
Citri Reticulatae Pericarpium Limits TLR-4-Triggered Inflammatory Response in Raw264.7 Macrophages by Activating RasGRP3

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
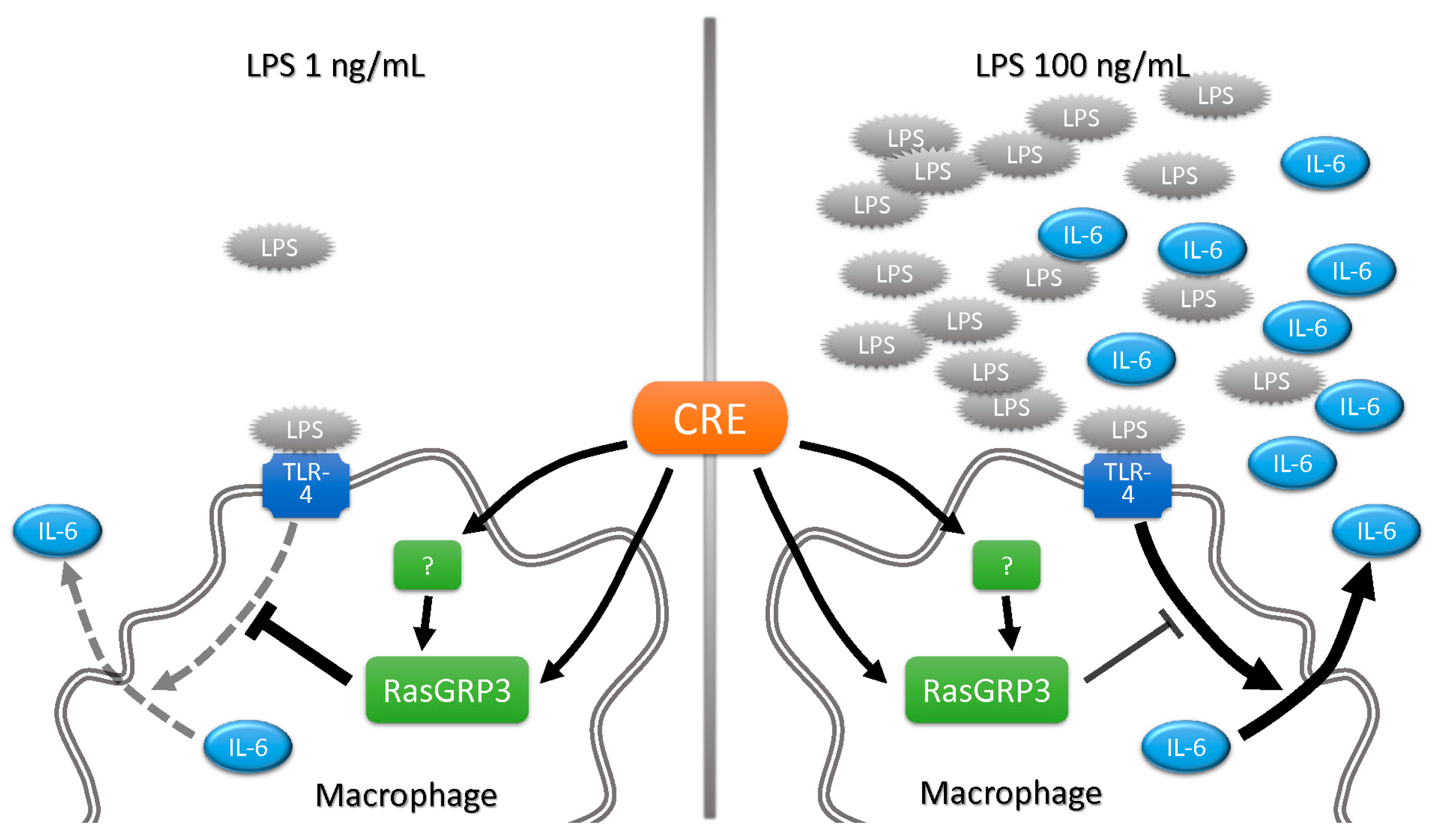
Abstract
:1. Introduction
2. Results
2.1. Effects of CRE on the IL-6 Production in Raw264.7 Cells
2.2. Effects of CRE on the RasGRP3 Expression in Raw264.7 Cells
2.3. Effects of CRE on the IL-6 Production in Raw264.7 Cells Transfected with RasGRP3 siRNA
2.4. Effects of CRE on the RasGRP3 Expression in Raw264.7 Cells Transfected with RasGRP3 siRNA
2.5. Effects of Nobiletin on the IL-6 Production in Raw264.7 Macrophages
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Preparation of Citri Reticulatae Pericarpium Extracts (CRE)
4.3. Cell Preparation and siRNA Transfection
4.4. IL-6 Production in Raw264.7 Cells
4.5. Western Blotting
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hotamisligil, G.S. Inflammation and Metabolic Disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Chen, T.; Yu, Z.; Zhu, X.; Yang, M.; Xie, B.; Li, N.; Cao, X.; Wang, J. RasGRP3 Limits Toll-like Receptor-Triggered Inflammatory Response in Macrophages by Activating Rap1 Small GTPase. Nat. Commun. 2014, 5, 4657. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N. Chapter 61—Nutrients and Gene Expression in Inflammation. In Principles of Nutrigenetics and Nutrigenomics; Caterina, R.D., Martinez, J.A., Kohlmeier, M., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 457–467. ISBN 978-0-12-804572-5. [Google Scholar]
- Li, Z.; Li, L.; Zhao, S.; Li, J.; Zhou, H.; Zhang, Y.; Yang, Z.; Yuan, B. Re-Understanding Anti-Influenza Strategy: Attach Equal Importance to Antiviral and Anti-Inflammatory Therapies. J. Thorac. Dis. 2018, 10, S2248–S2259. [Google Scholar] [CrossRef]
- Coperchini, F.; Chiovato, L.; Croce, L.; Magri, F.; Rotondi, M. The Cytokine Storm in COVID-19: An Overview of the Involvement of the Chemokine/Chemokine-Receptor System. Cytokine Growth Factor Rev. 2020, 53, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.-A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage Plasticity, Polarization, and Function in Health and Disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, N.; Kobayashi, K. Macrophages in Inflammation. Curr. Drug Targets-Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.; McCarthy, J.; O’Driscoll, C.; Melgar, S. Pattern Recognition Receptors—Molecular Orchestrators of Inflammation in Inflammatory Bowel Disease. Cytokine Growth Factor Rev. 2013, 24, 91–104. [Google Scholar] [CrossRef]
- Ali, A.M.; Gabbar, M.A.; Abdel-Twab, S.M.; Fahmy, E.M.; Ebaid, H.; Alhazza, I.M.; Ahmed, O.M. Antidiabetic Potency, Antioxidant Effects, and Mode of Actions of Citrus Reticulata Fruit Peel Hydroethanolic Extract, Hesperidin, and Quercetin in Nicotinamide/Streptozotocin-Induced Wistar Diabetic Rats. Oxid. Med. Cell. Longev. 2020, 2020, e1730492. [Google Scholar] [CrossRef]
- Guo, X.; Cao, X.; Fang, X.; Guo, A.; Li, E. Inhibitory Effects of Fermented Ougan (Citrus reticulata Cv. Suavissima) Juice on High-Fat Diet-Induced Obesity Associated with White Adipose Tissue Browning and Gut Microbiota Modulation in Mice. Food Funct. 2021, 12, 9300–9314. [Google Scholar] [CrossRef]
- Castro, M.A.; Rodenak-Kladniew, B.; Massone, A.; Polo, M.; de Bravo, M.G.; Crespo, R. Citrus Reticulata Peel Oil Inhibits Non-Small Cell Lung Cancer Cell Proliferation in Culture and Implanted in Nude Mice. Food Funct. 2018, 9, 2290–2299. [Google Scholar] [CrossRef]
- Jung, K.H.; Ha, E.; Kim, M.J.; Won, H.-J.; Zheng, L.T.; Kim, H.K.; Hong, S.J.; Chung, J.H.; Yim, S.-V. Suppressive Effects of Nitric Oxide (NO) Production and Inducible Nitric Oxide Synthase (INOS) Expression by Citrus Reticulata Extract in RAW 264.7 Macrophage Cells. Food Chem. Toxicol. 2007, 45, 1545–1550. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, H.; Guo, L.; Zhao, H.; Ho, C.-T. Chemistry and Bioactivity of Nobiletin and Its Metabolites. J. Funct. Foods 2014, 6, 2–10. [Google Scholar] [CrossRef]
- Nakajima, A.; Nemoto, K.; Ohizumi, Y. An Evaluation of the Genotoxicity and Subchronic Toxicity of the Peel Extract of Ponkan Cultivar ‘Ohta Ponkan’ (Citrus reticulata Blanco) That Is Rich in Nobiletin and Tangeretin with Anti-Dementia Activity. Regul. Toxicol. Pharmacol. 2020, 114, 104670. [Google Scholar] [CrossRef] [PubMed]
- Lannoy, V.; Côté-Biron, A.; Asselin, C.; Rivard, N. TIRAP, TRAM, and Toll-Like Receptors: The Untold Story. Mediat. Inflamm. 2023, 2023, e2899271. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Kawai, T.; Akira, S. Dissecting Negative Regulation of Toll-like Receptor Signaling. Trends Immunol. 2012, 33, 449–458. [Google Scholar] [CrossRef]
- Shin, M.-S.; Park, S.B.; Shin, K.-S. Molecular Mechanisms of Immunomodulatory Activity by Polysaccharide Isolated from the Peels of Citrus Unshiu. Int. J. Biol. Macromol. 2018, 112, 576–583. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, W.; Zheng, F.; Yu, H.; Wei, K. Xanthatin Alleviates LPS-Induced Inflammatory Response in RAW264.7 Macrophages by Inhibiting NF-ΚB, MAPK and STATs Activation. Molecules 2022, 27, 4603. [Google Scholar] [CrossRef]
- Koh, T.J.; DiPietro, L.A. Inflammation and Wound Healing: The Role of the Macrophage. Expert Rev. Mol. Med. 2011, 13, e23. [Google Scholar] [CrossRef]
- Liu, B.; Li, M.; Zhou, Z.; Guan, X.; Xiang, Y. Can We Use Interleukin-6 (IL-6) Blockade for Coronavirus Disease 2019 (COVID-19)-Induced Cytokine Release Syndrome (CRS)? J. Autoimmun. 2020, 111, 102452. [Google Scholar] [CrossRef]
- Long, T.; Lv, X.; Xu, Y.; Yang, G.; Xu, L.-Y.; Li, S. Supercritical Fluid CO2 Extraction of Three Polymethoxyflavones from Citri Reticulatae Pericarpium and Subsequent Preparative Separation by Continuous High-Speed Counter-Current Chromatography. J. Chromatogr. B 2019, 1124, 284–289. [Google Scholar] [CrossRef]
- Zhang, H.; Tian, G.; Zhao, C.; Han, Y.; DiMarco-Crook, C.; Lu, C.; Bao, Y.; Li, C.; Xiao, H.; Zheng, J. Characterization of Polymethoxyflavone Demethylation during Drying Processes of Citrus Peels. Food Funct. 2019, 10, 5707–5717. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Guo, Y.; Qiao, Y.; Zhang, J.; Jiang, P. Nobiletin Ameliorates NLRP3 Inflammasome-Mediated Inflammation through Promoting Autophagy via the AMPK Pathway. Mol. Neurobiol. 2020, 57, 5056–5068. [Google Scholar] [CrossRef] [PubMed]
- Ozkan, A.D.; Kaleli, S.; Onen, H.I.; Sarihan, M.; Eskiler, G.G.; Yigin, A.K.; Akdogan, M. Anti-Inflammatory Effects of Nobiletin on TLR4/TRIF/IRF3 and TLR9/IRF7 Signaling Pathways in Prostate Cancer Cells. Immunopharmacol. Immunotoxicol. 2020, 42, 93–100. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.H.; Kim, Y.-S.; Leem, K.H. Citri Reticulatae Pericarpium Limits TLR-4-Triggered Inflammatory Response in Raw264.7 Macrophages by Activating RasGRP3. Int. J. Mol. Sci. 2023, 24, 13777. https://doi.org/10.3390/ijms241813777
Lee JH, Kim Y-S, Leem KH. Citri Reticulatae Pericarpium Limits TLR-4-Triggered Inflammatory Response in Raw264.7 Macrophages by Activating RasGRP3. International Journal of Molecular Sciences. 2023; 24(18):13777. https://doi.org/10.3390/ijms241813777
Chicago/Turabian StyleLee, Ji Hye, Yon-Suk Kim, and Kang Hyun Leem. 2023. "Citri Reticulatae Pericarpium Limits TLR-4-Triggered Inflammatory Response in Raw264.7 Macrophages by Activating RasGRP3" International Journal of Molecular Sciences 24, no. 18: 13777. https://doi.org/10.3390/ijms241813777