

Effects of Dibutylphthalate and Steroid Hormone Mixture on Human Prostate Cells
 , , , , and
, , , , and 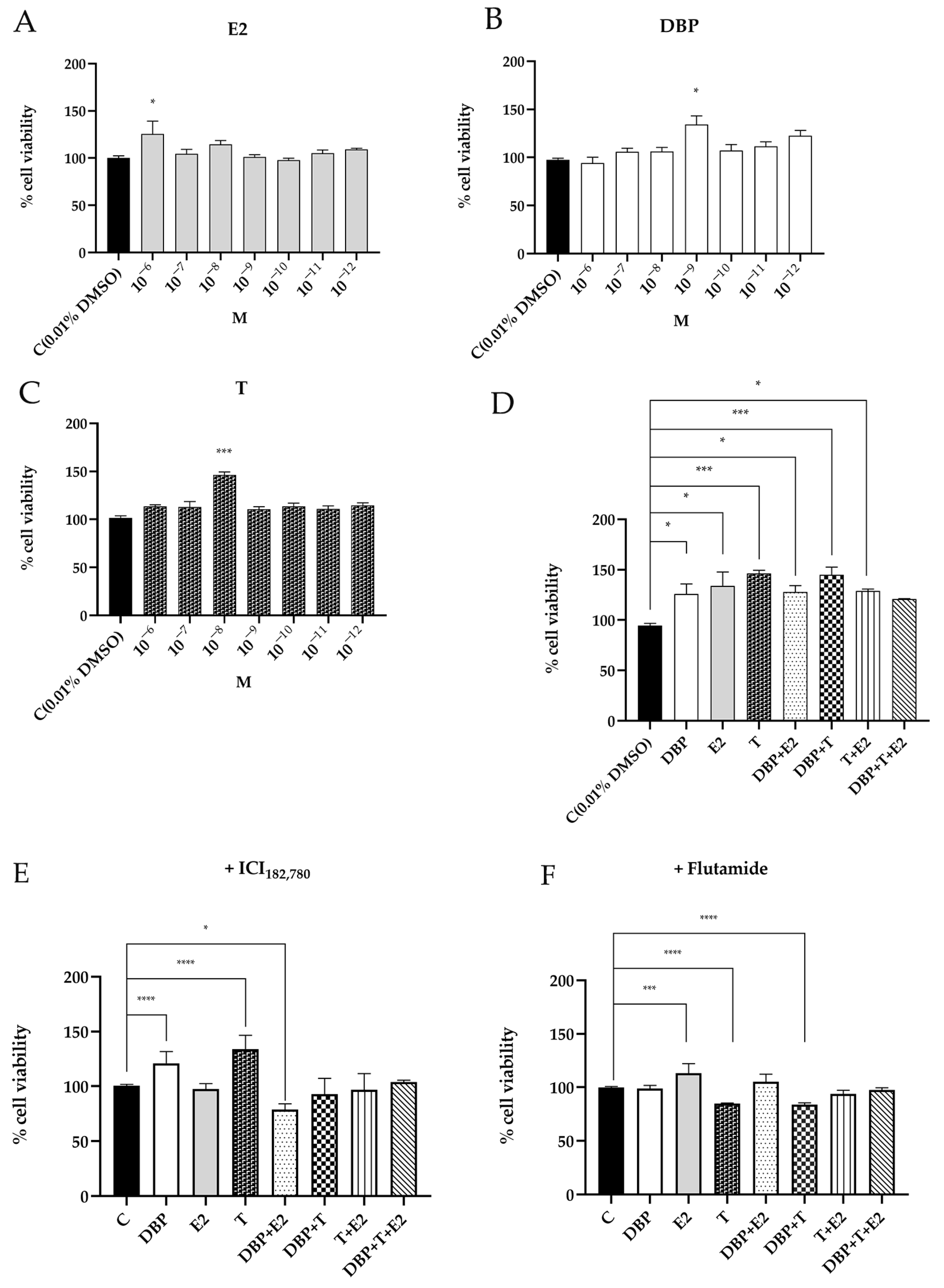{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. DBP Increases Cell Viability of Prostate Cells
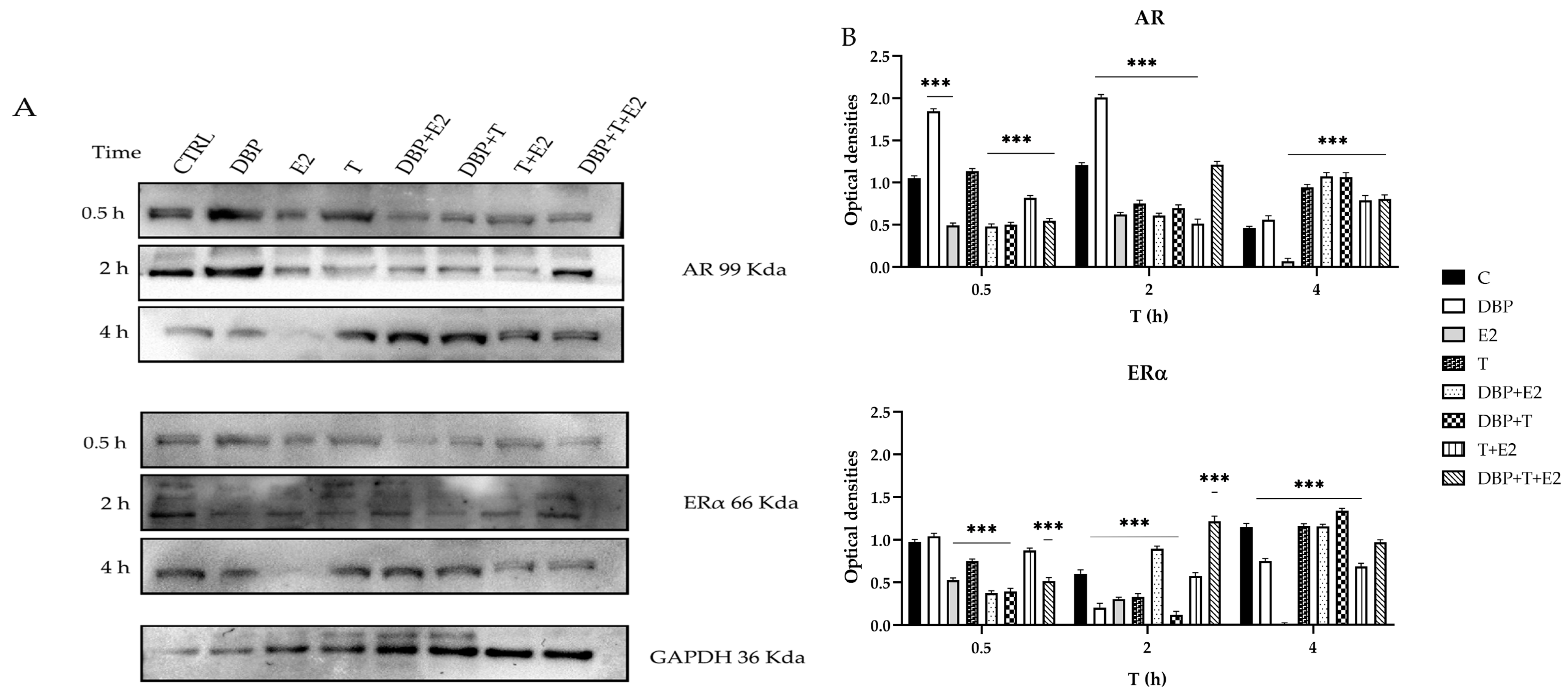
2.2. DBP Alters ERα and AR Protein Expression
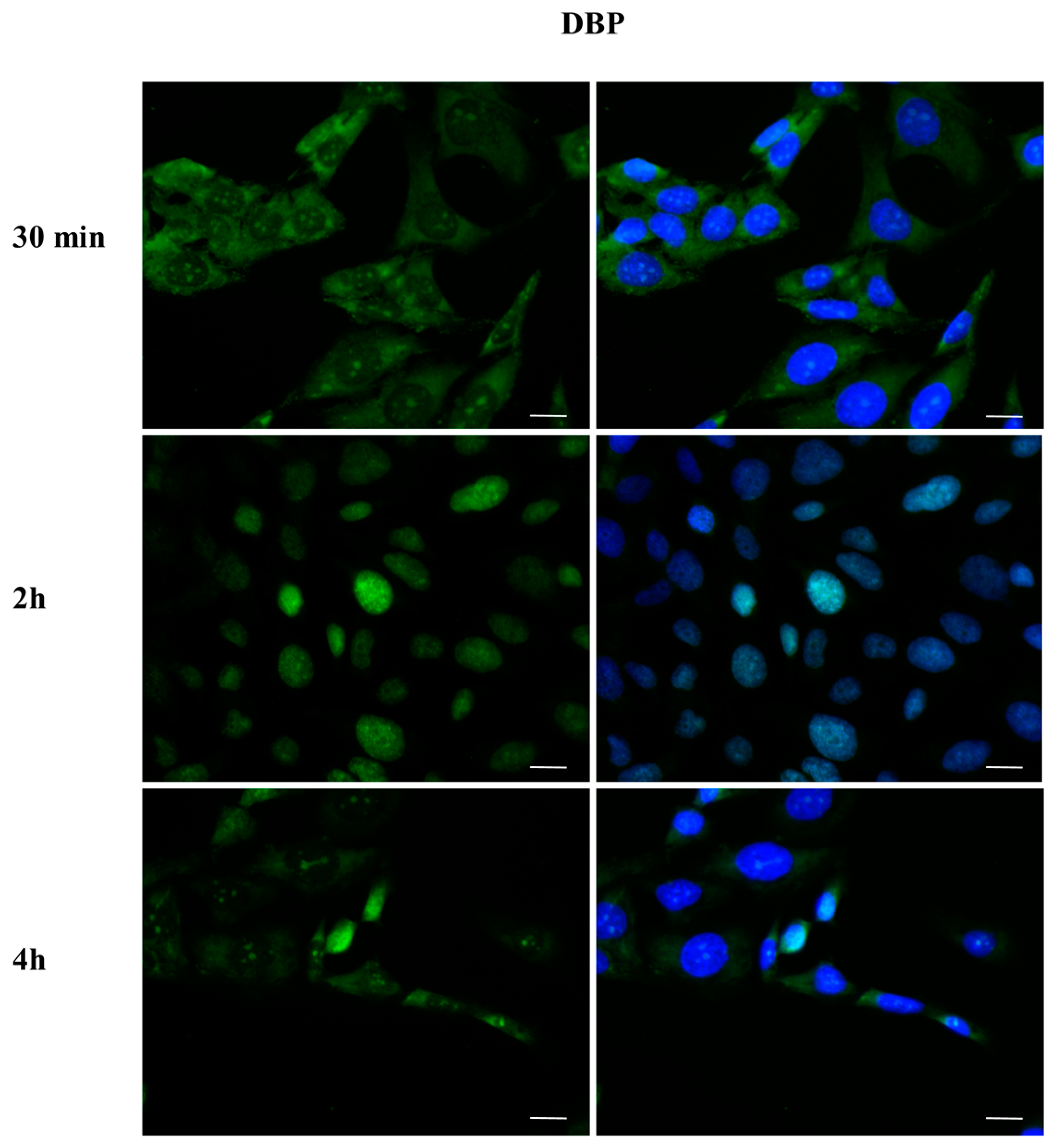
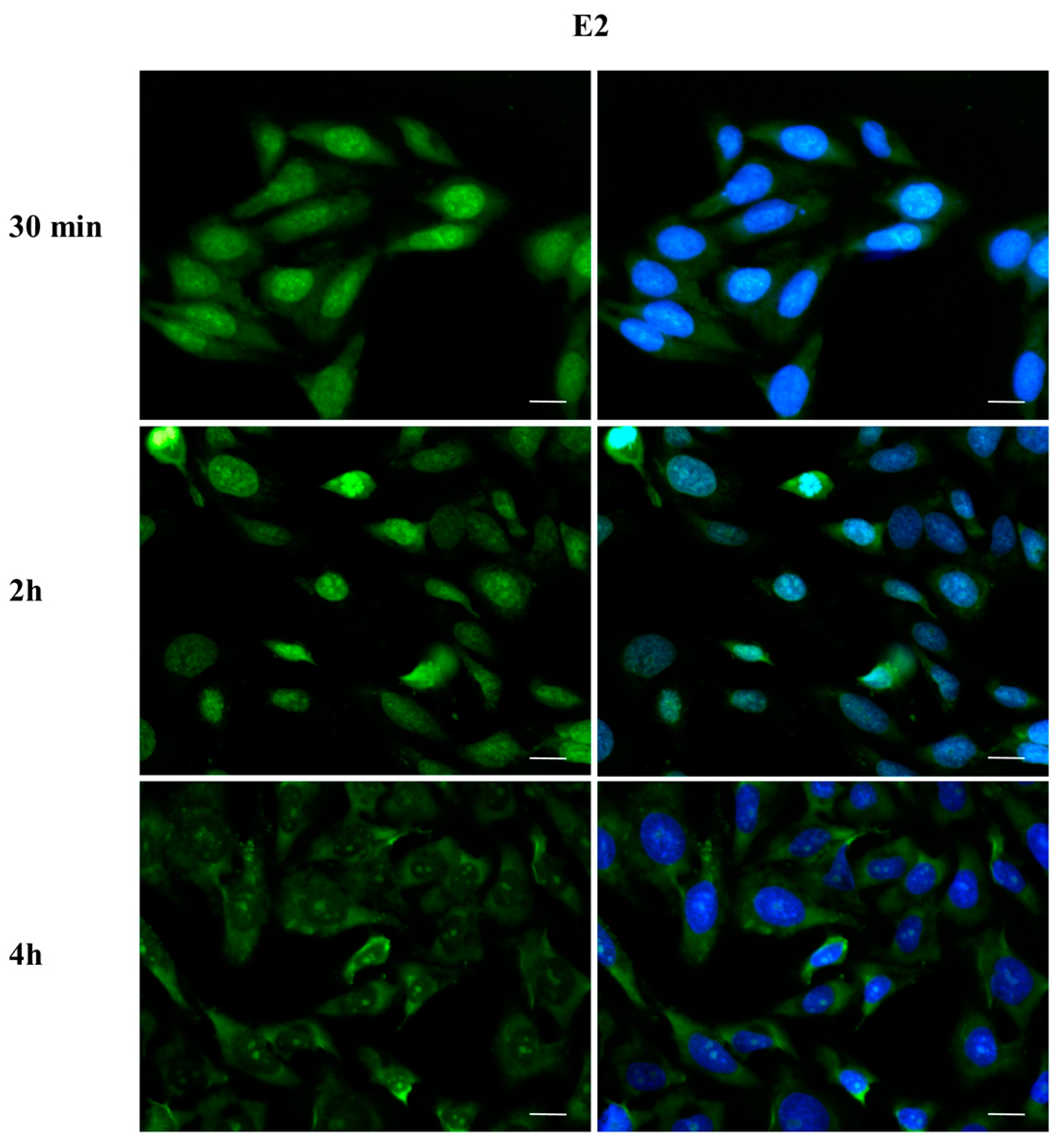
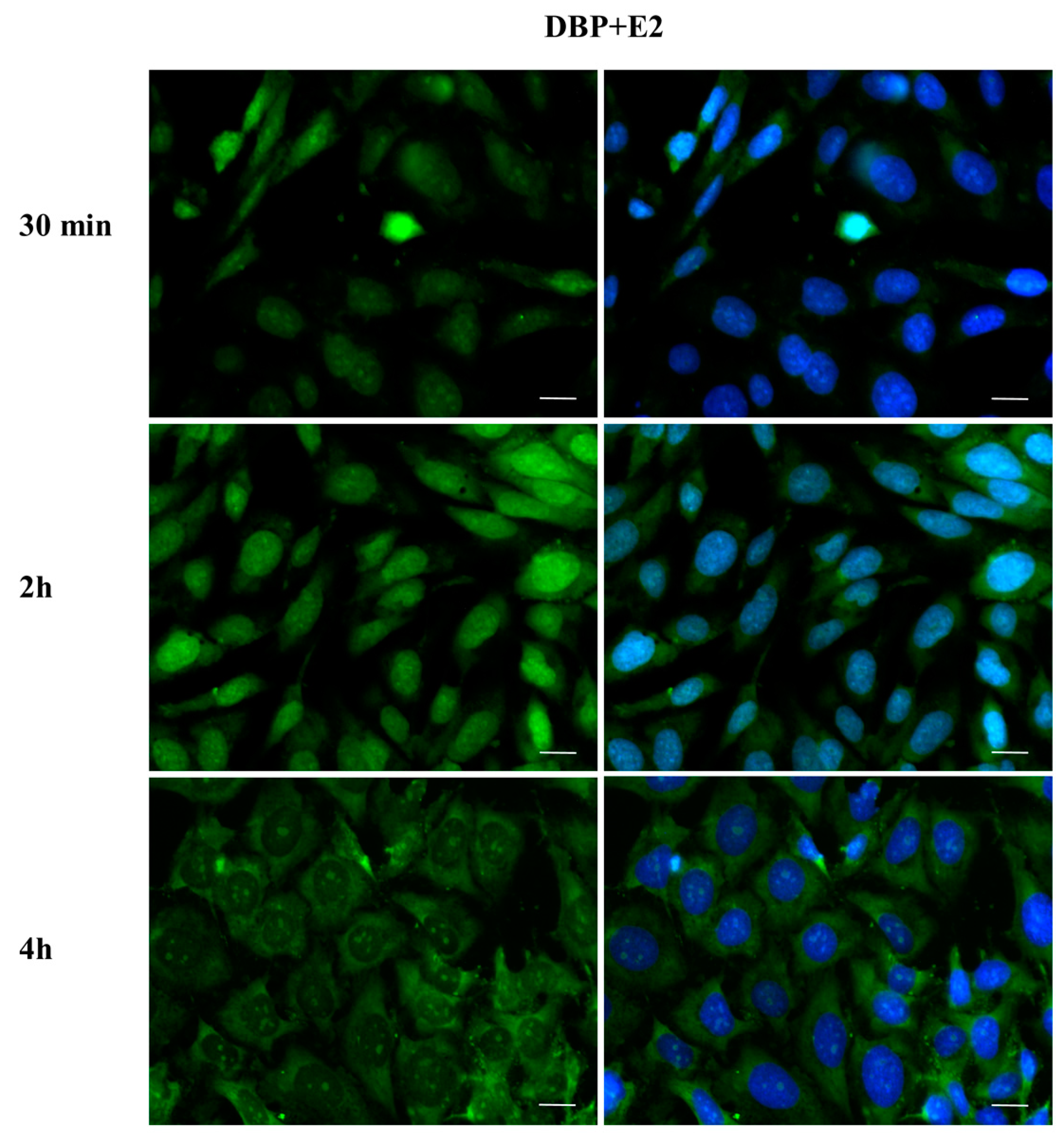
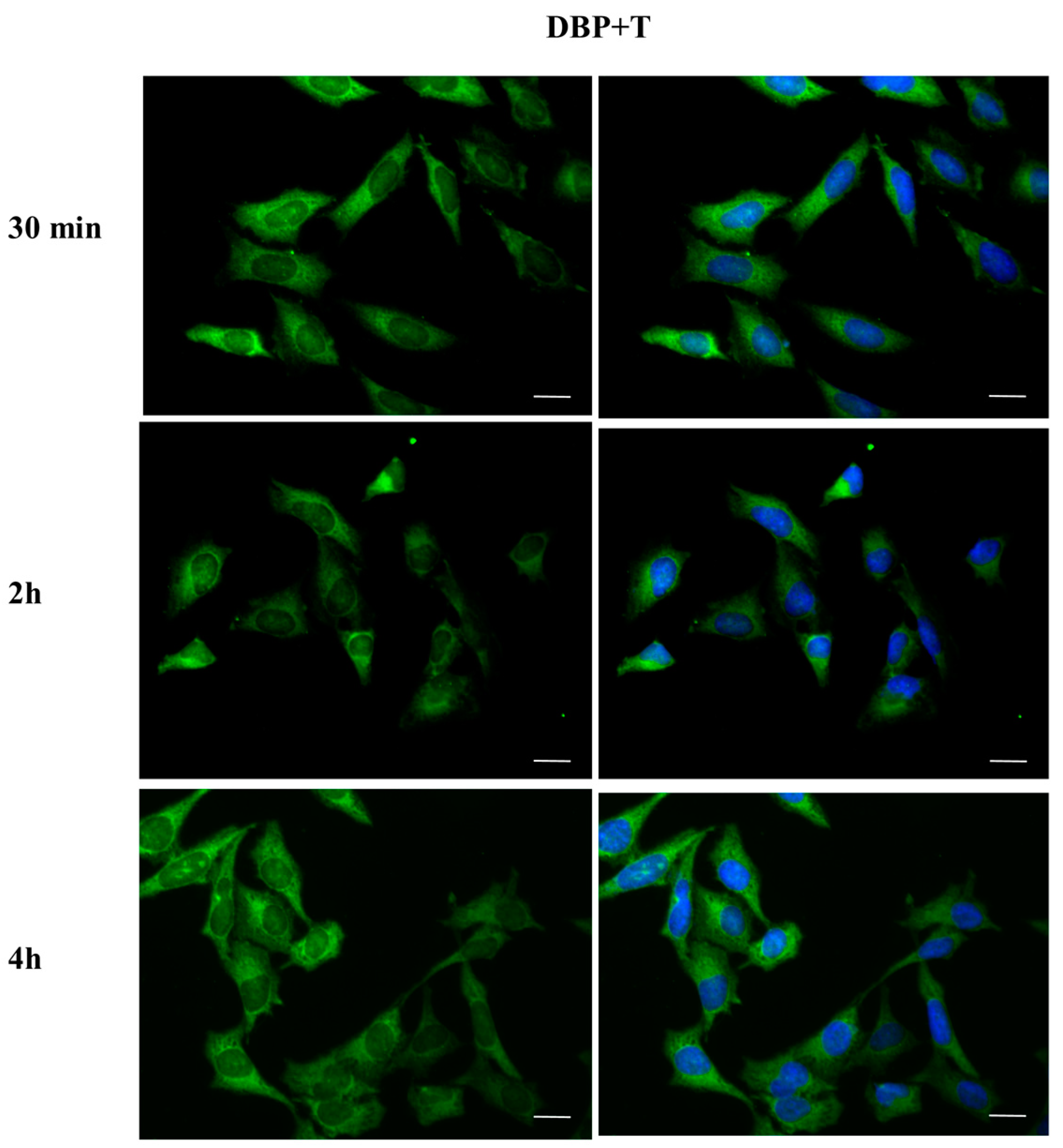
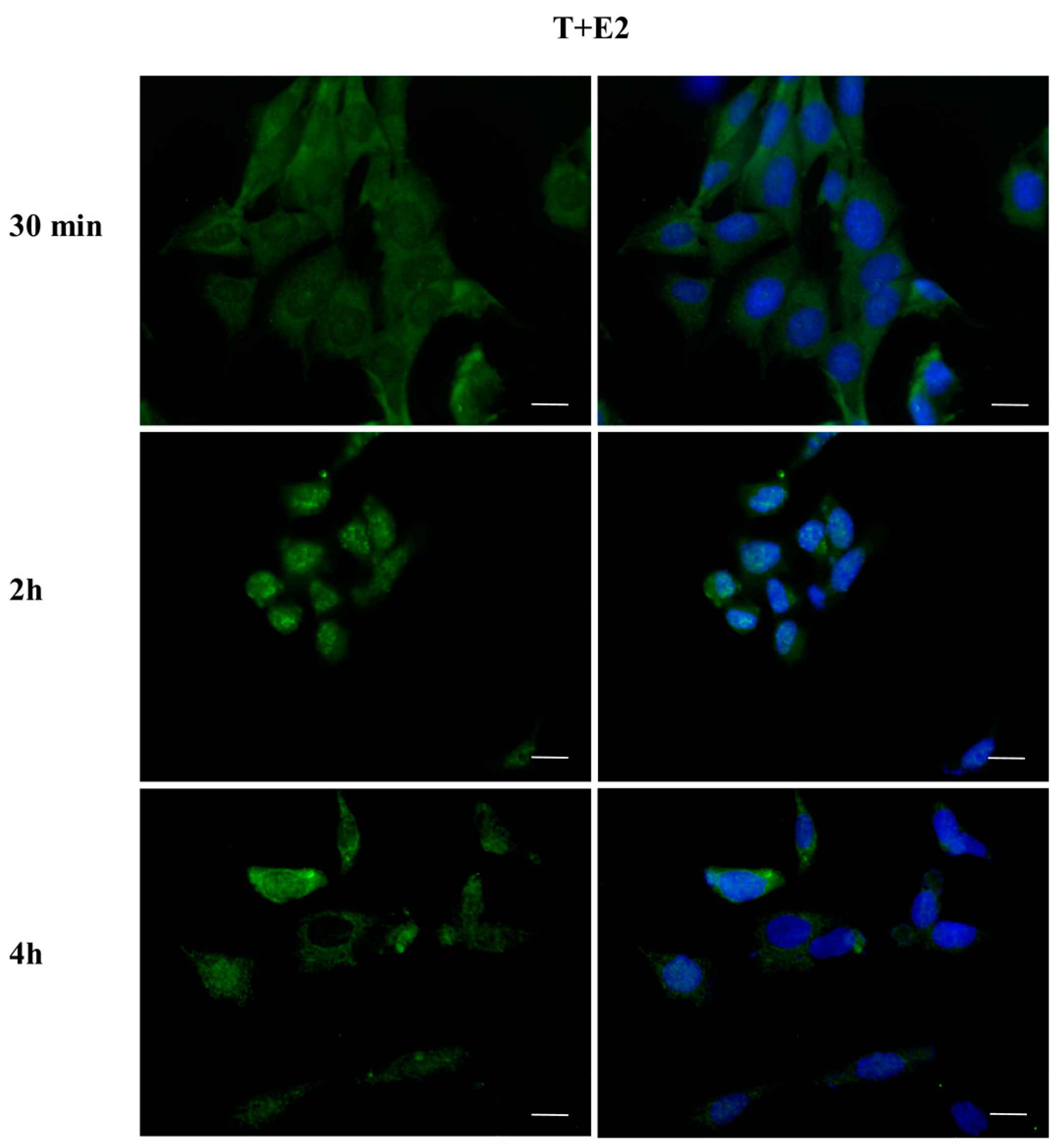
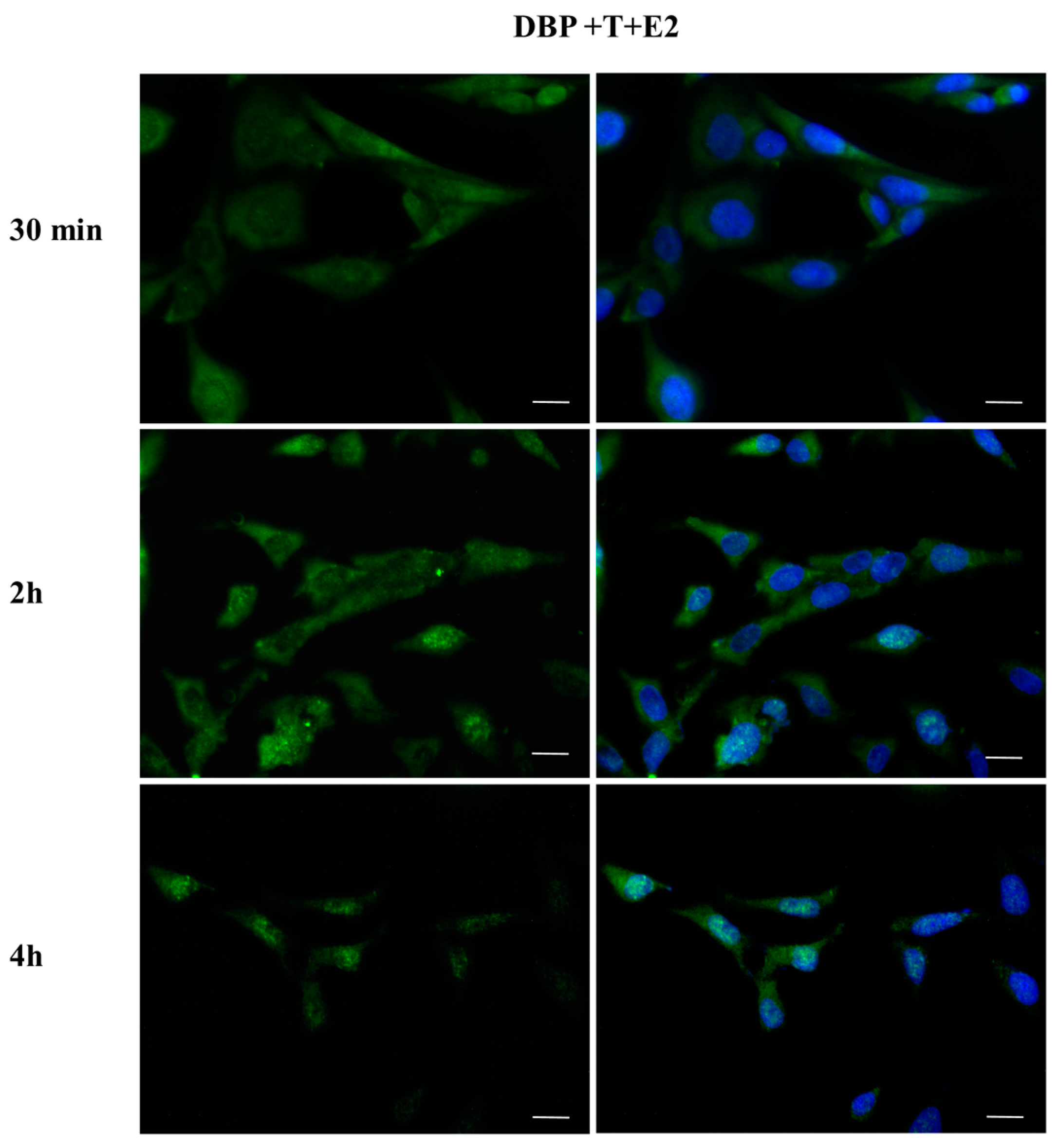
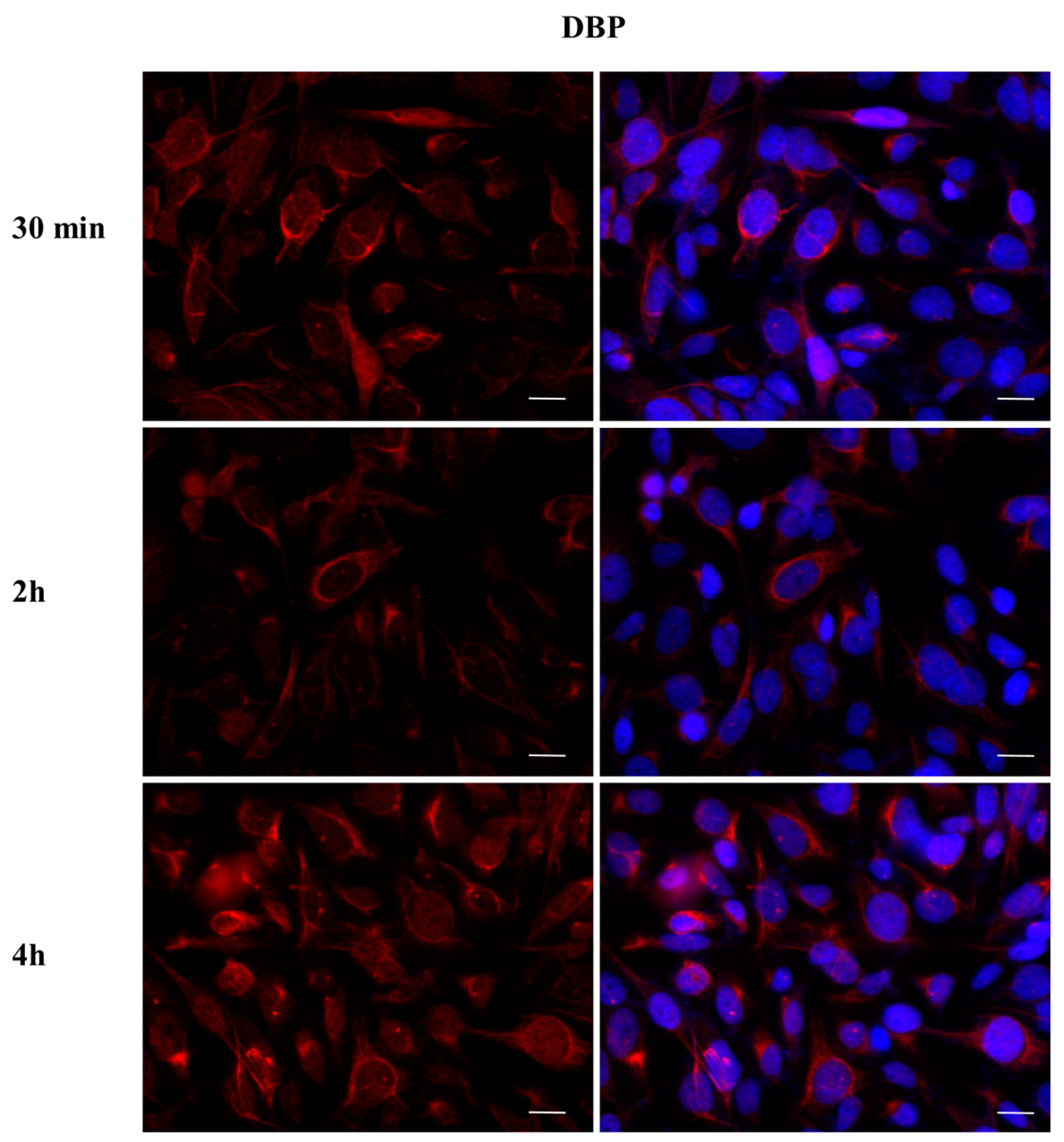
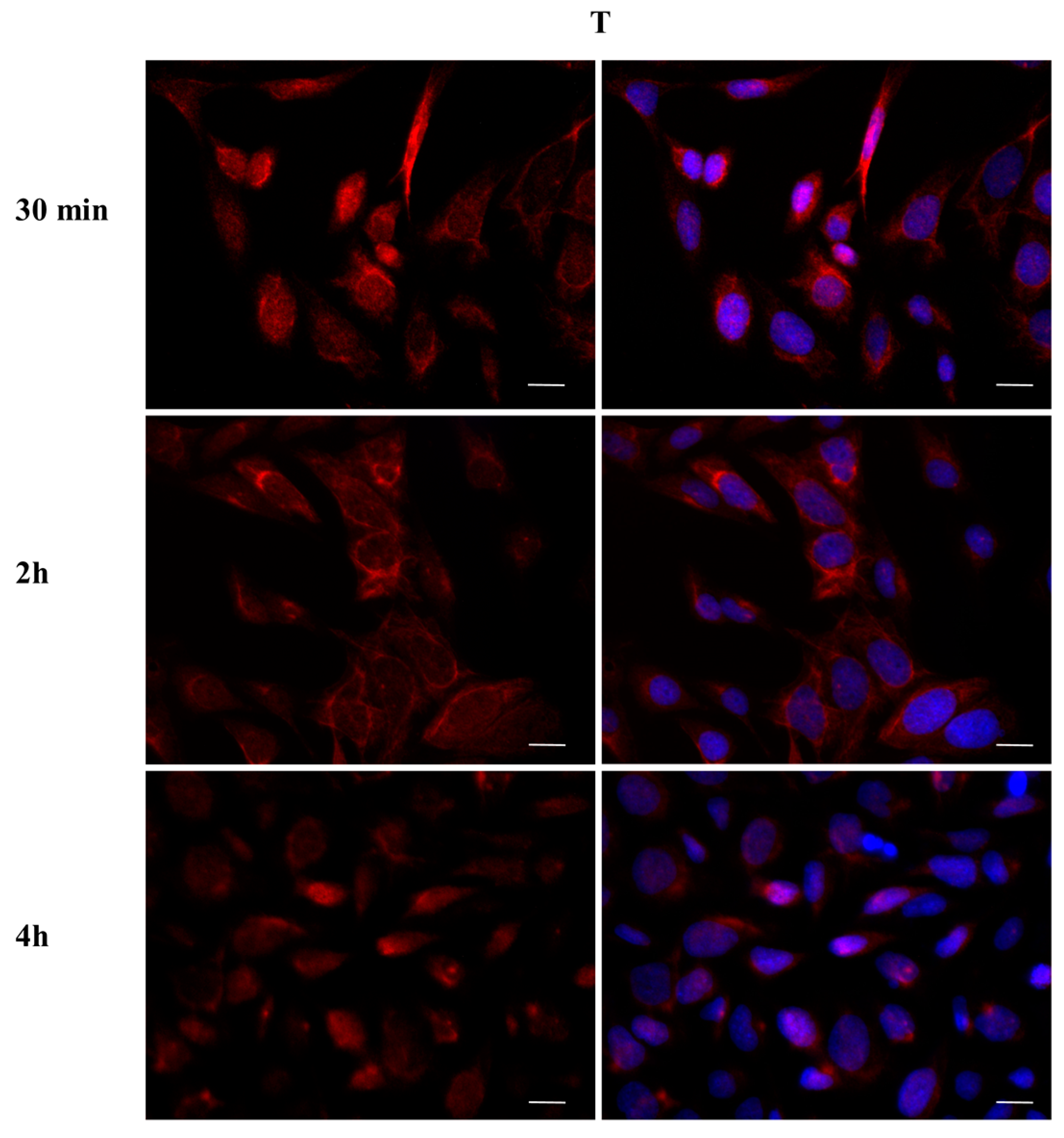
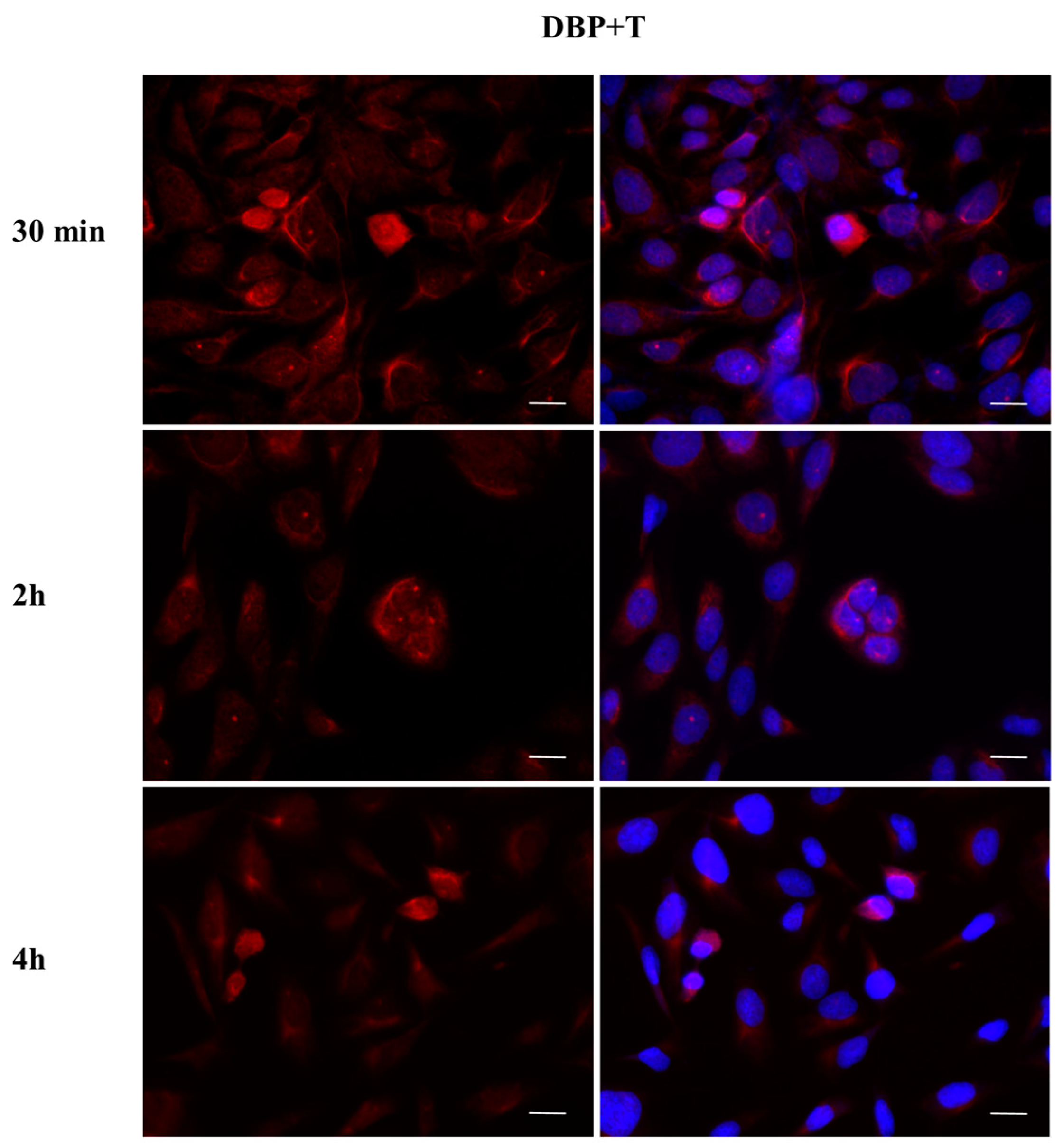
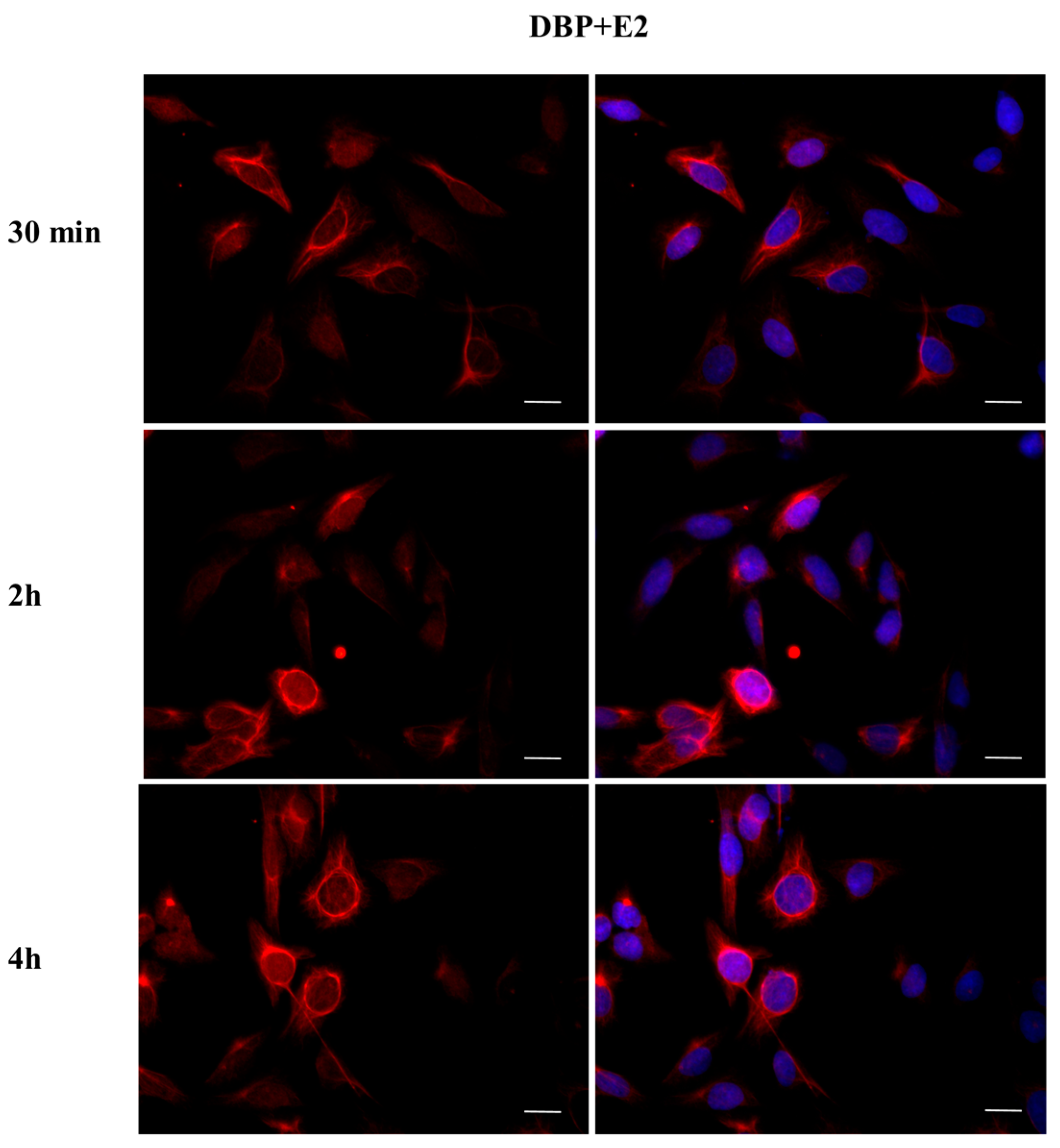
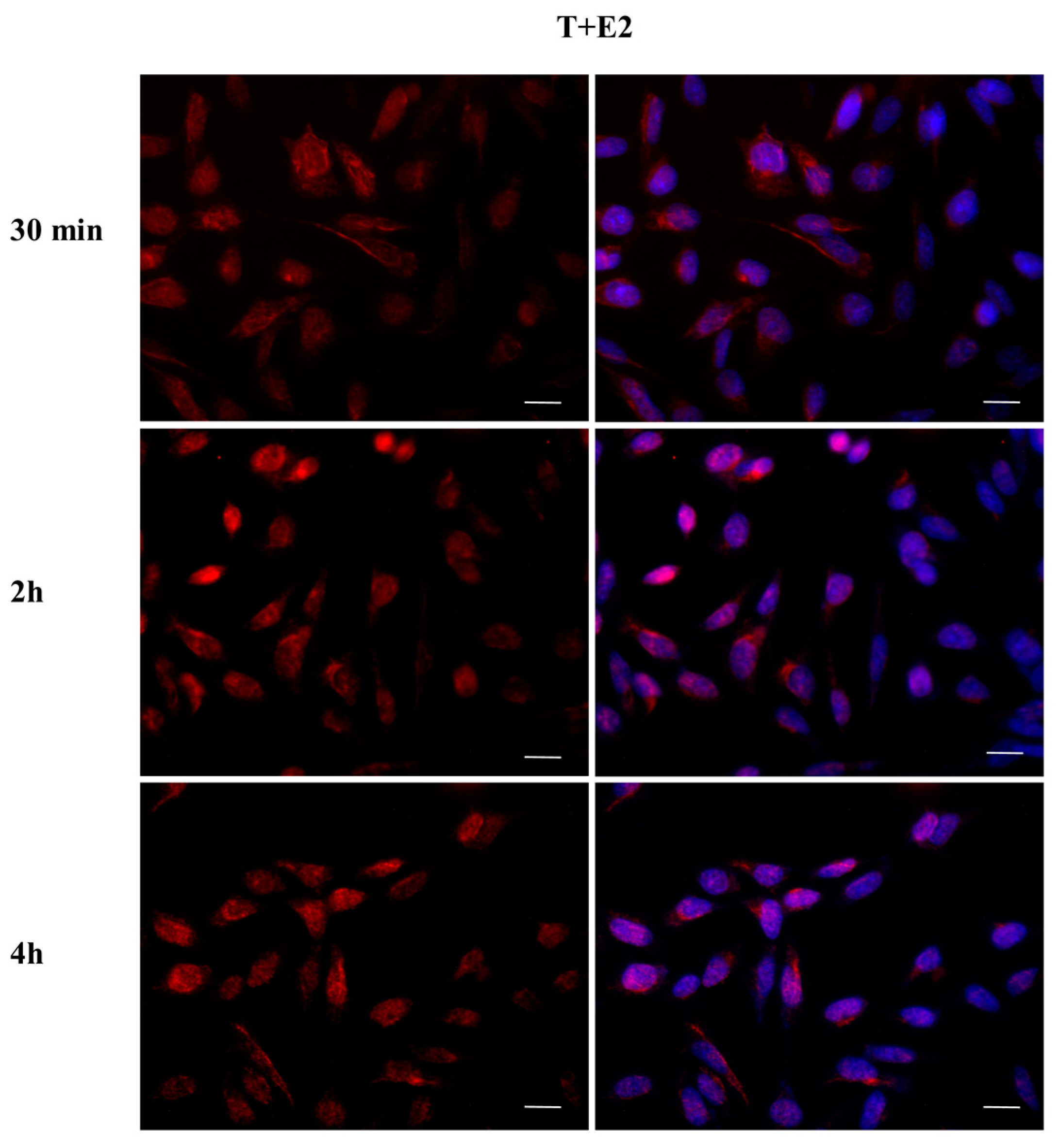
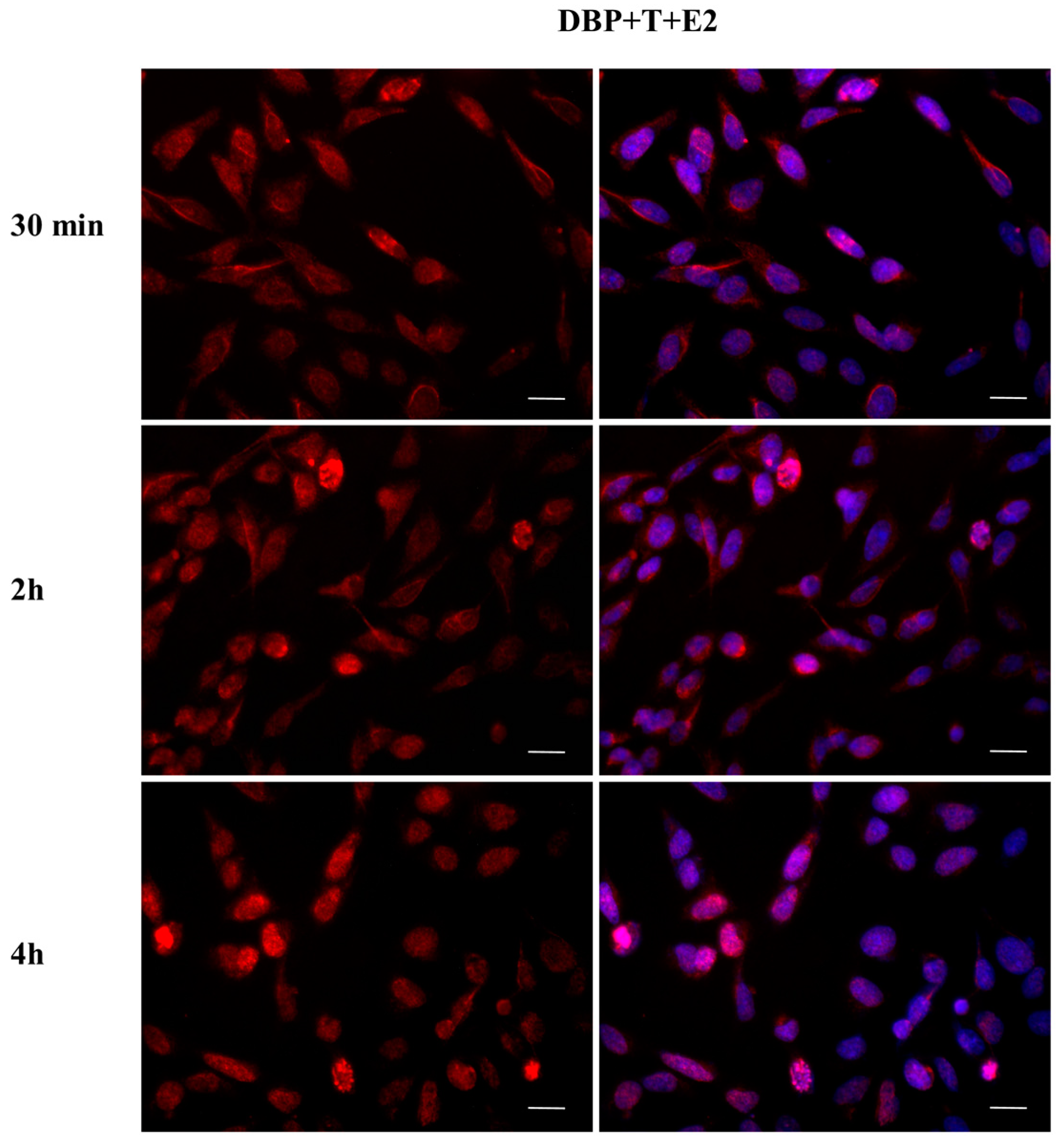
2.3. DBP Treatment Affects ERα Cellular Localization
2.4. DBP Treatment Does Not Affect AR Cellular Localization
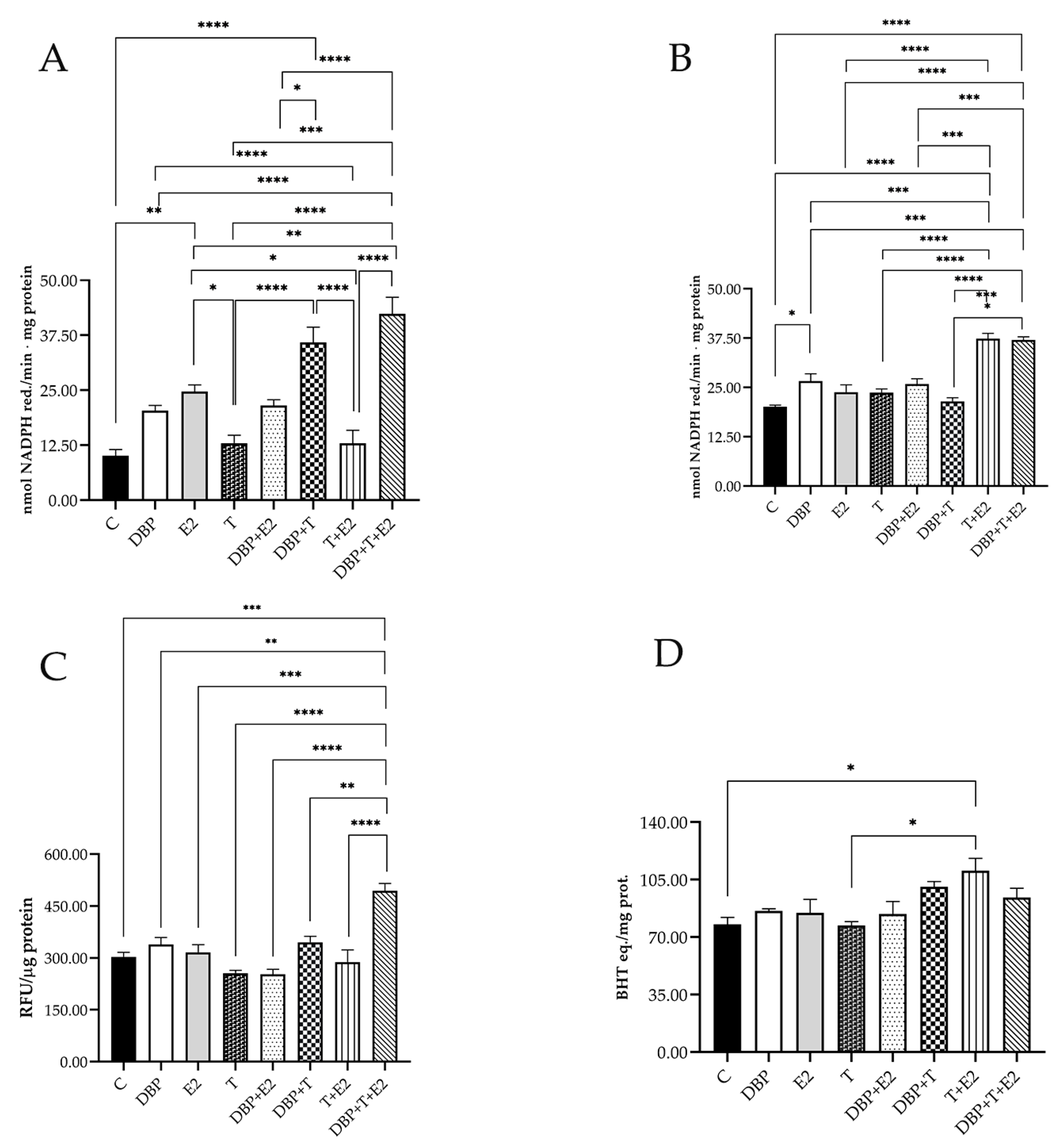
2.5. DBP Exposure Triggers Oxidative Stress
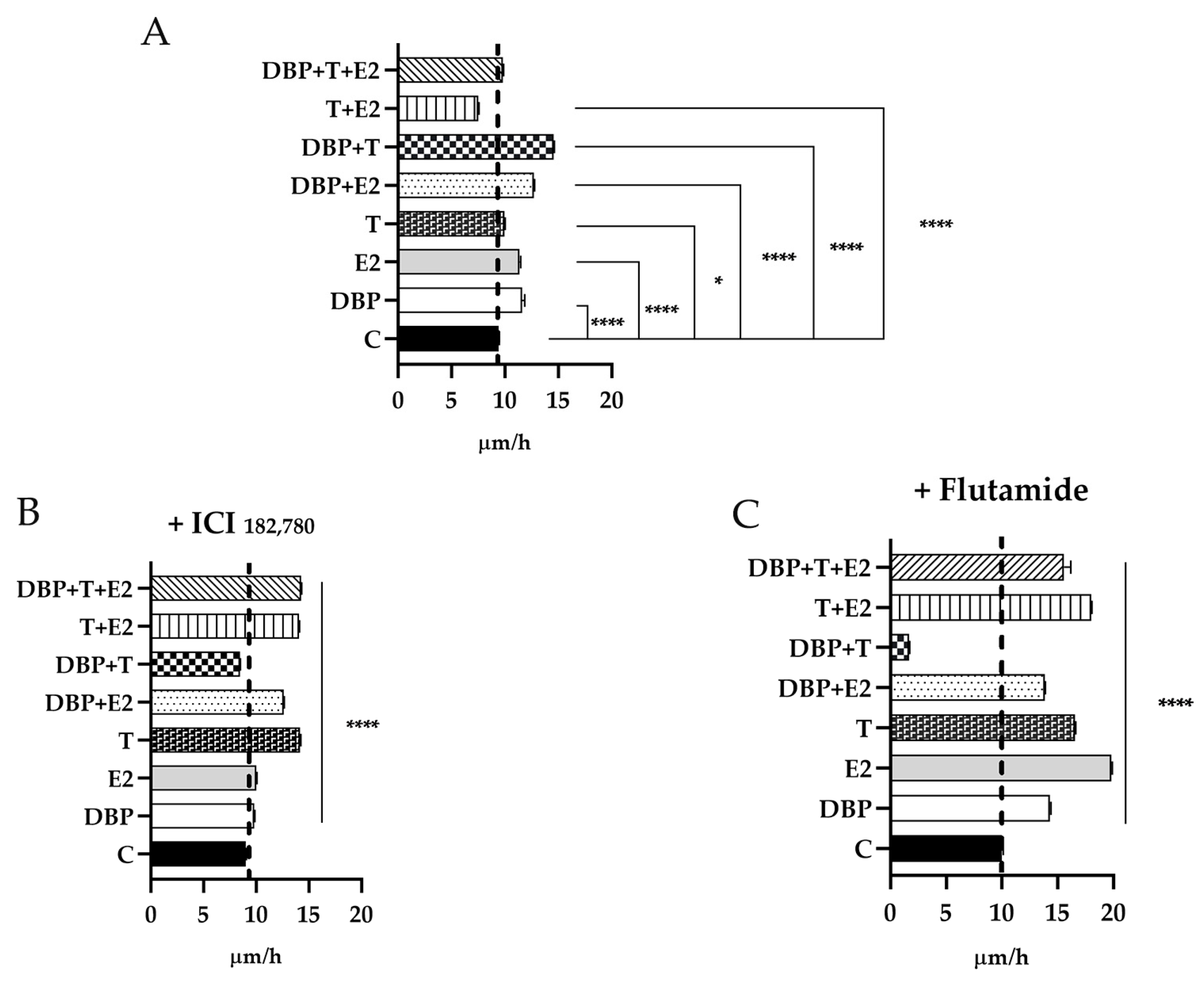
2.6. DBP Treatment Increases Cell Migration in PNT1A Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Experimental Design
4.3. Cell Culture
4.4. MTT Assay
4.5. Total Protein Extraction and Western Blotting
4.6. Immunofluorescence
4.7. Redox Homeostasis
4.8. Wound Healing Assay
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Foster, P.M.; Thomas, L.V.; Cook, M.W.; Gangolli, S.D. Study of the testicular effects and changes in zinc excretion produced by some n-alkyl phthalates in the rat. Toxicol. Appl. Pharmacol. 1980, 54, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Jiang, K.; Tang, X.; Liu, H.; Zhang, H.; Yang, X.; Nie, X.; Luo, H. Chronic exposure to di-n-butyl phthalate causes reproductive toxicity in zebrafish. J. Appl. Toxicol. 2020, 40, 1694–1703. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Sosa, J.R.; Ruiz, S.; Valdez, D.; Tullot, T. Dibutyl phthalate affects the recovery, size, and viability of pig testicular tissue ectopically grafted in immunocompromised mice. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Trovalusci, E.; Rossato, M.; Gamba, P.; Midrio, P. Testicular function and sexuality in adult patients with anorectal malformation. J. Pediatr. Surg. 2020, 55, 1839–1845. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, X.; Li, Y.; Liu, Y.; Tao, L. Exposure to Dibutyl Phthalate and Reproductive-Related Outcomes in Animal Models: Evidence From Rodents Study. Front. Physiol. 2021, 12, 684532. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Meng, X.; Liu, X.; Wang, J.; Yan, S.; Zhang, X.; Wang, M.; Ren, S.; Huang, Y. Associations of phthalates with prostate cancer among the US population. Reprod. Toxicol. 2023, 116, 108337. [Google Scholar] [CrossRef]
- Kavlock, R.; Barr, D.; Boekelheide, K.; Breslin, W.; Breysse, P.; Chapin, R.; Gaido, K.; Hodgson, E.; Marcus, M.; Shea, K.; et al. NTP-CERHR Expert Panel Update on the Reproductive and Developmental Toxicity of Di(2-ethylhexyl) phthalate. Reprod. Toxicol. 2006, 22, 291–399. [Google Scholar] [CrossRef]
- Braun, J.M.; Sathyanarayana, S.; Hauser, R. Phthalate exposure and children’s health. Curr. Opin. Pediatr. 2013, 25, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Meeker, J.D.; Sathyanarayana, S.; Swan, S.H. Phthalates and other additives in plastics: Human exposure and associated health outcomes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 2097–2113. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, M.; Forte, M.; Valiante, S.; Laforgia, V.; De Falco, M. Interference of dibutylphthalate on human prostate cell viability. Ecotoxicol. Environ. Saf. 2018, 147, 565–573. [Google Scholar] [CrossRef]
- Lacouture, A.; Lafront, C.; Peillex, C.; Pelletier, M.; Audet-Walsh, É. Impacts of endocrine-disrupting chemicals on prostate function and cancer. Environ. Res. 2021, 204, 112085. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Romero, E.; Scheringer, M. A review of phthalate pharmacokinetics in human and rat: What factors drive phthalate distribution and partitioning? Drug Metab. Rev. 2019, 51, 314–329. [Google Scholar] [CrossRef] [PubMed]
- Blount, B.C.; Milgram, K.E.; Silva, M.J.; Malek, N.A.; Reidy, J.A.; Needham, L.L.; Brock, J.W. Quantitative Detection of Eight Phthalate Metabolites in Human Urine Using HPLC-APCI-MS/MS. Anal. Chem. 2000, 72, 4127–4134. [Google Scholar] [CrossRef] [PubMed]
- Jornet-Martínez, N.; Antón-Soriano, C.; Campíns-Falcó, P. Estimation of the presence of unmetabolized dialkyl phthalates in untreated human urine by an on-line miniaturized reliable method. Sci. Total Environ. 2015, 532, 239–244. [Google Scholar] [CrossRef]
- Su, P.-H.; Chang, C.-K.; Lin, C.-Y.; Chen, H.-Y.; Liao, P.-C.; Hsiung, C.A.; Chiang, H.-C.; Wang, S.-L. Prenatal exposure to phthalate ester and pubertal development in a birth cohort in central Taiwan: A 12-year follow-up study. Environ. Res. 2015, 136, 324–330. [Google Scholar] [CrossRef]
- Santos, T.d.M.; da Silveira, L.T.R.; Rinaldi, J.C.; Scarano, W.R.; Domeniconi, R.F. Alterations in prostate morphogenesis in male rat offspring after maternal exposure to Di- n -butyl-phthalate (DBP). Reprod. Toxicol. 2017, 69, 254–264. [Google Scholar] [CrossRef]
- Wu, A.H.; Franke, A.A.; Wilkens, L.R.; Tseng, C.; Conroy, S.M.; Li, Y.; Polfus, L.M.; De Rouen, M.; Caberto, C.; Haiman, C.; et al. Urinary phthalate exposures and risk of breast cancer: The Multiethnic Cohort study. Breast Cancer Res. 2021, 23, 44. [Google Scholar] [CrossRef]
- Liu, C.; Deng, Y.-L.; Zheng, T.-Z.; Yang, P.; Jiang, X.-Q.; Liu, E.-N.; Miao, X.-P.; Wang, L.-Q.; Jiang, M.; Zeng, Q. Urinary biomarkers of phthalates exposure and risks of thyroid cancer and benign nodule. J. Hazard. Mater. 2020, 383, 121189. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, S.H. Exposure to Phthalate Esters and the Risk of Endometriosis. Dev. Reprod. 2020, 24, 71–78. [Google Scholar] [CrossRef]
- Choi, K.-C.; Park, M.-A.; Hwang, K.-A.; Lee, H.-R.; Yi, B.-R.; Jeung, E.-B. Cell growth of BG-1 ovarian cancer cells is promoted by di-n-butyl phthalate and hexabromocyclododecane via upregulation of the cyclin D and cyclin-dependent kinase-4 genes. Mol. Med. Rep. 2011, 5, 761–766. [Google Scholar] [CrossRef]
- Sun, G.; Li, Y. Exposure to DBP induces the toxicity in early development and adverse effects on cardiac development in zebrafish (Danio rerio). Chemosphere 2018, 218, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; He, X.; Song, Y.; Tu, Y.; Chen, W.; Yang, G. Sporoderm-broken spores of Ganoderma lucidum alleviates liver injury induced by DBP and BaP co-exposure in rat. Ecotoxicol. Environ. Saf. 2022, 241, 113750. [Google Scholar] [CrossRef]
- Wang, X.; Yan, X.; Yang, Y.; Yang, W.; Zhang, Y.; Wang, J.; Ye, D.; Wu, Y.; Ma, P.; Yan, B. Dibutyl phthalate-mediated oxidative stress induces splenic injury in mice and the attenuating effects of vitamin E and curcumin. Food Chem. Toxicol. 2020, 136, 110955. [Google Scholar] [CrossRef]
- Jiang, N.; Song, P.; Li, X.; Zhu, L.; Wang, J.; Yin, X.; Wang, J. Dibutyl phthalate induced oxidative stress and genotoxicity on adult zebrafish (Danio rerio) brain. J. Hazard. Mater. 2021, 424, 127749. [Google Scholar] [CrossRef] [PubMed]
- Bu, H.; Tang, S.; Liu, G.; Miao, C.; Zhou, X.; Yang, H.; Liu, B. In silico, in vitro and in vivo studies: Dibutyl phthalate promotes prostate cancer cell proliferation by activating Forkhead Box M1 and remission after Natura-α pretreatment. Toxicology 2023, 488, 153465. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, S.; Masai, E.; Kamimura, N.; Takahashi, K.; Anderson, R.C.; Faisal, P.A. Phthalates impact human health: Epidemiological evidences and plausible mechanism of action. J. Hazard. Mater. 2017, 340, 360–383. [Google Scholar] [CrossRef]
- Hauser, R.; Skakkebaek, N.E.; Hass, U.; Toppari, J.; Juul, A.; Andersson, A.-M.; Kortenkamp, A.; Heindel, J.J.; Trasande, L. Male Reproductive Disorders, Diseases, and Costs of Exposure to Endocrine-Disrupting Chemicals in the European Union. J. Clin. Endocrinol. Metab. 2015, 100, 1267–1277. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, W.; Zhang, J.; Li, H. Role of JNK and ERK1/2 MAPK signaling pathway in testicular injury of rats induced by di-N-butyl-phthalate (DBP). Biol. Res. 2019, 52, 41. [Google Scholar] [CrossRef]
- Nassan, F.L.; Coull, B.A.; Skakkebaek, N.E.; Williams, M.A.; Dadd, R.; Mínguez-Alarcón, L.; Krawetz, S.A.; Hait, E.J.; Korzenik, J.R.; Moss, A.C.; et al. A crossover–crossback prospective study of dibutyl-phthalate exposure from mesalamine medications and semen quality in men with inflammatory bowel disease. Environ. Int. 2016, 95, 120–130. [Google Scholar] [CrossRef]
- Foster, P.M.D.; Mylchreest, E.; Gaido, K.W.; Sar, M. Effects of phthalate esters on the developing reproductive tract of male rats. Hum. Reprod. Updat. 2001, 7, 231–235. [Google Scholar] [CrossRef]
- Mylchreest, E.; Wallace, D.G.; Cattley, R.C.; Foster, P.M.D. Dose-dependent alterations in androgen-regulated male reproductive development in rats exposed to Di(n-butyl) phthalate during late gestation. Toxicol. Sci. 2000, 55, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Kim, T.S.; Shin, J.-H.; Moon, H.J.; Kang, I.H.; Kim, I.Y.; Oh, J.Y.; Han, S.Y. Neonatal exposure to di(n-butyl) phthalate (DBP) alters male reproductive-tract development. J. Toxicol. Environ. Health Part A 2004, 67, 2045–2060. [Google Scholar] [CrossRef]
- Scarano, W.R.; Toledo, F.C.; Guerra, M.T.; Pinheiro, P.F.F.; Domeniconi, R.F.; Felisbino, S.L.; Campos, S.G.P.; Taboga, S.R.; Kempinas, W.G. Functional and Morphological Reproductive Aspects in Male Rats Exposed to Di-n-Butyl Phthalate (DBP) in Utero and During Lactation. J. Toxicol. Environ. Health Part A 2010, 73, 972–984. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Ma, L.; Yuan, L.; Wang, X.; Zhang, W. Study on developmental abnormalities in hypospadiac male rats induced by maternal exposure to di-n-butyl phthalate (DBP). Toxicology 2007, 232, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.-T.; Sun, W.-L.; Jing, Y.-F.; Liu, S.-B.; Ma, Z.; Hong, Y.; Ma, L.; Qin, C.; Liu, Q.; Stratton, H.J.; et al. Prenatal exposure to di-n-butyl phthalate induces anorectal malformations in male rat offspring. Toxicology 2011, 290, 322–326. [Google Scholar] [CrossRef]
- Bello, U.M.; Madekurozwa, M.-C.; Groenewald, H.B.; Aire, T.A.; Arukwe, A. The effects on steroidogenesis and histopathology of adult male Japanese quails (Coturnix coturnix japonica) testis following pre-pubertal exposure to di(n-butyl) phthalate (DBP). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2014, 166, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, A.R.; Santos, T.M.; Brandt, J.Z.; Delella, F.K.; Gonçalves, B.F.; Campos, S.G.P.; Taboga, S.R.; Favaro, W.J.; Domeniconi, R.F.; Scarano, W.R. Gestational and lactational exposition to Di-N-butyl-phthalate (DBP) increases inflammation and preneoplastic lesions in prostate of wistar rats after carcinogenic N-methyl-N-nitrosourea (MNU) plus testosterone protocol. Environ. Toxicol. 2015, 31, 1185–1195. [Google Scholar] [CrossRef]
- Adeniyi, A.A.; Okedeyi, O.O.; Yusuf, K.A. Flame ionization gas chromatographic determination of phthalate esters in water, surface sediments and fish species in the Ogun river catchments, Ketu, Lagos, Nigeria. Environ. Monit. Assess. 2010, 172, 561–569. [Google Scholar] [CrossRef]
- Available online: https://www.cpsc.gov/Business--Manufacturing/Business-Education/Business-Guidance/Phthalates-Information (accessed on 20 July 2023).
- Ventrice, P.; Ventrice, D.; Russo, E.; De Sarro, G. Phthalates: European regulation, chemistry, pharmacokinetic and related toxicity. Environ. Toxicol. Pharmacol. 2013, 36, 88–96. [Google Scholar] [CrossRef]
- Kassab, R.B.; Lokman, M.S.; Essawy, E.A. Neurochemical alterations following the exposure to di-n-butyl phthalate in rats. Metab Brain Dis. 2019, 34, 235–244. [Google Scholar] [CrossRef]
- Karabulut, G.; Barlas, N. Genotoxic, histologic, immunohistochemical, morphometric and hormonal effects of di-(2-ethylhexyl)-phthalate (DEHP) on reproductive systems in pre-pubertal male rats. Toxicol. Res. 2018, 7, 859–873. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, S.; Iida, M.; Kobayashi, S.; Jin, K.; Matsuda, T.; Kojima, H. Differential effects of phthalate esters on transcriptional activities via human estrogen receptors α and β, and androgen receptor. Toxicology 2005, 210, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.-H.; Tsai, Y.-S.; Wang, J.-Y.; Chen, H.-L.; Yang, W.-H.; Lee, C.-C. Sex hormones and oxidative stress mediated phthalate-induced effects in prostatic enlargement. Environ. Int. 2019, 126, 184–192. [Google Scholar] [CrossRef] [PubMed]
- La Vignera, S.; Condorelli, R.A.; Russo, G.I.; Morgia, G.; Calogero, A.E. Endocrine control of benign prostatic hyperplasia. Andrology 2016, 4, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, T.M.; Ricke, W.A. Androgens and estrogens in benign prostatic hyperplasia: Past, present and future. Differentiation 2011, 82, 184–199. [Google Scholar] [CrossRef]
- Roehrborn, C.G. Pathology of benign prostatic hyperplasia. Int. J. Impot. Res. 2008, 20, S11–S18. [Google Scholar] [CrossRef]
- Engel, A.; Buhrke, T.; Imber, F.; Jessel, S.; Seidel, A.; Völkel, W.; Lampen, A. Agonistic and antagonistic effects of phthalates and their urinary metabolites on the steroid hormone receptors ERα, ERβ, and AR. Toxicol. Lett. 2017, 277, 54–63. [Google Scholar] [CrossRef]
- Zhu, Q.; Liu, L.; Zhou, X.; Ma, M. In silico study of molecular mechanisms of action: Estrogenic disruptors among phthalate esters. Environ. Pollut. 2019, 255 Pt 1, 113193. [Google Scholar] [CrossRef]
- Ho, C.K.M.; Habib, F.K. Estrogen and androgen signaling in the pathogenesis of BPH. Nat. Rev. Urol. 2011, 8, 29–41. [Google Scholar] [CrossRef]
- Asiedu, B.; Anang, Y.; Nyarko, A.; Doku, D.A.; Amoah, B.Y.; Santa, S.; Ngala, R.A.; Asare, G.A. The role of sex steroid hormones in benign prostatic hyperplasia. Aging Male 2017, 20, 17–22. [Google Scholar] [CrossRef]
- Zhou, P.; Wu, S.; Huang, D.; Wang, K.; Su, X.; Yang, R.; Shao, C.; Wu, J. Oral exposure to DEHP may stimulate prostatic hyperplasia associated with upregulation of COX-2 and L-PGDS expressions in male adult rats. Reprod. Toxicol. 2022, 112, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Cunha, G.R.; Ricke, W.; Thomson, A.; Marker, P.C.; Risbridger, G.; Hayward, S.W.; Wang, Y.; Donjacour, A.A.; Kurita, T. Hormonal, cellular, and molecular regulation of normal and neoplastic prostatic development. J. Steroid Biochem. Mol. Biol. 2004, 92, 221–236. [Google Scholar] [CrossRef] [PubMed]
- Cavalca, A.M.B.; Aquino, A.M.; Mosele, F.C.; Justulin, L.A.; Delella, F.K.; Flaws, J.A.; Scarano, W.R. Effects of a phthalate metabolite mixture on both normal and tumoral human prostate cells. Environ. Toxicol. 2022, 37, 2566–2578. [Google Scholar] [CrossRef] [PubMed]
- Gandaglia, G.; Leni, R.; Bray, F.; Fleshner, N.; Freedland, S.J.; Kibel, A.; Stattin, P.; Van Poppel, H.; La Vecchia, C. Epidemiology and Prevention of Prostate Cancer. Eur. Urol. Oncol. 2021, 4, 877–892. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Scarano, W.R.; de Toledo, F.C.; Guerra, M.T.; de Campos, S.G.P.; Júnior, L.A.J.; Felisbino, S.L.; Anselmo-Franci, J.A.; Taboga, S.R.; Kempinas, W.D.G. Long-term effects of developmental exposure to di-n-butyl-phthalate (DBP) on rat prostate: Proliferative and inflammatory disorders and a possible role of androgens. Toxicology 2009, 262, 215–223. [Google Scholar] [CrossRef]
- Santos, P.M.G.; Barsky, A.R.; Hwang, W.; Deville, C.; Wang, X.; Both, S.; Bekelman, J.E.; Christodouleas, J.P.; Vapiwala, N. Comparative toxicity outcomes of proton-beam therapy versus intensity-modulated radiotherapy for prostate cancer in the postoperative setting. Cancer 2019, 125, 4278–4293. [Google Scholar] [CrossRef]
- Qie, Y.; Zhou, D.; Wu, Z.; Liu, S.; Shen, C.; Hu, H.; Zhang, C.; Xu, Y. Low-dose hexavalent chromium(VI) exposure promotes prostate cancer cell proliferation by activating MAGEB2-AR signal pathway. Ecotoxicol. Environ. Saf. 2022, 241, 113724. [Google Scholar] [CrossRef]
- Li, H.; Hammarstrand, S.; Midberg, B.; Xu, Y.; Li, Y.; Olsson, D.S.; Fletcher, T.; Jakobsson, K.; Andersson, E.M. Cancer incidence in a Swedish cohort with high exposure to perfluoroalkyl substances in drinking water. Environ. Res. 2021, 204, 112217. [Google Scholar] [CrossRef]
- Dai, C.; Heemers, H.; Sharifi, N. Androgen Signaling in Prostate Cancer. Cold Spring Harb. Perspect. Med. 2017, 7, a030452. [Google Scholar] [CrossRef]
- Lee, H.-R.; Hwang, K.-A.; Choi, K.-C. The estrogen receptor signaling pathway activated by phthalates is linked with transforming growth factor-β in the progression of LNCaP prostate cancer models. Int. J. Oncol. 2014, 45, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Yong, W.; Jiao, C.; Jianhui, W.; Yan, Z.; Qi, P.; Xiu, W.; Zuyue, S.; Yunhui, Z. Mono-2-ethyhexyl phthalate advancing the progression of prostate cancer through activating the hedgehog pathway in LNCaP cells. Toxicol. Vitr. 2016, 32, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Huang, C.; Ma, X.; Wu, R.; Zhu, W.; Li, X.; Liang, Z.; Deng, F.; Wu, J.; Geng, S.; et al. Phthalates promote prostate cancer cell proliferation through activation of ERK5 and p38. Environ. Toxicol. Pharmacol. 2018, 63, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Chimento, A.; Sirianni, R.; Delalande, C.; Silandre, D.; Bois, C.; Andò, S.; Maggiolini, M.; Carreau, S.; Pezzi, V. 17β-Estradiol activates rapid signaling pathways involved in rat pachytene spermatocytes apoptosis through GPR30 and ERα. Mol. Cell. Endocrinol. 2010, 320, 136–144. [Google Scholar] [CrossRef]
- Zhang, T.; Wu, J.; Zhang, X.; Zhou, X.; Wang, S.; Wang, Z. Pharmacophore based in silico study with laboratory verification—Environmental explanation of prostate cancer recurrence. Environ. Sci. Pollut. Res. Int. 2021, 28, 61581–61591. [Google Scholar] [CrossRef]
- Mathieu-Denoncourt, J.; Wallace, S.J.; de Solla, S.R.; Langlois, V.S. Plasticizer endocrine disruption: Highlighting developmental and reproductive effects in mammals and non-mammalian aquatic species. Gen. Comp. Endocrinol. 2015, 219, 74–88. [Google Scholar] [CrossRef]
- Sedha, S.; Kumar, S.; Shukla, S. Role of Oxidative Stress in Male Reproductive Dysfunctions with Reference to Phthalate Compounds. Urol. J. 2015, 12, 2304–2316. [Google Scholar] [CrossRef]
- Sobarzo, C.M.; Rosana, N.d.M.; Livia, L.; Berta, D.; Schteingart, H.F. Mono-(2-ethylhexyl) phthalate (MEHP) affects intercellular junctions of Sertoli cell: A potential role of oxidative stress. Reprod. Toxicol. 2015, 58, 203–212. [Google Scholar] [CrossRef]
- Udensi, U.K.; Tchounwou, P.B. Oxidative stress in prostate hyperplasia and carcinogenesis. J. Exp. Clin. Cancer Res. 2016, 35, 139. [Google Scholar] [CrossRef]
- Erkekoğlu, P.; Rachidi, W.; De Rosa, V.; Giray, B.; Favier, A.; Hıncal, F. Protective effect of selenium supplementation on the genotoxicity of di(2-ethylhexyl)phthalate and mono(2-ethylhexyl)phthalate treatment in LNCaP cells. Free Radic. Biol. Med. 2010, 49, 559–566. [Google Scholar] [CrossRef]
- Thomas, S.; Ricke, W.A.; Li, L. Toxicoproteomics of Mono(2-ethylhexyl) phthalate and Perfluorooctanesulfonic Acid in Models of Prostatic Diseases. Chem. Res. Toxicol. 2023, 36, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, Z.; Peng, Z.; Song, P.; Zhou, J.; Wang, L.; Chen, J.; Dong, Q. Exposure to Di-2-ethylhexyl Phthalate and Benign Prostatic Hyperplasia, NHANES 2001-2008. Front. Endocrinol. 2022, 12, 804457. [Google Scholar] [CrossRef]
- Bonkhoff, H.; Fixemer, T.; Hunsicker, I.; Remberger, K. Estrogen Receptor Expression in Prostate Cancer and Premalignant Prostatic Lesions. Am. J. Pathol. 1999, 155, 641–647. [Google Scholar] [CrossRef]
- Nelles, J.L.; Hu, W.-Y.; Prins, G.S. Estrogen action and prostate cancer. Expert Rev. Endocrinol. Metab. 2011, 6, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Christoforou, P.; Christopoulos, P.F.; Koutsilieris, M. The Role of Estrogen Receptor β in Prostate Cancer. Mol. Med. 2014, 20, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Han, B.; Wu, P.; Li, S.; Lv, Y.; Lu, J.; Yang, Q.; Li, J.; Zhu, Y.; Zhang, Z. Dibutyl phthalate induces allergic airway inflammation in rats via inhibition of the Nrf2/TSLP/JAK1 pathway. Environ. Pollut. 2020, 267, 115564. [Google Scholar] [CrossRef]
- Cartwright, C.D.; Owen, S.A.; Thompson, I.P.; Burns, R.G. Biodegradation of diethyl phthalate in soil by a novel pathway. FEMS Microbiol. Lett. 2000, 186, 27–34. [Google Scholar] [CrossRef]
- Högberg, J.; Hanberg, A.; Berglund, M.; Skerfving, S.; Remberger, M.; Calafat, A.M.; Filipsson, A.F.; Jansson, B.; Johansson, N.; Appelgren, M.; et al. Phthalate Diesters and Their Metabolites in Human Breast Milk, Blood or Serum, and Urine as Biomarkers of Exposure in Vulnerable Populations. Environ. Health Perspect. 2008, 116, 334–339. [Google Scholar] [CrossRef]
- Giovanoulis, G.; Bui, T.; Xu, F.; Papadopoulou, E.; Padilla-Sanchez, J.A.; Covaci, A.; Haug, L.S.; Cousins, A.P.; Magnér, J.; Cousins, I.T.; et al. Multi-pathway human exposure assessment of phthalate esters and DINCH. Environ. Int. 2017, 112, 115–126, Erratum in Environ Int. 2020, 143, 106071. [Google Scholar] [CrossRef]
- Shi, X.-Y.; Wang, Z.; Liu, L.; Feng, L.-M.; Li, N.; Liu, S.; Gao, H. Low concentrations of bisphenol A promote human ovarian cancer cell proliferation and glycolysis-based metabolism through the estrogen receptor-α pathway. Chemosphere 2017, 185, 361–367. [Google Scholar] [CrossRef]
- Mughees, M.; Chugh, H.; Wajid, S. Mechanism of phthalate esters in the progression and development of breast cancer. Drug Chem. Toxicol. 2020, 45, 1021–1025. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.-A.; Li, Q.-L.; Li, Z.-S.; Zheng, P.-J.; Zhang, H.-Z.; Huang, X.-F.; Chi, S.-M.; Yang, A.-G.; Cui, R. Apoptosis drives cancer cells proliferate and metastasize. J. Cell. Mol. Med. 2013, 17, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Wang, Y.; Xue, H.; Romanish, M.T.; Mager, D.L.; Helgason, C.D.; Wang, Y. Genistein versus ICI 182, 780: An ally or enemy in metastatic progression of prostate cancer. Prostate 2013, 73, 1747–1760. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, K.; Habrowska-Górczyńska, D.E.; Urbanek, K.A.; Domińska, K.; Piastowska-Ciesielska, A.W. Estrogen Receptor α Is Crucial in Zearalenone-Induced Invasion and Migration of Prostate Cancer Cells. Toxins 2018, 10, 98. [Google Scholar] [CrossRef]
- Awada, Z.; Nasr, R.; Akika, R.; Cahais, V.; Cuenin, C.; Zhivagui, M.; Herceg, Z.; Ghantous, A.; Zgheib, N.K. DNA methylome-wide alterations associated with estrogen receptor-dependent effects of bisphenols in breast cancer. Clin. Epigenetics 2019, 11, 138. [Google Scholar] [CrossRef]
- Bilancio, A.; Bontempo, P.; Di Donato, M.; Conte, M.; Giovannelli, P.; Altucci, L.; Migliaccio, A.; Castoria, G. Bisphenol A induces cell cycle arrest in primary and prostate cancer cells through EGFR/ERK/p53 signaling pathway activation. Oncotarget 2017, 8, 115620–115631. [Google Scholar] [CrossRef]
- Ferguson, K.K.; Loch-Caruso, R.; Meeker, J.D. Exploration of Oxidative Stress and Inflammatory Markers in Relation to Urinary Phthalate Metabolites: NHANES 1999–2006. Environ. Sci. Technol. 2011, 46, 477–485. [Google Scholar] [CrossRef]
- Sley, E.G.; Rosen, E.M.; Erve, T.J.v.; Sathyanarayana, S.; Barrett, E.S.; Nguyen, R.H.N.; Bush, N.R.; Milne, G.L.; Swan, S.H.; Ferguson, K.K. Omega-3 fatty acid supplement use and oxidative stress levels in pregnancy. PLoS ONE 2020, 15, e0240244. [Google Scholar] [CrossRef]
- Rusyn, I.; Kadiiska, M.B.; Dikalova, A.; Kono, H.; Yin, M.; Tsuchiya, K.; Mason, R.P.; Peters, J.M.; Gonzalez, F.J.; Segal, B.H.; et al. Phthalates Rapidly Increase Production of Reactive Oxygen Species in Vivo: Role of Kupffer Cells. Mol. Pharmacol. 2001, 59, 744–750. [Google Scholar] [CrossRef]
- Tang, X.; Wu, S.; Shen, L.; Wei, Y.; Cao, X.; Wang, Y.; Long, C.; Zhou, Y.; Li, D.; Huang, F.; et al. Di-(2-ethylhexyl) phthalate (DEHP)-induced testicular toxicity through Nrf2-mediated Notch1 signaling pathway in Sprague-Dawley rats. Environ. Toxicol. 2018, 33, 720–728. [Google Scholar] [CrossRef]
- Tetz, L.M.; Cheng, A.A.; Korte, C.S.; Giese, R.W.; Wang, P.; Harris, C.; Meeker, J.D.; Loch-Caruso, R. Mono-2-ethylhexyl phthalate induces oxidative stress responses in human placental cells in vitro. Toxicol. Appl. Pharmacol. 2013, 268, 47–54. [Google Scholar] [CrossRef]
- Hlisníková, H.; Petrovičová, I.; Kolena, B.; Šidlovská, M.; Sirotkin, A. Effects and Mechanisms of Phthalates’ Action on Reproductive Processes and Reproductive Health: A Literature Review. Int. J. Environ. Res. Public Health 2020, 17, 6811. [Google Scholar] [CrossRef]
- Huang, S.; Wa, Q.; Pan, J.; Peng, X.; Ren, D.; Huang, Y.; Chen, X.; Tang, Y. Downregulation of miR-141-3p promotes bone metastasis via activating NF-κB signaling in prostate cancer. J. Exp. Clin. Cancer Res. 2017, 36, 173. [Google Scholar] [CrossRef]
- Mu, J.-W.; Zhou, X.-Y.; Wang, Q.-J.; Han, L.-H.; Jiao, J.-B. MicroRNA-141-3p promoted the progression of nasopharyngeal carcinoma through targeting DLC1. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 11105–11113. [Google Scholar]
- Wu, J.; Shen, Z. Exosomal miRNAs as biomarkers for diagnostic and prognostic in lung cancer. Cancer Med. 2020, 9, 6909–6922. [Google Scholar] [CrossRef] [PubMed]
- Yao, P.-L.; Lin, Y.-C.; Richburg, J.H. Mono-(2-Ethylhexyl) Phthalate (MEHP) Promotes Invasion and Migration of Human Testicular Embryonal Carcinoma Cells1. Biol. Reprod. 2012, 86, 160. [Google Scholar] [CrossRef] [PubMed]
- Forte, M.; Di Lorenzo, M.; Carrizzo, A.; Valiante, S.; Vecchione, C.; Laforgia, V.; De Falco, M. Nonylphenol effects on human prostate non tumorigenic cells. Toxicology 2016, 357–358, 21–32. [Google Scholar] [CrossRef]
- Forte, M.; Di Lorenzo, M.; Iachetta, G.; Mita, D.G.; Laforgia, V.; De Falco, M. Nonylphenol acts on prostate adenocarcinoma cells via estrogen molecular pathways. Ecotoxicol. Environ. Saf. 2019, 180, 412–419. [Google Scholar] [CrossRef]
- Heath, R.L.; Tappel, A.L. A new sensitive assay for the measurement of hydroperoxides. Anal. Biochem. 1976, 76, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Fasciolo, G.; Napolitano, G.; Aprile, M.; Cataldi, S.; Costa, V.; Tomajoli, M.T.M.; Lombardi, A.; Di Meo, S.; Venditti, P. Muscle Oxidative Stress Plays a Role in Hyperthyroidism-Linked Insulin Resistance. Antioxidants 2023, 12, 592. [Google Scholar] [CrossRef]
- Fasciolo, G.; Napolitano, G.; Aprile, M.; Cataldi, S.; Costa, V.; Ciccodicola, A.; Di Meo, S.; Venditti, P. Hepatic Insulin Resistance in Hyperthyroid Rat Liver: Vitamin E Supplementation Highlights a Possible Role of ROS. Antioxidants 2022, 11, 1295. [Google Scholar] [CrossRef] [PubMed]
- Erel, O. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin. Biochem. 2004, 37, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, G.; Capriello, T.; Venditti, P.; Fasciolo, G.; La Pietra, A.; Trifuoggi, M.; Giarra, A.; Agnisola, C.; Ferrandino, I. Aluminum induces a stress response in zebrafish gills by influencing metabolic parameters, morphology, and redox homeostasis. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2023, 271, 109633. [Google Scholar] [CrossRef] [PubMed]
















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mileo, A.; Chianese, T.; Fasciolo, G.; Venditti, P.; Capaldo, A.; Rosati, L.; De Falco, M. Effects of Dibutylphthalate and Steroid Hormone Mixture on Human Prostate Cells. Int. J. Mol. Sci. 2023, 24, 14341. https://doi.org/10.3390/ijms241814341
Mileo A, Chianese T, Fasciolo G, Venditti P, Capaldo A, Rosati L, De Falco M. Effects of Dibutylphthalate and Steroid Hormone Mixture on Human Prostate Cells. International Journal of Molecular Sciences. 2023; 24(18):14341. https://doi.org/10.3390/ijms241814341
Chicago/Turabian StyleMileo, Aldo, Teresa Chianese, Gianluca Fasciolo, Paola Venditti, Anna Capaldo, Luigi Rosati, and Maria De Falco. 2023. "Effects of Dibutylphthalate and Steroid Hormone Mixture on Human Prostate Cells" International Journal of Molecular Sciences 24, no. 18: 14341. https://doi.org/10.3390/ijms241814341