and Function in a Murine Model of Alzheimer’s Disease" International Journal of Molecular Sciences 24, no. 19: 14474. https://doi.org/10.3390/ijms241914474