
Fluconazole-Induced Protein Changes in Osteogenic and Immune Metabolic Pathways of Dental Pulp Mesenchymal Stem Cells of Osteopetrosis Patients

Abstract
:1. Introduction
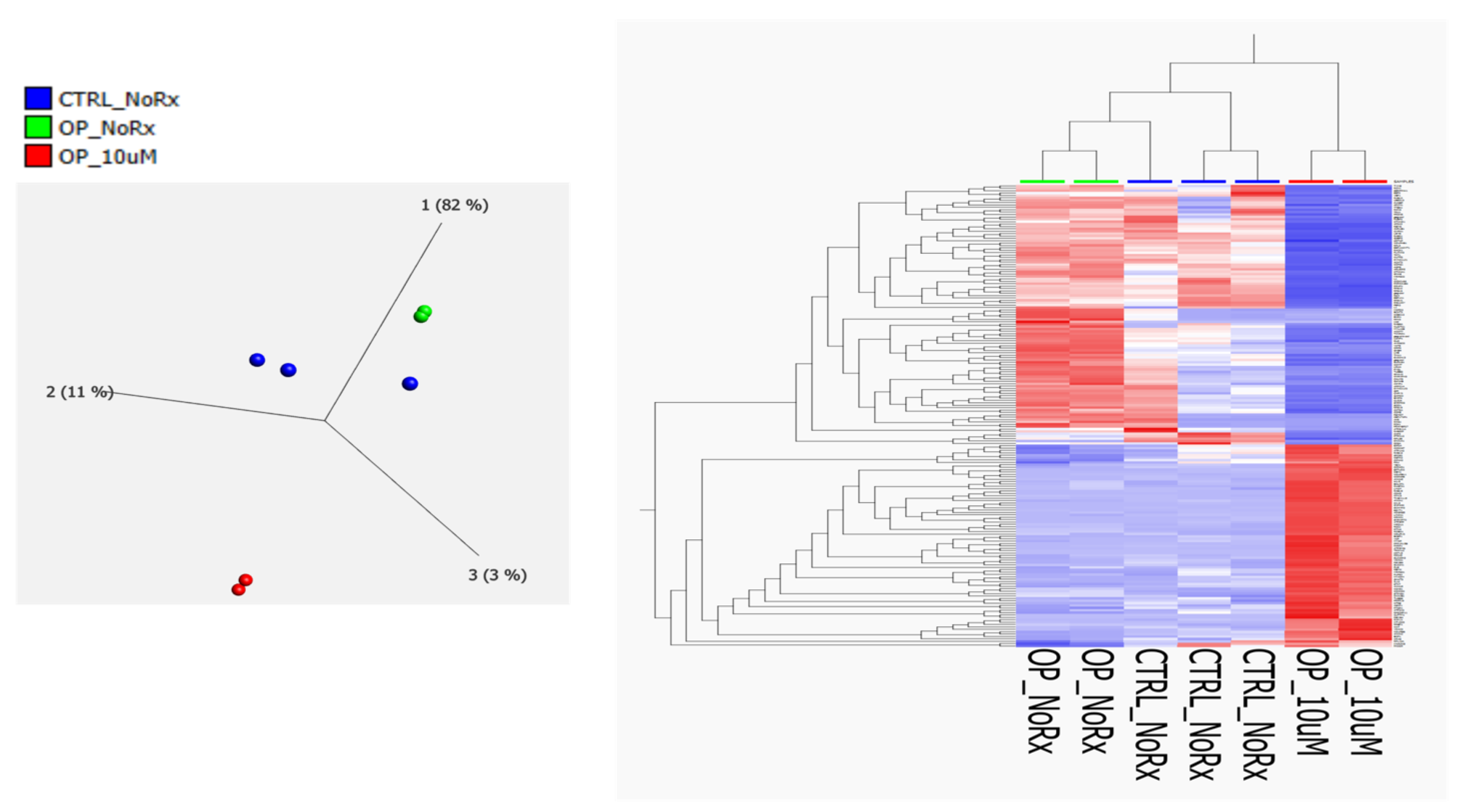
2. Results
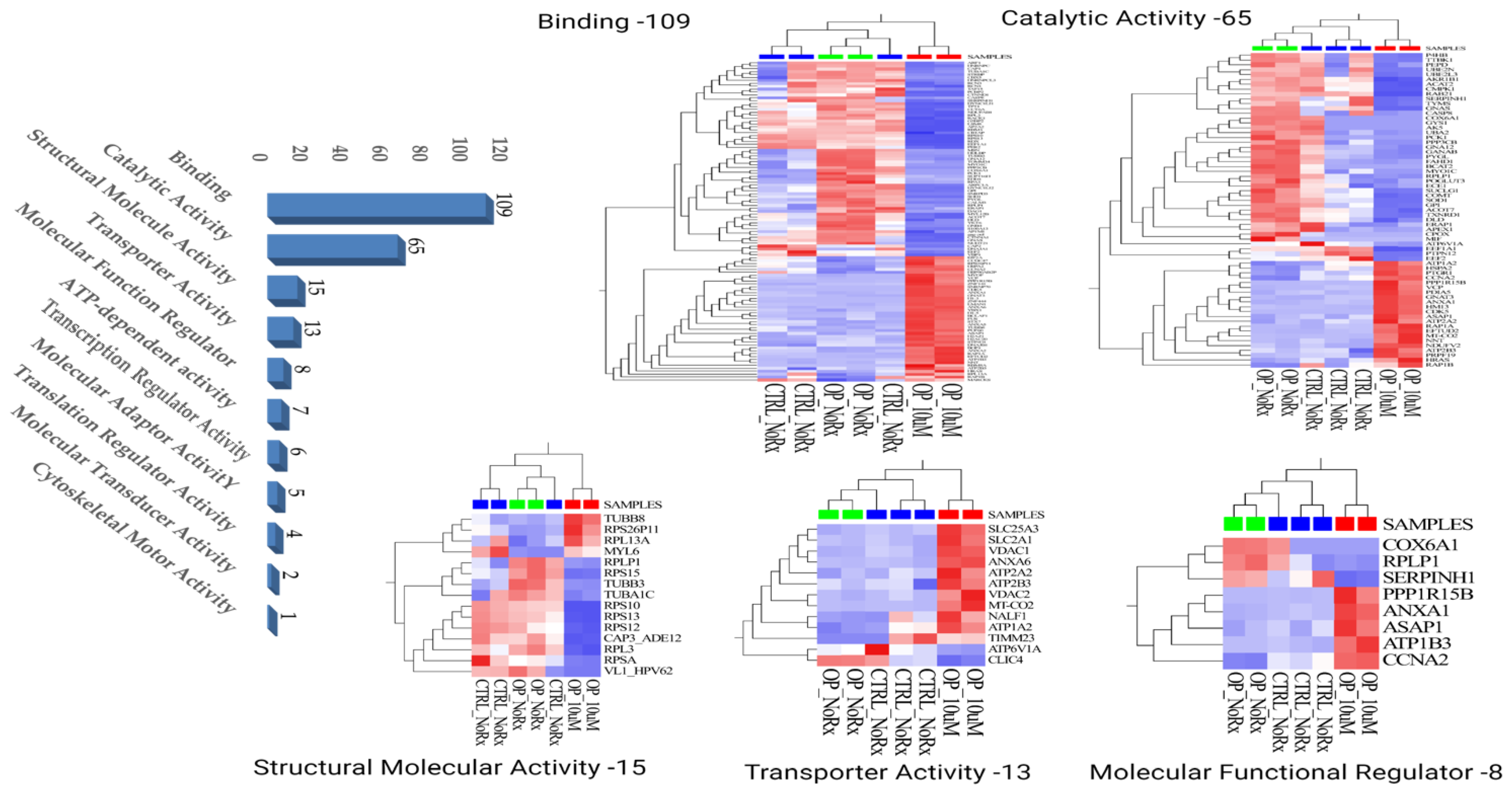
2.1. Molecular Functional Classification of the Treatment Induced Protein Changes
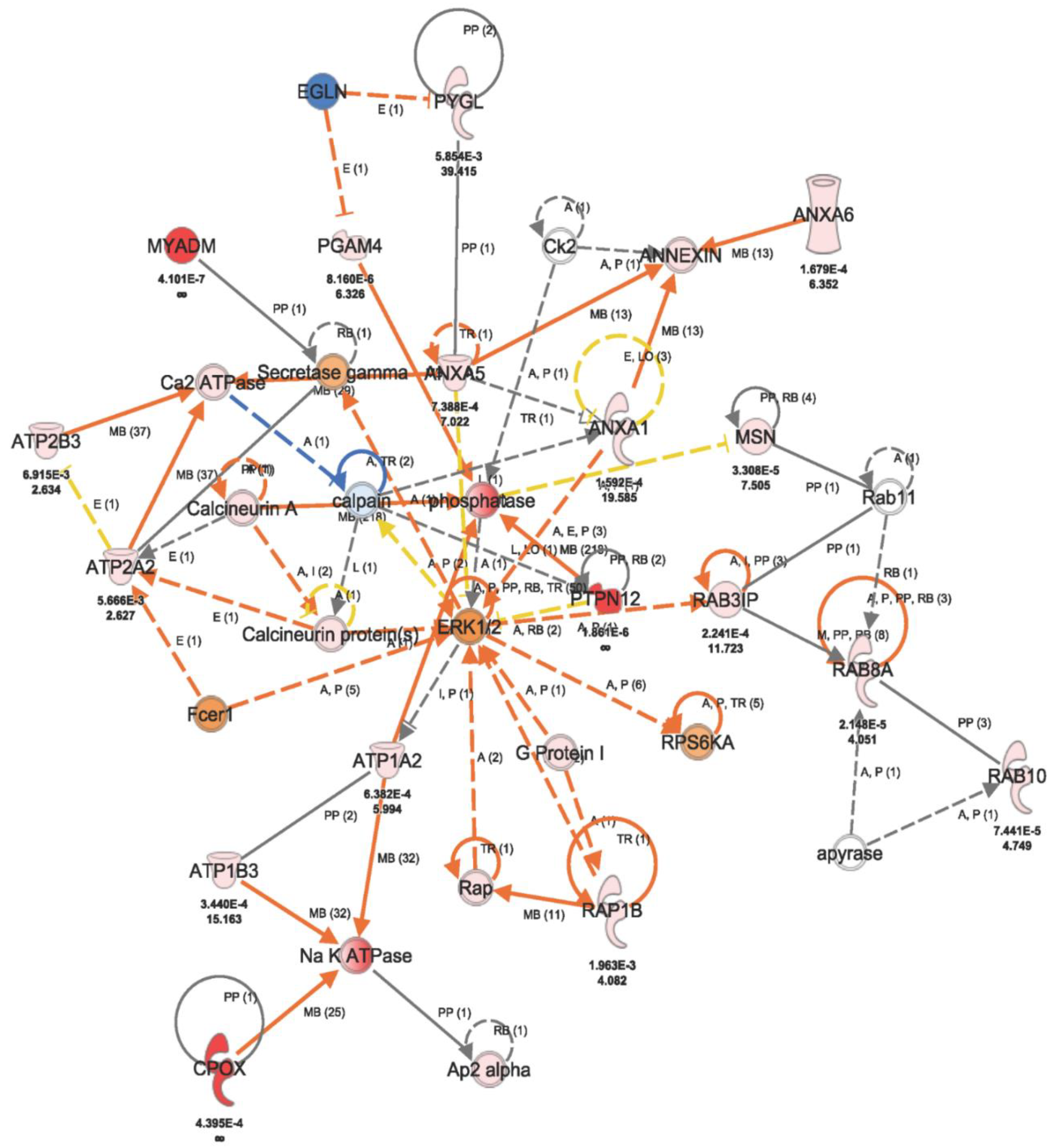
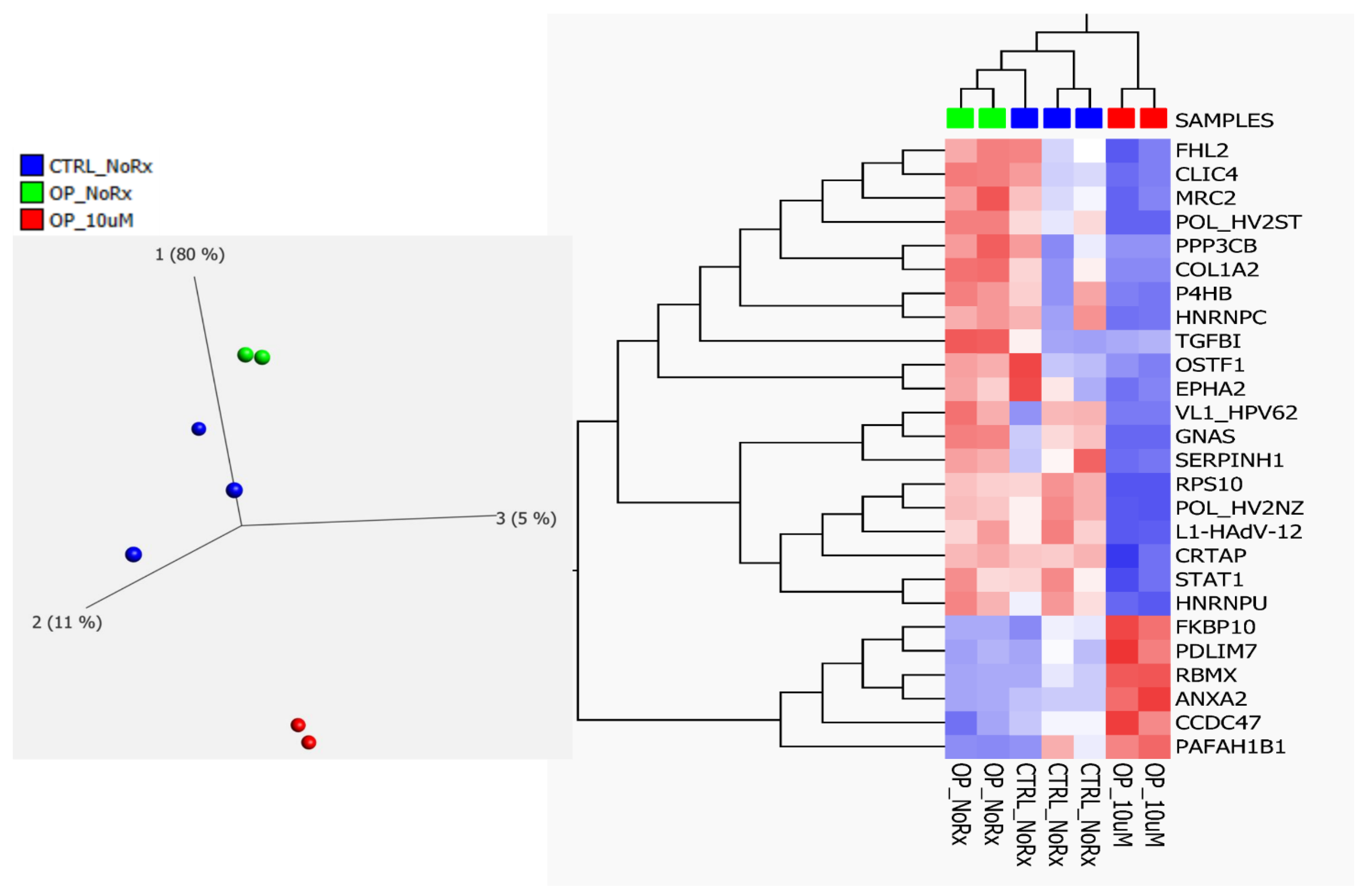
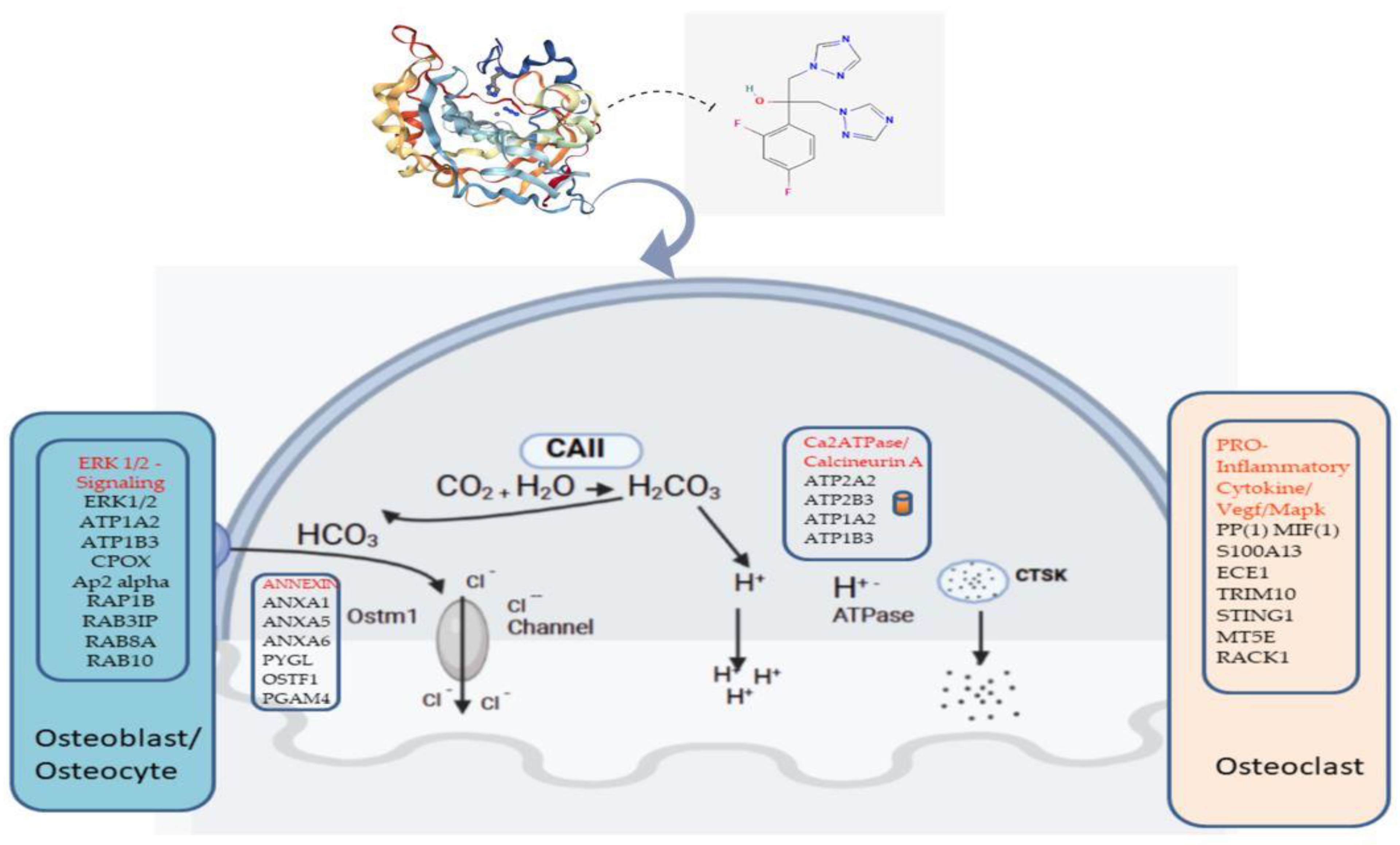
2.2. Fluconazole-Induced Protein Changes Involving Osteogenesis
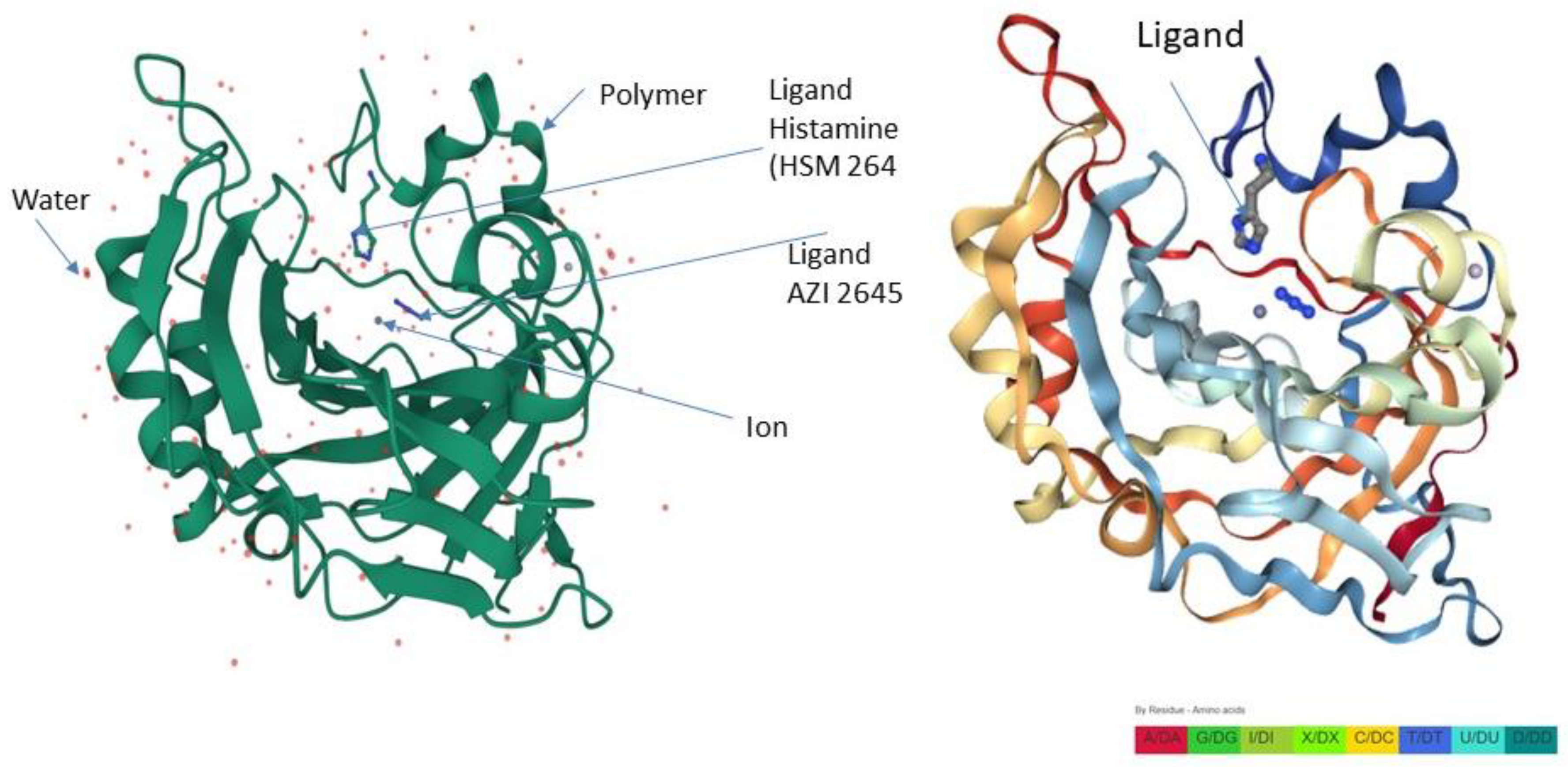
3. Discussion
4. Materials and Methods
4.1. Patient Samples
4.2. Cell Isolation, Culture and Fluconazole Treatment of Dental Pulp Mesenchymal Cells
4.3. Whole-Cell Lysate of Mesenchymal Stromal Cells
4.4. In-Solution Digestion of Complex Protein Mixture
4.5. Protein Identification by Label-Free Liquid Chromatography Mass Spectrometry (LC/MSE)
4.6. Data Analysis and Informatics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SHED | Stem cells from human exfoliated deciduous teeth |
| MSCs | Mesenchymal stem cells |
| LC–MS/MS | Liquid chromatography–tandem mass spectrometry |
| CA-II | Carbonic Anhydrase II |
| CAA | Carbonic Anhydrase Activator |
| HC | Healthy control |
| OP | Osteopetrosis patients |
| DP | Dental pulp |
| MSSCs | Mesenchymal stem/stromal cells |
References
- Ohlsson, A.; Cumming, W.A.; Paul, A.; Sly, W.S. Carbonic anhydrase II deficiency syndrome: Recessive osteopetrosis with renal tubular acidosis and cerebral calcification. Pediatrics 1986, 77, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Ohlsson, A.; Stark, G.; Sakati, N. Marble brain disease: Recessive osteopetrosis, renal tubular acidosis and cerebral calcification in three Saudi Arabian families. Dev. Med. Child Neurol. 1980, 22, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.G. Control of bone resorption by hematopoietic tissue. The induction and reversal of congenital osteopetrosis in mice through use of bone marrow and splenic transplants. J. Exp. Med. 1975, 142, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Udagawa, N.; Takahashi, N.; Akatsu, T.; Tanaka, H.; Sasaki, T.; Nishihara, T.; Koga, T.; Martin, T.J.; Suda, T. Origin of osteoclasts: Mature monocytes and macrophages are capable of differentiating into osteoclasts under a suitable microenvironment prepared by bone marrow-derived stromal cells. Proc. Natl. Acad. Sci. USA 1990, 87, 7260–7264. [Google Scholar] [CrossRef] [PubMed]
- Madel, M.B.; Ibanez, L.; Wakkach, A.; de Vries, T.J.; Teti, A.; Apparailly, F.; Blin-Wakkach, C. Immune Function and Diversity of Osteoclasts in Normal and Pathological Conditions. Front. Immunol. 2019, 10, 1408. [Google Scholar] [CrossRef] [PubMed]
- Cappariello, A.; Maurizi, A.; Veeriah, V.; Teti, A. The Great Beauty of the osteoclast. Arch. Biochem. Biophys. 2014, 558, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Penna, S.; Capo, V.; Palagano, E.; Sobacchi, C.; Villa, A. One Disease, Many Genes: Implications for the Treatment of Osteopetroses. Front. Endocrinol. 2019, 10, 85. [Google Scholar] [CrossRef]
- Alkhayal, Z.; Shinwari, Z.; Gaafar, A.; Alaiya, A. Proteomic Profiling of the First Human Dental Pulp Mesenchymal Stem/Stromal Cells from Carbonic Anhydrase II Deficiency Osteopetrosis Patients. Int. J. Mol. Sci. 2020, 22, 380. [Google Scholar] [CrossRef]
- Wu, C.C.; Econs, M.J.; DiMeglio, L.A.; Insogna, K.L.; Levine, M.A.; Orchard, P.J.; Miller, W.P.; Petryk, A.; Rush, E.T.; Shoback, D.M.; et al. Diagnosis and Management of Osteopetrosis: Consensus Guidelines From the Osteopetrosis Working Group. J. Clin. Endocrinol. Metab. 2017, 102, 3111–3123. [Google Scholar] [CrossRef]
- Stark, Z.; Savarirayan, R. Osteopetrosis. Orphanet J. Rare Dis. 2009, 4, 5. [Google Scholar] [CrossRef]
- Coudert, A.E.; de Vernejoul, M.C.; Muraca, M.; Del Fattore, A. Osteopetrosis and its relevance for the discovery of new functions associated with the skeleton. Int. J. Endocrinol. 2015, 2015, 372156. [Google Scholar] [CrossRef] [PubMed]
- Villa, A.; Guerrini, M.M.; Cassani, B.; Pangrazio, A.; Sobacchi, C. Infantile malignant, autosomal recessive osteopetrosis: The rich and the poor. Calcif. Tissue Int. 2009, 84, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, Y.; Zhang, B.; Zhou, J.; Li, T.; Liu, Z.; Li, Y.; Yang, M. Molecular insights into the human CLC-7/Ostm1 transporter. Sci. Adv. 2020, 6, eabb4747. [Google Scholar] [CrossRef] [PubMed]
- Landa, J.; Margolis, N.; Di Cesare, P. Orthopaedic management of the patient with osteopetrosis. J. Am. Acad. Orthop. Surg. 2007, 15, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Askmyr, M.K.; Fasth, A.; Richter, J. Towards a better understanding and new therapeutics of osteopetrosis. Br. J. Haematol. 2008, 140, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Kato, H.; Nonaka, K.; Nakanishi, H. Stem cells from human exfoliated deciduous teeth attenuate mechanical allodynia in mice through distinct from the siglec-9/MCP-1-mediated tissue-repairing mechanism. Sci. Rep. 2021, 11, 20053. [Google Scholar] [CrossRef] [PubMed]
- Miura, M.; Gronthos, S.; Zhao, M.; Lu, B.; Fisher, L.W.; Robey, P.G.; Shi, S. SHED: Stem cells from human exfoliated deciduous teeth. Proc. Natl. Acad. Sci. USA 2003, 100, 5807–5812. [Google Scholar] [CrossRef] [PubMed]
- Nagai, R.; Kooh, S.W.; Balfe, J.W.; Fenton, T.; Halperin, M.L. Renal tubular acidosis and osteopetrosis with carbonic anhydrase II deficiency: Pathogenesis of impaired acidification. Pediatr. Nephrol. 1997, 11, 633–636. [Google Scholar] [CrossRef]
- Akocak, S.; Lolak, N.; Bua, S.; Nocentini, A.; Supuran, C.T. Activation of human α-carbonic anhydrase isoforms I, II, IV and VII with bis-histamine schiff bases and bis-spinaceamine substituted derivatives. J. Enzym. Inhib. Med. Chem. 2019, 34, 1193–1198. [Google Scholar] [CrossRef]
- Akocak, S.; Supuran, C.T. Activation of α-, β-, γ- δ-, ζ- and η- class of carbonic anhydrases with amines and amino acids: A review. J. Enzym. Inhib. Med. Chem. 2019, 34, 1652–1659. [Google Scholar] [CrossRef]
- Lolak, N.; Akocak, S.; Bua, S.; Koca, M.; Supuran, C.T. Design and synthesis of novel 1,3-diaryltriazene-substituted sulfonamides as potent and selective carbonic anhydrase II inhibitors. Bioorgan. Chem. 2018, 77, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Lolak, N.; Akocak, S.; Bua, S.; Supuran, C.T. Design, synthesis and biological evaluation of novel ureido benzenesulfonamides incorporating 1,3,5-triazine moieties as potent carbonic anhydrase IX inhibitors. Bioorgan. Chem. 2019, 82, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, X.; Lai, J.; Xu, W.; Zhu, W.; Chen, G. Protein tyrosine phosphatase non-receptor type 12 suppresses tumor progression in osteosarcoma cells. J. Orthop. Sci. 2023, 28, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.M.; Schwartz, G.J.; Alper, S.L. Urinary Acidification. In Fetal and Neonatal Physiology, 4th ed.; Polin, R.A., Abman, S.H., Rowitch, D.H., Benitz, W.E., Fox, W.W., Eds.; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Blandina, P.; Supuran, C.T. Carbonic anhydrase activators and their potential in the pharmaceutical field. In Carbonic Anhydrases; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar] [CrossRef]
- Coban, T.A.; Beydemir, Ş.; Gücin, İ.; Ekinci, D.; Innocenti, A.; Vullo, D.; Supuran, C.T. Sildenafil is a strong activator of mammalian carbonic anhydrase isoforms I–XIV. Bioorgan. Med. Chem. 2009, 17, 5791–5795. [Google Scholar] [CrossRef] [PubMed]
- Akocak, S.; Lolak, N.; Bua, S.; Nocentini, A.; Karakoc, G.; Supuran, C.T. α-Carbonic anhydrases are strongly activated by spinaceamine derivatives. Bioorgan. Med. Chem. 2019, 27, 800–804. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, A.; Ikeda, H.; Tsukamoto, H.; Kihira, K.; Ishioka, M.; Hirose, J.; Hata, T.; Fujioka, H.; Ono, Y. Timolol activates the enzyme activities of human carbonic anhydrase I and II. Biol. Pharm. Bull. 2010, 33, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Almstedt, K.; Rafstedt, T.; Supuran, C.T.; Carlsson, U.; Hammarstrom, P. Small-molecule suppression of misfolding of mutated human carbonic anhydrase II linked to marble brain disease. Biochemistry 2009, 48, 5358–5364. [Google Scholar] [CrossRef]
- Datta, R.; Shah, G.N.; Rubbelke, T.S.; Waheed, A.; Rauchman, M.; Goodman, A.G.; Katze, M.G.; Sly, W.S. Progressive renal injury from transgenic expression of human carbonic anhydrase IV folding mutants is enhanced by deficiency of p58IPK. Proc. Natl. Acad. Sci. USA 2010, 107, 6448–6452. [Google Scholar] [CrossRef]
- Feldshtein, M.; Elkrinawi, S.; Yerushalmi, B.; Marcus, B.; Vullo, D.; Romi, H.; Ofir, R.; Landau, D.; Sivan, S.; Supuran, C.T.; et al. Hyperchlorhidrosis caused by homozygous mutation in CA12, encoding carbonic anhydrase XII. Am. J. Hum. Genet. 2010, 87, 713–720. [Google Scholar] [CrossRef]
- Van Karnebeek, C.D.; Sly, W.S.; Ross, C.J.; Salvarinova, R.; Yaplito-Lee, J.; Santra, S.; Shyr, C.; Horvath, G.A.; Eydoux, P.; Lehman, A.M.; et al. Mitochondrial carbonic anhydrase VA deficiency resulting from CA5A alterations presents with hyperammonemia in early childhood. Am. J. Hum. Genet. 2014, 94, 453–461. [Google Scholar] [CrossRef]
- Van Wesenbeeck, L.; Odgren, P.R.; Coxon, F.P.; Frattini, A.; Moens, P.; Perdu, B.; MacKay, C.A.; Van Hul, E.; Timmermans, J.P.; Vanhoenacker, F.; et al. Involvement of PLEKHM1 in osteoclastic vesicular transport and osteopetrosis in incisors absent rats and humans. J. Clin. Investig. 2007, 117, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Ogilvie, J.M.; Ohlemiller, K.K.; Shah, G.N.; Ulmasov, B.; Becker, T.A.; Waheed, A.; Hennig, A.K.; Lukasiewicz, P.D.; Sly, W.S. Carbonic anhydrase XIV deficiency produces a functional defect in the retinal light response. Proc. Natl. Acad. Sci. USA 2007, 104, 8514–8519. [Google Scholar] [CrossRef] [PubMed]
- Muller, W.E.; Schroder, H.C.; Schlossmacher, U.; Grebenjuk, V.A.; Ushijima, H.; Wang, X. Induction of carbonic anhydrase in SaOS-2 cells, exposed to bicarbonate and consequences for calcium phosphate crystal formation. Biomaterials 2013, 34, 8671–8680. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Schroder, H.C.; Muller, W.E. Biocalcite, a multifunctional inorganic polymer: Building block for calcareous sponge spicules and bioseed for the synthesis of calcium phosphate-based bone. Beilstein J. Nanotechnol. 2014, 5, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Margolis, D.S.; Szivek, J.A.; Lai, L.-W.; Lien, Y.-H.H. Phenotypic characteristics of bone in carbonic anhydrase II-deficient mice. Calcif. Tissue Int. 2008, 82, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Zheng, Y.; Yang, Q.; Wang, L.; Pan, J.; Xia, Y.; Yan, X.; Han, J. Carbonic anhydrase I (CA1) is involved in the process of bone formation and is susceptible to ankylosing spondylitis. Arthritis Res. Ther. 2012, 14, R176. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Guitron, L.E.; Ceron-Torres, T.; Sobacchi, C.; Ochoa-Ruiz, E.; Villegas-Huesca, S. Autosomal recessive osteopetrosis type I: Description of pathogenic variant of TCIRG1 gene. Bol. Med. Hosp. Infant. Mex. 2018, 75, 255–259. [Google Scholar] [CrossRef]
- Athanasiadou, E.; Vlachou, C.; Theocharidou, A.; Tilaveridis, I.; Vargiami, E.; Antoniadis, K.; Arapostathis, K. When a pedodontic examination leads to the diagnosis of osteopetrosis: A case report. Spec. Care Dent. 2020, 40, 113–120. [Google Scholar] [CrossRef]
- Debruyne, D.; Ryckelynck, J.P. Clinical pharmacokinetics of fluconazole. Clin. Pharmacokinet. 1993, 24, 10–27. [Google Scholar] [CrossRef]
- Rose, A.S.; Bradley, A.R.; Valasatava, Y.; Duarte, J.M.; Prlić, A.; Rose, P.W. NGL viewer: Web-based molecular graphics for large complexes. Bioinformatics 2018, 34, 3755–3758. [Google Scholar] [CrossRef]
- Kim, J.M.; Yang, Y.S.; Hong, J.; Chaugule, S.; Chun, H.; van der Meulen, M.C.H.; Xu, R.; Greenblatt, M.B.; Shim, J.H. Biphasic regulation of osteoblast development via the ERK MAPK–mTOR pathway. Elife 2022, 11, e78069. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Xu, Y.; Gan, Y.; Song, L.; Zhang, C.; Wang, L.; Zhou, Q. Role of the ERK1/2 Signaling Pathway in Osteogenesis of Rat Tendon-Derived Stem Cells in Normoxic and Hypoxic Cultures. Int. J. Med. Sci. 2016, 13, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Rousselle, A.V.; Heymann, D. Osteoclastic acidification pathways during bone resorption. Bone 2002, 30, 533–540. [Google Scholar] [CrossRef]
- Schlesinger, P.H.; Mattsson, J.P.; Blair, H.C. Osteoclastic acid transport: Mechanism and implications for physiological and pharmacological regulation. Miner. Electrolyte Metab. 1994, 20, 31–39. [Google Scholar] [PubMed]
- Schneider, M.R.; Sibilia, M.; Erben, R.G. The EGFR network in bone biology and pathology. Trends Endocrinol. Metab. TEM 2009, 20, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Alkhayal, Z.; Shinwari, Z.; Gaafar, A.; Alaiya, A. Carbonic Anhydrase II Activators in Osteopetrosis Treatment: A Review. Curr. Issues Mol. Biol. 2023, 45, 1373–1386. [Google Scholar] [CrossRef] [PubMed]
- Even-Or, E.; NaserEddin, A.; Dinur Schejter, Y.; Shadur, B.; Zaidman, I.; Stepensky, P. Haploidentical stem cell transplantation with post-transplant cyclophosphamide for osteopetrosis and other nonmalignant diseases. Bone Marrow Transplant. 2021, 56, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Handler, D.C.; Pascovici, D.; Mirzaei, M.; Gupta, V.; Salekdeh, G.H.; Haynes, P.A. The Art of Validating Quantitative Proteomics Data. Proteomics 2018, 18, e1800222. [Google Scholar] [CrossRef]
- Franzen, B.; Hirano, T.; Okuzawa, K.; Uryu, K.; Alaiya, A.A.; Linder, S.; Auer, G. Sample preparation of human tumors prior to two-dimensional electrophoresis of proteins. Electrophoresis 1995, 16, 1087–1089. [Google Scholar] [CrossRef]
- Franzen, B.; Linder, S.; Okuzawa, K.; Kato, H.; Auer, G. Nonenzymatic extraction of cells from clinical tumor material for analysis of gene expression by two-dimensional polyacrylamide gel electrophoresis. Electrophoresis 1993, 14, 1045–1053. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Alaiya, A.A.; Aljurf, M.; Shinwari, Z.; Almohareb, F.; Malhan, H.; Alzahrani, H.; Owaidah, T.; Fox, J.; Alsharif, F.; Mohamed, S.Y.; et al. Protein signatures as potential surrogate biomarkers for stratification and prediction of treatment response in chronic myeloid leukemia patients. Int. J. Oncol. 2016, 49, 913–933. [Google Scholar] [CrossRef]
- AlZahrani, S.; Shinwari, Z.; Gaafar, A.; Alaiya, A.; Al-Kahtani, A. Anti-Inflammatory Effect of Specialized Proresolving Lipid Mediators on Mesenchymal Stem Cells: An In Vitro Study. Cells 2022, 12, 122. [Google Scholar] [CrossRef]
- Alaiya, A.; Alshukairi, A.; Shinwari, Z.; Al-Fares, M.; Alotaibi, J.; AlOmaim, W.; Alsharif, I.; Bakheet, R.; Alharbi, L.; Allam, R.; et al. Alterations in the Plasma Proteome Induced by SARS-CoV-2 and MERS-CoV Reveal Biomarkers for Disease Outcomes for COVID-19 Patients. J. Inflamm. Res. 2021, 14, 4313–4328. [Google Scholar] [CrossRef]
- Dasouki, M.; Alaiya, A.; ElAmin, T.; Shinwari, Z.; Monies, D.; Abouelhoda, M.; Jabaan, A.; Almourfi, F.; Rahbeeni, Z.; Alsohaibani, F.; et al. Comprehensive multi-omics analysis of G6PC3 deficiency-related congenital neutropenia with inflammatory bowel disease. iScience 2021, 24, 102214. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subjects | Age (Years) | Gender | Pathogenetics | Skeletal Radiology | Clinical Findings |
|---|---|---|---|---|---|
| OP#1 | 11 | M | c.232 + 1G > A, IVS2 + 1G > Aof the CA2 gene, (AR, RTA) | High frequency of fractures, dense bones | Dental abnormalities, developmental delay, optic atrophy/esotropia, ADHD |
| OP#2 | 10 | F | c.232 + 1G > A, IVS2 + 1G > Aof the CA2 gene (AR, RTA) | Dense skull bones | Short stature, esotropia and optic atrophy |
| OP#3 | 16 | F | CAII deficiency Renal Profile Low CO2, (RTA) | Diffuse increased density of skeletal bones, sclerosis more prominent in skull | ADHD, developmental delay learning disabilities, scoliosis |
| OP#4 | 15 | M | CAII deficiency Renal profile Low CO2, (RTA) | Brachiocephaly, increased bone density, Erlenmeyer flask deformity | Failure to thrive, fractures, intracranial calcification |
| OP#5 | 9 | M | c.232 + 1G > A;IVS2 + 1G > A (RTA), AD, CAII | Generalized increase in bone density shape normal | Calcification, fractures |
| HC#1 | 11 | F | Healthy | Not indicated | No Abnormalities |
| HC#2 | 9 | F | Healthy | Not indicated | No Abnormalities |
| HC#3 | 10 | M | Healthy | Not indicated | No Abnormalities |
| HC#4 | 16 | M | Healthy | Not indicated | No Abnormalities |
| HC#5 | 9 | M | Healthy | Not indicated | No Abnormalities |
| HC#6 | 13 | F | Healthy | Not indicated | No Abnormalities |
| Accession | GN | Anova (p) | Max Fold Change | Highest Mean Condition | Lowest Mean Condition | Role in Bone Related Disease Association |
|---|---|---|---|---|---|---|
| O75718 | CRTAP | 0.0193 | 3.30 | OP-NoRx | OP-Rx | Ostegenesis Imperfecta |
| P05962 | POL_HV2NZ | 0.0001 | 81.79 | OP-NoRx | OP-Rx | Not known |
| P07237 | P4HB | 0.0002 | 3.32 | OP-NoRx | OP-Rx | Ostegenesis Imperfecta |
| P07910 | HNRNPC | 0.0008 | 3.48 | OP-NoRx | OP-Rx | Osteoblast |
| P08123 | COL1A2 | 0.0305 | 170.43 | OP-NoRx | OP-Rx | Ostegenesis Imperfecta |
| P16298 | PPP3CB | 0.0010 | 42.88 | OP-NoRx | OP-Rx | Osteoclast |
| P20876 | POL_HV2ST | 0.0000 | 22.26 | OP-NoRx | OP-Rx | Not known |
| P29317 | EPHA2 | 0.0444 | 7.03 | OP-NoRx | OP-Rx | Osteoblast-Clast |
| P36712 | L1-HAdV-12 | 0.0004 | 5.12 | OP-NoRx | OP-Rx | Not known |
| P42224 | STAT1 | 0.0461 | 2.29 | OP-NoRx | OP-Rx | Osteoclast |
| P46783 | RPS10 | 0.0041 | 1850.80 | OP-NoRx | OP-Rx | Osteoblast-Clast |
| P50454 | SERPINH1 | 0.0000 | 5.35 | OP-NoRx | OP-Rx | Oste Imperfecta |
| P50823 | VL1_HPV62 | 0.0000 | ∞ | OP-NoRx | OP-Rx | Not known |
| Q5JWF2 | GNAS | 0.0015 | 42.22 | OP-NoRx | OP-Rx | Osteoblast-Clast |
| Q9UBG0 | MRC2 | 0.0151 | 8.64 | OP-NoRx | OP-Rx | Osteoblast |
| Q9Y696 | CLIC4 | 0.0007 | 9.14 | OP-NoRx | OP-Rx | Osteoblast |
| Q00839 | HNRNPU | 0.0117 | 2.46 | OP-NoRx | OP-Rx | Osteoblast |
| Q14192 | FHL2 | 0.0834 | 8.20 | OP-NoRx | OP-Rx | Osteoblast |
| Q92882 | OSTF1 | 0.0109 | 4.56 | OP-NoRx | OP-Rx | Osteoblast |
| Q15582 | TGFBI | 0.0009 | 4.02 | OP-NoRx | OP-Rx | Osteoblast |
| P07355 | ANXA2 | 0.0000 | 6.68 | OP-Rx | OP-NoRx | Osteoblast |
| Q9NR12 | PDLIM7 | 0.0004 | 4.86 | OP-Rx | OP-NoRx | Osteoblast |
| Q96A33 | CCDC47 | 0.0014 | 3.43 | OP-Rx | OP-NoRx | Osteoblast |
| Q96AY3 | FKBP10 | 0.0030 | 5.92 | OP-Rx | OP-NoRx | Oste Imperfata |
| P38159 | RBMX | 0.0020 | 979.35 | OP-Rx | OP-NoRx | Osteoblast |
| P43034 | PAFAH1B1 | 0.0085 | 39.22 | OP-Rx | OP-NoRx | Osteoclast |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alkhayal, Z.; Shinwari, Z.; Gaafar, A.; Alaiya, A. Fluconazole-Induced Protein Changes in Osteogenic and Immune Metabolic Pathways of Dental Pulp Mesenchymal Stem Cells of Osteopetrosis Patients. Int. J. Mol. Sci. 2023, 24, 13841. https://doi.org/10.3390/ijms241813841
Alkhayal Z, Shinwari Z, Gaafar A, Alaiya A. Fluconazole-Induced Protein Changes in Osteogenic and Immune Metabolic Pathways of Dental Pulp Mesenchymal Stem Cells of Osteopetrosis Patients. International Journal of Molecular Sciences. 2023; 24(18):13841. https://doi.org/10.3390/ijms241813841
Chicago/Turabian StyleAlkhayal, Zikra, Zakia Shinwari, Ameera Gaafar, and Ayodele Alaiya. 2023. "Fluconazole-Induced Protein Changes in Osteogenic and Immune Metabolic Pathways of Dental Pulp Mesenchymal Stem Cells of Osteopetrosis Patients" International Journal of Molecular Sciences 24, no. 18: 13841. https://doi.org/10.3390/ijms241813841