
Anti-Rheumatoid Arthritis Pharmacodynamic Substances Screening of Periploca forrestii Schltr.: Component Analyses In Vitro and In Vivo Combined with Multi-Technical Metabolomics
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Basic Physiological Parameters, Hematoxylin-Eosin (H&E) Staining, and Quantification of Serum RF, TNF-α, and IL-1β
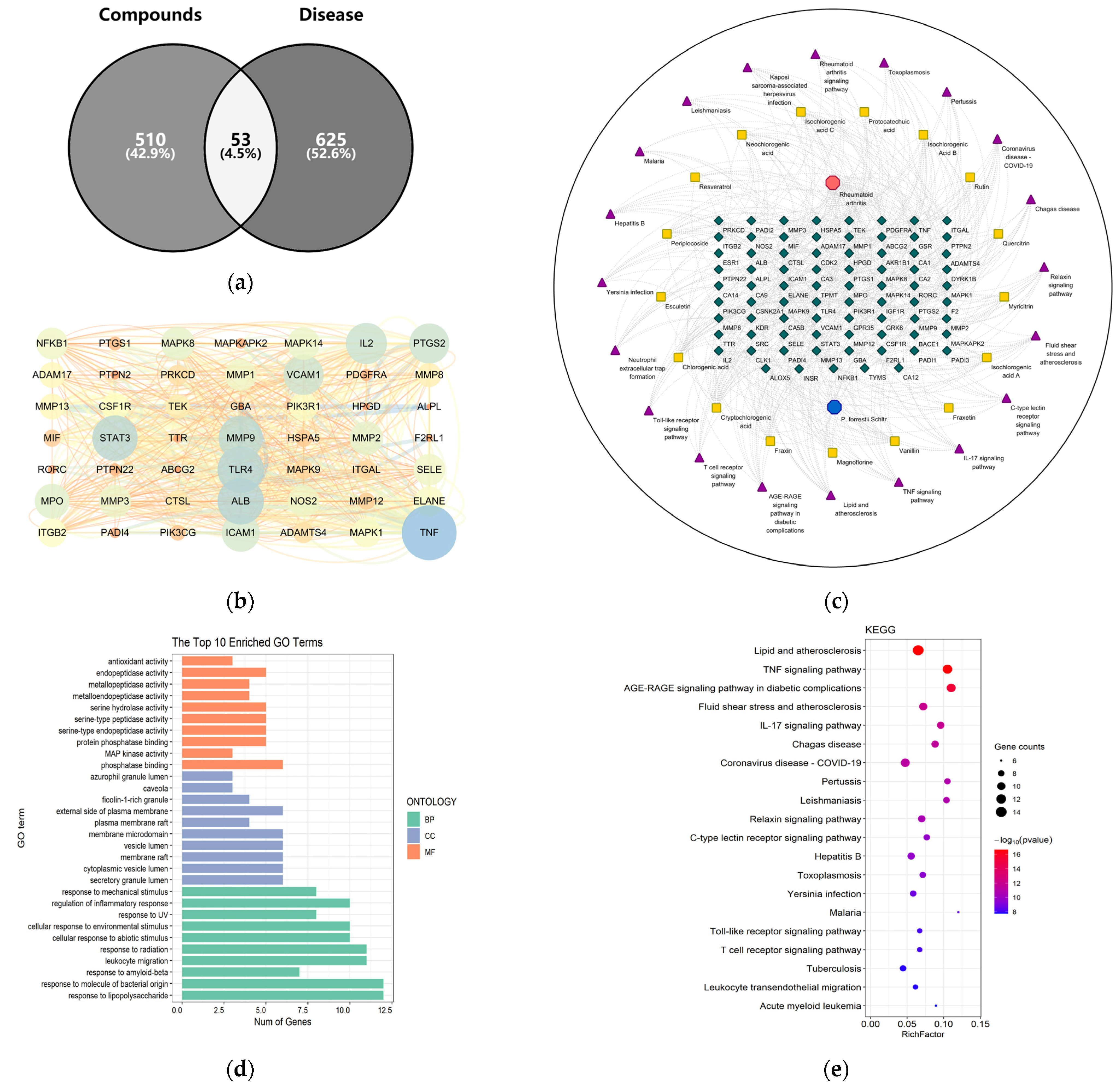
2.2. Component Analyses In Vitro and In Vivo and the Acquisition of Active Ingredient Groups
2.3. Multi-Tech Metabolomics Analyses
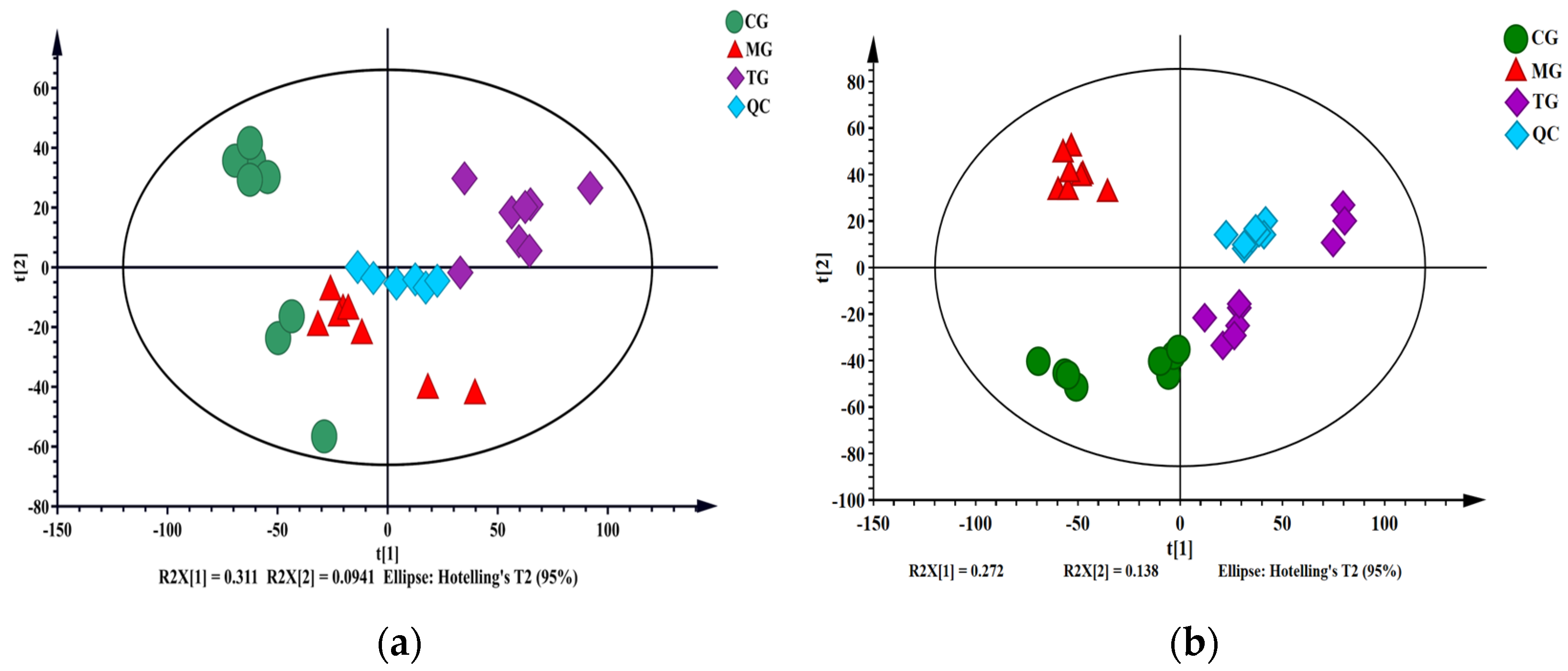
2.3.1. System Stability and Reproducibility Analyses of UPLC-Triple-TOF-MS/MS and GC-MS
2.3.2. Multivariate Statistical Analysis
2.3.3. Differential Metabolite Analysis
2.3.4. Analysis of Metabolic Pathways
2.4. Pharmacodynamic Substances of the Periploca forrestii Schltr. Extracts against Rheumatoid Arthritis
3. Discussion
4. Materials and Methods
4.1. Herbal and Chemicals
4.2. Periploca forrestii Schltr. Extraction
4.3. Model Construction and Drug Administration
4.4. Basic Physiological Parameters
4.5. Quantification of Serum TNF-α and IL-1β and Histopathological Analysis
4.6. In Vitro and In Vivo Chemical Composition Identification
4.6.1. In Vitro
4.6.2. In Vivo
4.6.3. Apparatus Condition
4.7. Network Pharmacology
4.8. Multi-Technical Metabolomics Analyses
4.8.1. UPLC-Triple-TOF-MS/MS Metabolomics Analysis
4.8.2. GC-MS Metabolomics Analysis
4.9. Anti-RA Pharmacodynamic Substances Screening of P. forrestii Extracts
4.10. Data Processing and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, L.; Zhong, X.; Zhu, Y.; Chen, B.; Yang, J.; Yu, C. Establishment and evaluation of adjuvant induced arthritis model in female Lewisrats. J. Shanxi Med. Univ. 2019, 50, 1453–1460. [Google Scholar]
- Zhao, Y. The Inhibitory Mechanism of Baicalein on Tumour Necrosis Factor-Alpha-Induced Proliferation of Human Rheumatoid Arthritis Fibroblast-like Synoviocytes. Master’s Thesis, Nanjing University, Nanjing, China, 2016. [Google Scholar]
- Gao, W. Pristimerin Inhibits Rheumatoid Arthritis Fibroblast-like Synoviocytes Proliferation and Underlying Mechanisms. Master’s Thesis, Southern Medical University, Shenzhen, China, 2016. [Google Scholar]
- Zhou, S.; Zhou, Y. Progress in Clinical Research on the Treatment of Rheumatoid Arthritis Based on Traditional Chinese Medicine Syndrome Differentiation. Clin. J. Tradit. Chin. Med. 2019, 31, 2035–2038. [Google Scholar]
- National Institute of Arthritis and Musculoskeletal and Skin Diseases. Handout on Health: Rheumatoid Arthritis. Available online: https://web.archive.org/web/20150630212818/ or http://niamsnih.gov/Health_Info/Rheumatic_Disease/default.asp; (accessed on 2 July 2015).
- Singh, J.A.; Wells, G.A.; Christensen, R.; Tanjong Ghogomu, E.; Maxwell, L.; Macdonald, J.K.; Filippini, G.; Skoetz, N.; Francis, D.; Lopes, L.C.; et al. Adverse effects of biologics: A network meta-analysis and Cochrane overview. Cochrane Database Syst. Rev. 2011, 2011, Cd008794. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Qin, X.; Fang, J.; Sun, X. Nanomedicines for the treatment of rheumatoid arthritis: State of art and potential therapeutic strategies. Acta Pharm. Sin. B 2021, 11, 1158–1174. [Google Scholar] [CrossRef]
- Shan, X.; Gong, X.; Li, J.; Wen, J.; Li, Y.; Zhang, Z. Current approaches of nanomedicines in the market and various stage of clinical translation. Acta Pharm. Sin. B 2022, 12, 3028–3048. [Google Scholar] [CrossRef]
- Radu, A.F.; Bungau, S.G. Nanomedical approaches in the realm of rheumatoid arthritis. Ageing Res. Rev. 2023, 87, 101927. [Google Scholar] [CrossRef]
- Oliveira, I.M.; Gonçalves, C.; Reis, R.L.; Oliveira, J.M. Engineering nanoparticles for targeting rheumatoid arthritis: Past, present, and future trends. Nano Res. 2018, 11, 4489–4506. [Google Scholar] [CrossRef]
- Xu, W.; Xing, F.J.; Dong, K.; You, C.; Yan, Y.; Zhang, L.; Zhao, G.; Chen, Y.; Wang, K. Application of traditional Chinese medicine preparation in targeting drug delivery system. Drug Deliv. 2015, 22, 258–265. [Google Scholar] [CrossRef]
- Liu, H.; Liu, Y.; Li, N.; Zhang, G.; Wang, M. Ginsenoside Rg3 liposome targeted delivery of dihydroartemisinin and paclitaxel for the treatment of triple-negative breast cancer. China J. Chin. Mater. Medica 2023, 48, 3472–3484. [Google Scholar]
- Li, D.; Li, Y.; Wu, D. Preparation of curcumin nanoemulsion and its protective effect on myocardial ischemia-reperfusion in rats. Chin. J. Comp. Med. 2020, 30, 97–103. [Google Scholar]
- Liu, J.; Xiao, M. Protective effect of aerobic exercise combined with paeonol PLGA nanoparticles on obese rats with hyperlipidemia. Chin. J. Comp. Med. 2019, 29, 88–93. [Google Scholar]
- Hu, B.; Lian, J.; Xu, X.; Wang, Y.; Zhang, Y.; Zhang, B. Preparation of glycyrrhizic acid liposomes and evaluation of liver targeting efficiency in mice. Acta Lab. Anim. Sci. Sin. 2015, 23, 401–405. [Google Scholar]
- Chen, Y.; Wu, J.; Yuan, L.; Liang, D.; Zhen, L. The Research Progress of Rheumatoid Arthritis Treatment by TCM. Pop. Sci. Technol. 2019, 21, 63–65. [Google Scholar]
- Lv, L.; Hu, Y.; Song, H.; Sun, X.; Zhou, A. Research progress in metabolomics of traditional Chinese medicine and ethnic medicine in the treatment of rheumatoid arthritis. Cent. South Pharm. 2023, 21, 192–198. [Google Scholar]
- Flora of China Editorial Committee. Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 1977; Volume 63. [Google Scholar]
- Feng, J.; Zhang, R.; Zhou, Y.; Chen, Z.; Tang, W.; Liu, Q.; Zuo, J.-P.; Zhao, W. Immunosuppressive pregnane glycosides from Periploca sepium and Periploca forrestii. Phytochem 2008, 69, 2716–2723. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Gan, X.; Gong, X.; Zhao, C.; Chen, H. Glycosides from Periploca forrestii. China J. Chin. Mater. Medica 2010, 35, 610–612. [Google Scholar]
- Zhao, S.; Zhang, B.; Xiong, D.; Zou, H.; Ma, X.; Sun, J.; Lu, Y.; Zheng, L.; Li, Y. Chemical constituents from Periploca Radix. Chin. Tradit. Herb. Drugs 2017, 48, 1513–1518. [Google Scholar]
- Liu, Y.; Jin, W.; Liu, G.; Yang, W.; Yan, Z.; Zhang, P. Chemical Constituents of Petroleum Ether Extract from Miao Medicine Periploca forrestii. J. Chin. Med. Mater. 2018, 41, 1622–1625. [Google Scholar]
- Wang, X.; Jiang, L.; He, Y.; Zheng, L.; Liu, T.; Dong, L.; Ma, X.; Li, Y. Effect of caffeoylquinic acid from Periploca forrestii on anti-rheumatoid arthritis in vitro and its mechanism. Chin. Pharmacol. Bull. 2018, 34, 1362–1367. [Google Scholar]
- Wang, X.; Yang, J.; Song, F.; Dong, L.; Liu, T.; Zheng, L.; Ma, X.; Li, Y. Effects of Caffeoylquinic Acid Derivative Fractions in Miao Medicine Periploca forrestiion Proliferation and Secretion of Inflammatory Cytokines in Human Rheumatoid Arthritis Fibroblast-like Synoviocytes MH7A. China Pharm. 2017, 28, 3949–3952. [Google Scholar]
- Qin, K.; Wang, B.; Chen, L.; Zhang, M.; Yang, G.; Shu, Y.; Cai, B. Perspective and application of metabonomics in modern study of traditional Chinese medicine. China J. Chin. Mater. Medica 2014, 39, 3010–3017. [Google Scholar]
- Lu, J.; Yang, J.; Xu, Y.; Yu, C. Advances in application of metabolomics in rheumatoid arthritis research. Chin. Pharmacol. Bull. 2019, 35, 1193–1196. [Google Scholar]
- Nemutlu, E.; Zhang, S.; Gupta, A.; Juranic, N.O.; Macura, S.I.; Terzic, A.; Jahangir, A.; Dzeja, P. Dynamic phosphometabolomic profiling of human tissues and transgenic models by 18O-assisted ³¹P NMR and mass spectrometry. Physiol Genom. 2012, 44, 386–402. [Google Scholar] [CrossRef] [PubMed]
- Nemutlu, E.; Zhang, S.; Xu, Y.Z.; Terzic, A.; Zhong, L.; Dzeja, P.D.; Cha, Y.M. Cardiac resynchronization therapy induces adaptive metabolic transitions in the metabolomic profile of heart failure. J. Card. Fail. 2015, 21, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Lanza, I.R.; Zhang, S.; Ward, L.E.; Karakelides, H.; Raftery, D.; Nair, K.S. Quantitative metabolomics by H-NMR and LC-MS/MS confirms altered metabolic pathways in diabetes. PLoS ONE 2010, 5, e10538. [Google Scholar] [CrossRef] [PubMed]
- Doerfler, H.; Lyon, D.; Nägele, T.; Sun, X.; Fragner, L.; Hadacek, F.; Egelhofer, V.; Weckwerth, W. Granger causality in integrated GC-MS and LC-MS metabolomics data reveals the interface of primary and secondary metabolism. Metabolomics 2013, 9, 564–574. [Google Scholar] [CrossRef]
- Shi, X. Serum Metabonomics of Pancreatic Cancer Based on LC-MS. Master’s Thesis, Harbin Institute of Technology, Harbin, China, 2015. [Google Scholar]
- Chen, C.; Yin, Q.; Tian, J.; Gao, X.; Qin, X.; Du, G.; Zhou, Y. Study on the antidepressant mechanism of Chaihu Bai Shaoyao based on liver metabolomics. Acta Pharma. Sin. 2020, 55, 941–949. [Google Scholar]
- Fan, Y. Study on Serum Metabolomics of Early Colorectal Cancer Based on Liquid Chromatography-Mass Spectrometry Technology. Master’s Thesis, Jilin University, Changchun, China, 2020. [Google Scholar]
- Chen, D.; Su, X.; Wang, N.; Li, Y.; Yin, H.; Li, L.; Li, L. Chemical Isotope Labeling LC-MS for Monitoring Disease Progression and Treatment in Animal Models: Plasma Metabolomics Study of Osteoarthritis Rat Model. Sci. Rep. 2017, 7, 40543. [Google Scholar] [CrossRef]
- Yang, X.; Yang, Z.; Li, S.; Deng, Y.; Yang, Y.; Man, Q.; Li, P. Rat plasma metabolomics in blood stasis model based on ultra performance liquid chromatography-quadrupole-time-of-flight mass spectrometry. Chin. J. Chromatogr. 2019, 37, 71–79. [Google Scholar] [CrossRef]
- Radu, A.F.; Bungau, S.G.; Negru, A.P.; Uivaraseanu, B.; Bogdan, M.A. Novel Potential Janus Kinase Inhibitors with Therapeutic Prospects in Rheumatoid Arthritis Addressed by In Silico Studies. Molecules 2023, 28, 4699. [Google Scholar] [CrossRef]
- Pan, J.; Wang, C.; Li, K.; Yuan, T.; Qin, X.; Wang, A.; Gong, Z.; Zheng, L. Pharmacokinetic-pharmacodynamic link model of Periploca forrestii based on rheumatiod arthritis rat model. Chin. Tradit. Herb. Drugs 2020, 51, 5194–5200. [Google Scholar]
- Obiri, D.D.; Osafo, N.; Ayande, P.G.; Antwi, A.O. Xylopia aethiopica (Annonaceae) fruit extract suppresses Freund’s adjuvant-induced arthritis in Sprague-Dawley rats. J. Ethnopharmacol. 2014, 152, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. ‘Metabonomics’: Understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef]
- Lee, J.W.; Su, Y.; Baloni, P.; Chen, D.; Pavlovitch-Bedzyk, A.J.; Yuan, D.; Duvvuri, V.R.; Ng, R.H.; Choi, J.; Xie, J.; et al. Integrated analysis of plasma and single immune cells uncovers metabolic changes in individuals with COVID-19. Nat. Biotechnol. 2022, 40, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Jacob, M.; Lopata, A.L.; Dasouki, M.; Abdel Rahman, A.M. Metabolomics toward personalized medicine. Mass Spectrom. Rev. 2019, 38, 221–238. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Ao, H.; Gao, H.; Zhu, Z. Metabolite biomarker discovery for human gastric cancer using dried blood spot mass spectrometry metabolomic approach. Sci. Rep. 2022, 12, 14632. [Google Scholar] [CrossRef]
- Wang, W.; Li, S.; Li, Z.; Zhang, J.; Fan, K.; Tan, G.; Ai, G.; Lam, S.M.; Shui, G.; Yang, Z.; et al. Harnessing the intracellular triacylglycerols for titer improvement of polyketides in Streptomyces. Nat. Biotechnol. 2020, 38, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wei, X.; Chen, W.; Lv, Y.; Wei, Y.; Zu, X.; Zhang, X. Application of metabolomics technology in the study of the compatibility laws of traditional Chinese medicine prescriptions. Chin. Tradit. Herb. Drugs 2022, 53, 1528–1539. [Google Scholar]
- Xu, T.; Feng, G.; Zhao, B.; Zhao, J.; Pi, Z.; Liu, S.; Song, F.; Liu, Z. A non-target urinary and serum metabolomics strategy reveals therapeutical mechanism of Radix Astragali on adjuvant-induced arthritis rats. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017, 1048, 94–101. [Google Scholar] [CrossRef]
- Zhang, B. Pharmacodynamic and Metabolomic Studies on Five Types of “Zhu” for Spleen Strengthening and Anti Rheumatism. Master’s Thesis, Liaoning University of Traditional Chinese Medicine, Shenyang, China, 2020. [Google Scholar]
- Xu, D. A metabolomics Study on the Effective Fraction of Lei-Ye-Mu-Dan Intervening in Collagen Induced Arthritis Rats. Master’s Thesis, Heilongjiang University of Chinese Medicine, Harbin, China, 2018. [Google Scholar]
- Tang, C.; Gan, Z.; Luo, S.; Yang, J.; Yu, M.; Zou, Z.; Zhang, Y. Study on the Mechanism of Action of Tibetan Medicine Extracts of Pterocarya pilosa in the Treatment of Rheumatoid Arthritis Based on Serum Metabolomics. China J. Chin. Mater. Medica 2022, 47, 1001–1008. [Google Scholar]
- Schober, C.; Schiller, J.; Pinker, F.; Hengstler, J.G.; Fuchs, B.; Fuchs, B. Lysophosphatidylethanolamine is–in contrast to–choline–generated under in vivo conditions exclusively by phospholipase A2 but not by hypochlorous acid. Bioorg. Chem. 2009, 37, 202–210. [Google Scholar] [CrossRef]
- Ding, X.; Hu, J.; Li, J.; Zhang, Y.; Shui, B.; Ding, Z.; Yao, L.; Fan, Y. Metabolomics analysis of collagen-induced arthritis in rats and interventional effects of oral tolerance. Anal. Biochem. 2014, 458, 49–57. [Google Scholar] [CrossRef]
- Surowiec, I.; Ärlestig, L.; Rantapää-Dahlqvist, S.; Trygg, J. Metabolite and Lipid Profiling of Biobank Plasma Samples Collected Prior to Onset of Rheumatoid Arthritis. PLoS ONE 2016, 11, e0164196. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Zhong, C. The role of ceramide in mitochondrial-dependent apoptosis induced by exogenous chemicals. Health Res. 2014, 43, 1031–1037. [Google Scholar]
- Lin, C.L.; Xu, R.; Yi, J.K.; Li, F.; Chen, J.; Jones, E.C.; Slutsky, J.B.; Huang, L.; Rigas, B.; Cao, J.; et al. Alkaline Ceramidase 1 Protects Mice from Premature Hair Loss by Maintaining the Homeostasis of Hair Follicle Stem Cells. Stem Cell Rep. 2017, 9, 1488–1500. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Cheng, C.; Zhang, X.; Peng, J. Inclusion of Soluble Fiber in the Gestation Diet Changes the Gut Microbiota, Affects Plasma Propionate and Odd-Chain Fatty Acids Levels, and Improves Insulin Sensitivity in Sows. Int. J. Mol. Sci. 2020, 21, 635. [Google Scholar] [CrossRef]
- Zhang, Y.; Xue, R.; Zhang, Z.; Yang, X.; Shi, H. Palmitic and linoleic acids induce ER stress and apoptosis in hepatoma cells. Lipids Health Dis. 2012, 11, 1. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, X.; Xie, J.; Hou, Z.; Li, Y. A GC-MS Based Metabonomics Study of Rheumatoid Arthritis and the Interventional Effects of the Simiaowan in Rats. Molecules 2015, 20, 21364–21372. [Google Scholar] [CrossRef]
- Pang, Z.; Wang, G.; Ran, N.; Lin, H.; Wang, Z.; Guan, X.; Yuan, Y.; Fang, K.; Liu, J.; Wang, F. Inhibitory Effect of Methotrexate on Rheumatoid Arthritis Inflammation and Comprehensive Metabolomics Analysis Using Ultra-Performance Liquid Chromatography-Quadrupole Time of Flight-Mass Spectrometry (UPLC-Q/TOF-MS). Int. J. Mol. Sci. 2018, 19, 2894. [Google Scholar] [CrossRef]
- Ren, S.; Shao, Y.; Zhao, X.; Hong, C.S.; Wang, F.; Lu, X.; Li, J.; Ye, G.; Yan, M.; Zhuang, Z.; et al. Integration of Metabolomics and Transcriptomics Reveals Major Metabolic Pathways and Potential Biomarker Involved in Prostate Cancer. Mol. Cell. Proteom. 2016, 15, 154–163. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhang, S.; Wang, P.; Chen, X.; Bi, J.; Cheng, L.; Zhang, X. A comprehensive review of the analysis and integration of omics data for SARS-CoV-2 and COVID-19. Brief. Bioinform. 2022, 23, bbab446. [Google Scholar] [CrossRef] [PubMed]
- Solovev, I.; Shaposhnikov, M.; Moskalev, A. Multi-omics approaches to human biological age estimation. Mech. Ageing Dev. 2020, 185, 111192. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.; Wang, C.; Li, K.; Qin, X.; Wang, A.; Huang, Y.; Li, Y.; Gong, Z.; Zheng, L. Pharmacokinetics of three active ingredients in Periploca forrestii extract after administrations in AA rats. Chin. Pharmacol. Bull. 2020, 36, 1270–1275. [Google Scholar]
- Wang, R.; Su, S.; Ma, T.; Liu, F.; Sun, S.; Gao, H. Effects of Qinjiao in Different Herb Pair Combinations on the AnklePathology and Expression of VEGF in Wind-damp-heat Arthralgia RA Model Rats. Tradit. Chin. Drug Res. Clin. Pharmacol. 2018, 29, 136–142. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | RT | Metabolites | VIP | Tend | |
|---|---|---|---|---|---|
| (min) | MG a | TG b | |||
| LC 1 | 1.87 | Indoleacrylic acid | 1.22 | ↓ | ↑ |
| LC 2 | 5.52 | Chenodeoxycholic Acid | 1.10 | ↓ | ↑ |
| LC 3 | 6.14 | Arachidonic acid | 2.27 | ↓ | ↑ |
| LC 4 | 6.17 | 10E,12Z-Octadecadienoic acid | 4.52 | ↑ | ↓ |
| LC 5 | 6.19 | Glycerophosphocholine | 1.27 | ↓ | ↑ |
| LC 6 | 6.19 | Iso-Valeraldehyde | 2.52 | ↓ | ↑ |
| LC 7 | 6.62 | LysoPA(0:0/16:0) | 1.07 | ↓ | ↑ |
| LC 8 | 6.87 | LysoPC(18:1(9Z)) | 1.82 | ↓ | ↑ |
| LC 9 | 6.92 | Choline | 1.43 | ↓ | ↑ |
| LC 10 | 8.09 | PG(18:3(6Z,9Z,12Z)/0:0) | 1.09 | ↓ | ↑ |
| LC 11 | 12.60 | Glucosylceramide (d18:1/16:0) | 1.50 | ↑ | ↓ |
| LC 12 | 13.99 | PC(22:4(7Z,10Z,13Z,16Z)/P-18:1(11Z)) | 1.09 | ↑ | ↓ |
| LC 13 | 14.01 | PC(15:0/20:0) | 1.01 | ↑ | ↓ |
| LC 14 | 14.03 | SM(d18:0/24:1(15Z)(OH)) | 1.33 | ↑ | ↓ |
| LC 15 | 14.06 | DG(18:0/20:4(5Z,8Z,11Z,14Z)/0:0) | 2.63 | ↓ | ↑ |
| GC 1 | 10.74 | Estragole | 1.42 | ↑ | ↓ |
| GC 2 | 10.85 | L-Valine | 1.30 | ↑ | ↓ |
| GC 3 | 11.57 | Butanoic acid | 1.30 | ↓ | ↑ |
| GC 4 | 12.22 | Urea | 2.18 | ↓ | ↑ |
| GC 5 | 12.36 | Glycerol | 1.92 | ↑ | ↓ |
| GC 6 | 12.56 | Aminoethanesulfonic acid | 1.22 | ↓ | ↑ |
| GC 7 | 13.10 | Glycine | 1.56 | ↓ | ↑ |
| GC 8 | 20.08 | L-phenylalanine | 1.72 | ↓ | ↑ |
| GC 9 | 21.48 | Hentriacontane | 1.25 | ↓ | ↑ |
| GC 10 | 23.47 | DL-Ornithine | 1.28 | ↑ | ↓ |
| GC 11 | 24.04 | 1,5-Anhydro-D-sorbitol | 1.79 | ↑ | ↓ |
| GC 12 | 24.36 | D-Fructose | 1.02 | ↓ | ↑ |
| GC 13 | 25.20 | D-Glucose | 1.07 | ↓ | ↑ |
| GC 14 | 25.28 | L-Lysine | 1.08 | ↓ | ↑ |
| GC 15 | 27.86 | Myo-Inositol | 1.05 | ↓ | ↑ |
| GC 16 | 29.15 | Heptacosane | 2.77 | ↓ | ↑ |
| GC 17 | 29.90 | 9,12-Octadecadienoic acid | 1.27 | ↑ | ↓ |
| GC 18 | 29.91 | Oleic acid | 1.48 | ↑ | ↓ |
| GC 19 | 33.06 | Eicosanoic acid | 1.06 | ↑ | ↓ |
| GC 20 | 34.91 | Hexadecanoic acid | 1.06 | ↓ | ↑ |
| GC 21 | 37.79 | Octadecanoic acid | 1.00 | ↑ | ↓ |
| GC 22 | 38.22 | Octadecane | 1.50 | ↓ | ↑ |
| GC 23 | 40.94 | Triacontane | 1.30 | ↓ | ↑ |
| No | Pathway Name | Match Status | p | −log(p) | Holm p | FDR | Impact |
|---|---|---|---|---|---|---|---|
| 1 | Biosynthesis of unsaturated fatty acids a,b | 6/36 | 1.02 × 10−5 | 4.9934 | 0.0009 | 0.0009 | 0 |
| 2 | Glycerophospholipid metabolism a | 5/36 | 1.57 × 10−4 | 3.8041 | 0.0130 | 0.0066 | 0.3257 |
| 3 | Linoleic acid metabolism a,b | 2/5 | 2.23 × 10−3 | 2.6508 | 0.1832 | 0.0626 | 1.0000 |
| 4 | Aminoacyl-tRNA biosynthesis b | 4/48 | 5.46 × 10−3 | 2.2631 | 0.4420 | 0.1146 | 0 |
| 5 | Glycerolipid metabolism a,b | 2/16 | 2.42 × 10−2 | 1.6169 | 1.0000 | 0.4059 | 0.2492 |
| 6 | Sphingolipid metabolism a | 2/21 | 4.03 × 10−2 | 1.3943 | 1.0000 | 0.5648 | 0.0385 |
| 7 | Phenylalanine, tyrosine and tryptophan biosynthesis b | 1/4 | 6.06 × 10−2 | 1.2177 | 1.0000 | 0.6348 | 0.5000 |
| 8 | Galactose metabolism b | 2/27 | 6.37 × 10−2 | 1.1956 | 1.0000 | 0.6348 | 0 |
| 9 | Phosphatidylinositol signaling system a,b | 2/28 | 6.80 × 10−2 | 1.1674 | 1.0000 | 0.6348 | 0.0389 |
| 10 | Glycine, serine and threonine metabolism a,b | 2/33 | 9.07 × 10−2 | 1.0425 | 1.0000 | 0.7600 | 0.2458 |
| 11 | Arachidonic acid metabolism a,b | 2/36 | 1.05 × 10−1 | 0.9779 | 1.0000 | 0.7600 | 0.3135 |
| 12 | Ascorbate and aldarate metabolism b | 1/8 | 1.18 × 10−1 | 0.9295 | 1.0000 | 0.7600 | 0 |
| 13 | Valine, leucine and isoleucine biosynthesis b | 1/8 | 1.18 × 10−1 | 0.9295 | 1.0000 | 0.7600 | 0 |
| 14 | Biotin metabolism b | 1/10 | 1.45 × 10−1 | 0.8390 | 1.0000 | 0.8113 | 0 |
| 15 | Phenylalanine metabolism b | 1/10 | 1.45 × 10−1 | 0.8390 | 1.0000 | 0.8113 | 0.3571 |
| 16 | Primary bile acid biosynthesis a,b | 2/46 | 1.58 × 10−1 | 0.8024 | 1.0000 | 0.8276 | 0.0076 |
| 17 | alpha-Linolenic acid metabolism a | 1/13 | 1.84 × 10−1 | 0.7346 | 1.0000 | 0.9105 | 0 |
| 18 | Arginine biosynthesis b | 1/14 | 1.97 × 10−1 | 0.7055 | 1.0000 | 0.9193 | 0 |
| 19 | Butanoate metabolism b | 1/15 | 2.10 × 10−1 | 0.6787 | 1.0000 | 0.9264 | 0 |
| 20 | Pantothenate and CoA biosynthesis b | 1/19 | 2.58 × 10−1 | 0.5886 | 1.0000 | 1.0000 | 0 |
| 21 | Ether lipid metabolism a | 1/20 | 2.70 × 10−1 | 0.5694 | 1.0000 | 1.0000 | 0 |
| 22 | Lysine degradation b | 1/25 | 3.25 × 10−1 | 0.4880 | 1.0000 | 1.0000 | 0 |
| 23 | Glycolysis/Gluconeogenesis b | 1/26 | 3.36 × 10−1 | 0.4740 | 1.0000 | 1.0000 | 0.0002 |
| 24 | Glutathione metabolism b | 1/28 | 3.56 × 10−1 | 0.4480 | 1.0000 | 1.0000 | 0.088 |
| 25 | Porphyrin and chlorophyll metabolism b | 1/30 | 3.77 × 10−1 | 0.4241 | 1.0000 | 1.0000 | 0 |
| 26 | Inositol phosphate metabolism b | 1/30 | 3.77 × 10−1 | 0.4241 | 1.0000 | 1.0000 | 0.1294 |
| 27 | Glyoxylate and dicarboxylate metabolism b | 1/32 | 3.96 × 10−1 | 0.4021 | 1.0000 | 1.0000 | 0.1058 |
| 28 | Amino sugar and nucleotide sugar metabolism b | 1/37 | 4.42 × 10−1 | 0.3541 | 1.0000 | 1.0000 | 0 |
| 29 | Fatty acid elongation b | 1/39 | 4.60 × 10−1 | 0.3372 | 1.0000 | 1.0000 | 0 |
| 30 | Fatty acid degradation b | 1/39 | 4.60 × 10−1 | 0.3372 | 1.0000 | 1.0000 | 0 |
| 31 | Valine, leucine and isoleucine degradation b | 1/40 | 4.69 × 10−1 | 0.3292 | 1.0000 | 1.0000 | 0 |
| 32 | Fatty acid biosynthesis b | 1/47 | 5.25 × 10−1 | 0.2798 | 1.0000 | 1.0000 | 0.0147 |
| 33 | Purine metabolism b | 1/65 | 6.45 × 10−1 | 0.1903 | 1.0000 | 1.0000 | 0 |
| 34 | Biosynthesis of unsaturated fatty acids a,b | 6/36 | 1.02 × 10−5 | 4.9934 | 0.0009 | 0.0009 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, J.; Zhou, Z.; Zhou, Y.; Liu, T.; Li, Y.; Gong, Z.; Jin, Y.; Zheng, L.; Huang, Y. Anti-Rheumatoid Arthritis Pharmacodynamic Substances Screening of Periploca forrestii Schltr.: Component Analyses In Vitro and In Vivo Combined with Multi-Technical Metabolomics. Int. J. Mol. Sci. 2023, 24, 13695. https://doi.org/10.3390/ijms241813695
Sun J, Zhou Z, Zhou Y, Liu T, Li Y, Gong Z, Jin Y, Zheng L, Huang Y. Anti-Rheumatoid Arthritis Pharmacodynamic Substances Screening of Periploca forrestii Schltr.: Component Analyses In Vitro and In Vivo Combined with Multi-Technical Metabolomics. International Journal of Molecular Sciences. 2023; 24(18):13695. https://doi.org/10.3390/ijms241813695
Chicago/Turabian StyleSun, Jia, Zuying Zhou, Yang Zhou, Ting Liu, Yueting Li, Zipeng Gong, Yang Jin, Lin Zheng, and Yong Huang. 2023. "Anti-Rheumatoid Arthritis Pharmacodynamic Substances Screening of Periploca forrestii Schltr.: Component Analyses In Vitro and In Vivo Combined with Multi-Technical Metabolomics" International Journal of Molecular Sciences 24, no. 18: 13695. https://doi.org/10.3390/ijms241813695