
Non-Covalent Interactions between dUTP C5-Substituents and DNA Polymerase Decrease PCR Efficiency
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. PCR Efficiency Using Different DNA Polymerases and dUTPs C5-Modified with Bulky Substituents
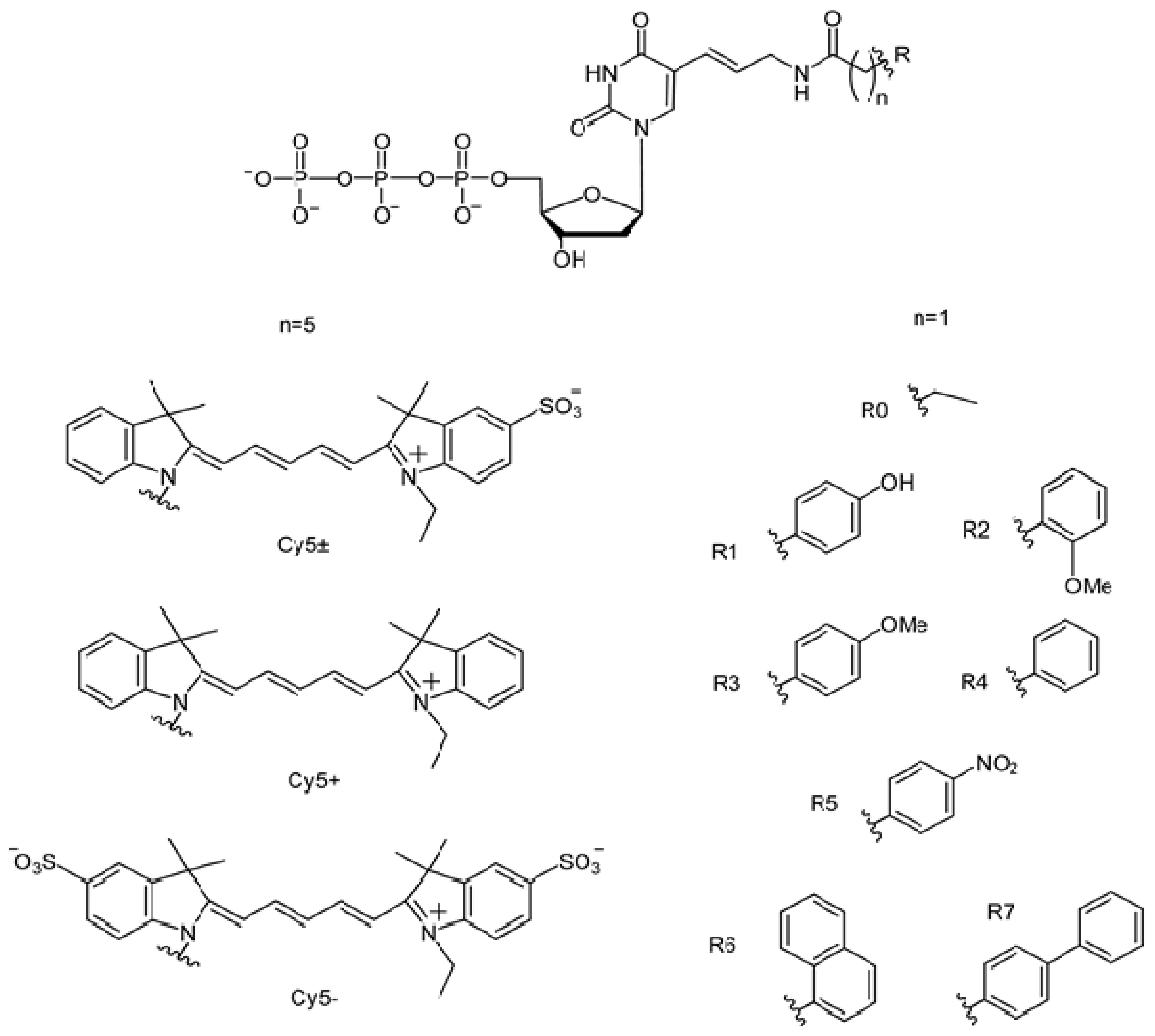
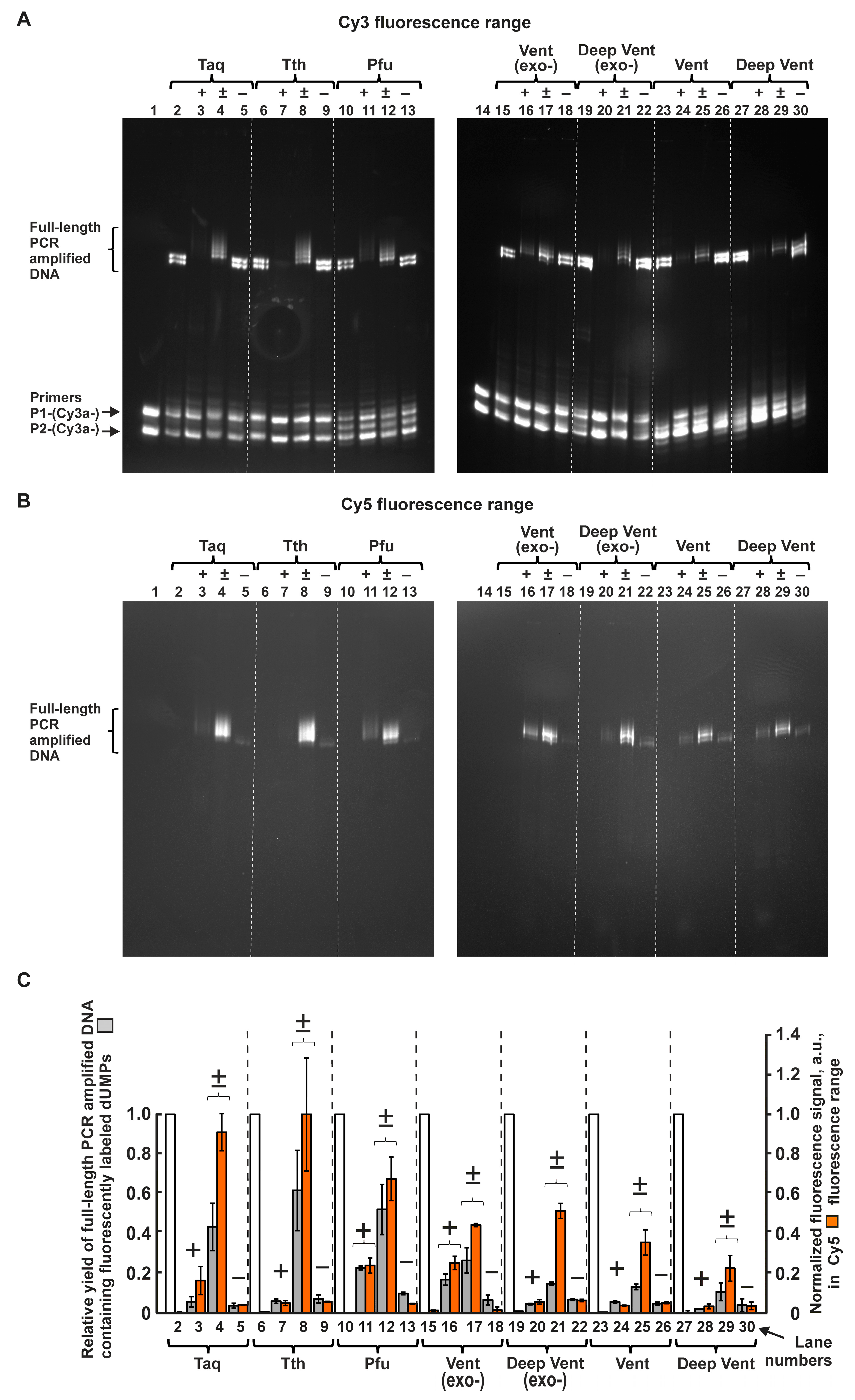
2.1.1. The Structures of C5-Attached dUTP Substituents and Electrophoretic Separation of PCR Products
2.1.2. Inhibiting Effect of Modified dUTPs in the Reaction Mixture on the Yield of PCR Product Consisting Only of Natural Nucleotides
2.1.3. Ratings of Seven A and B Family DNA Polymerases in Ability to Use Modified dUTPs in PCR
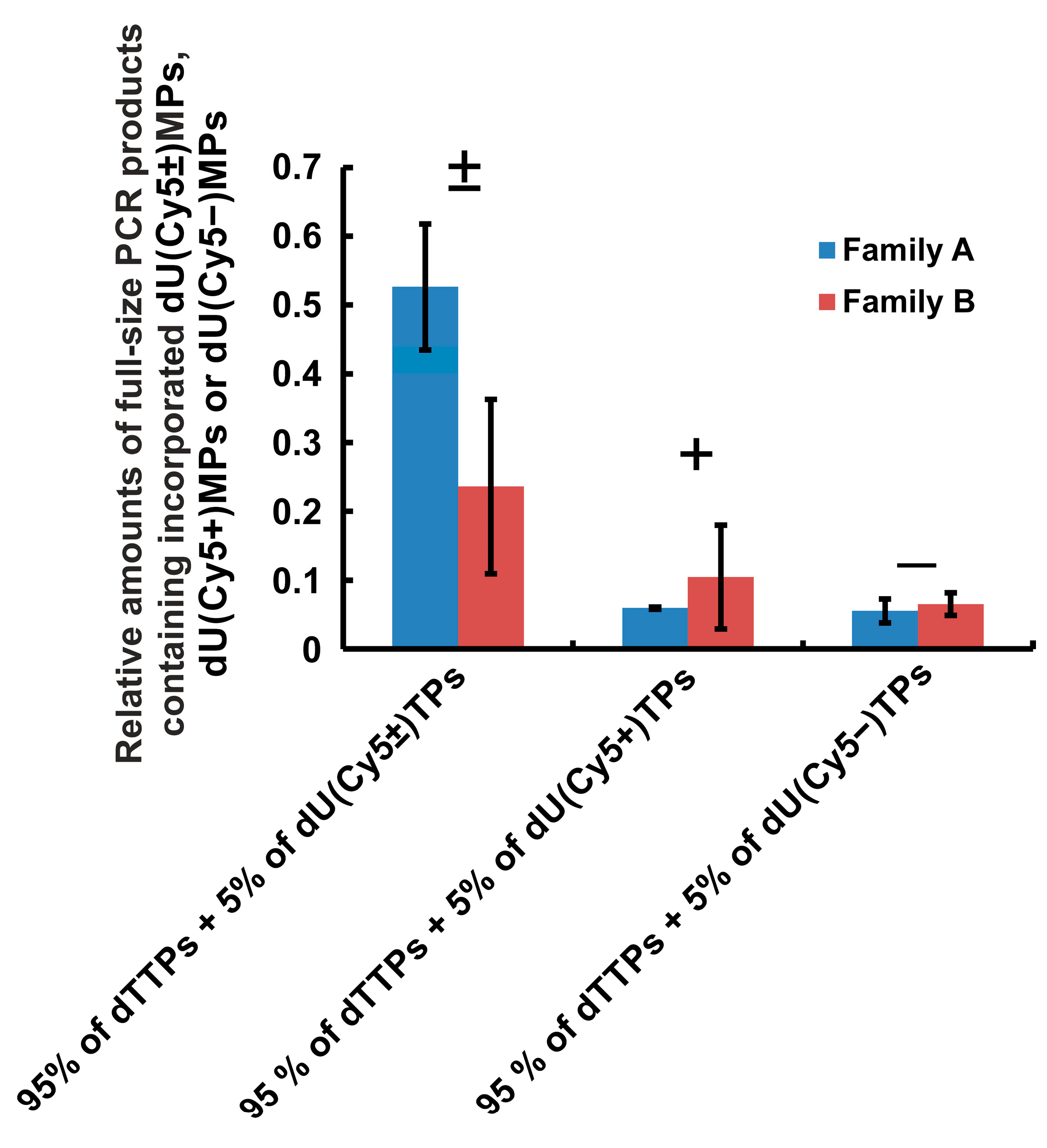
2.1.4. General Rating of dUTPs C5-Modified with Cy5 Dye Analogs According to PCR Efficiency with DNA Polymerases of the A and B Families
2.2. Structural Factors Reducing the PCR Efficiency When dUTPs C5-Modified by Various Low Molecular Weight Substituents Are Used
- Small (126–251 Da) substituents consisting of -CH=CH-CH2-NHCO-CH2- linker and R0–R7 functional groups;
- Bulky (536–694 Da) substituents consisting of -CH=CH-CH2-NHCO-(CH2)5- linker and the analogs of Cy5 dye.
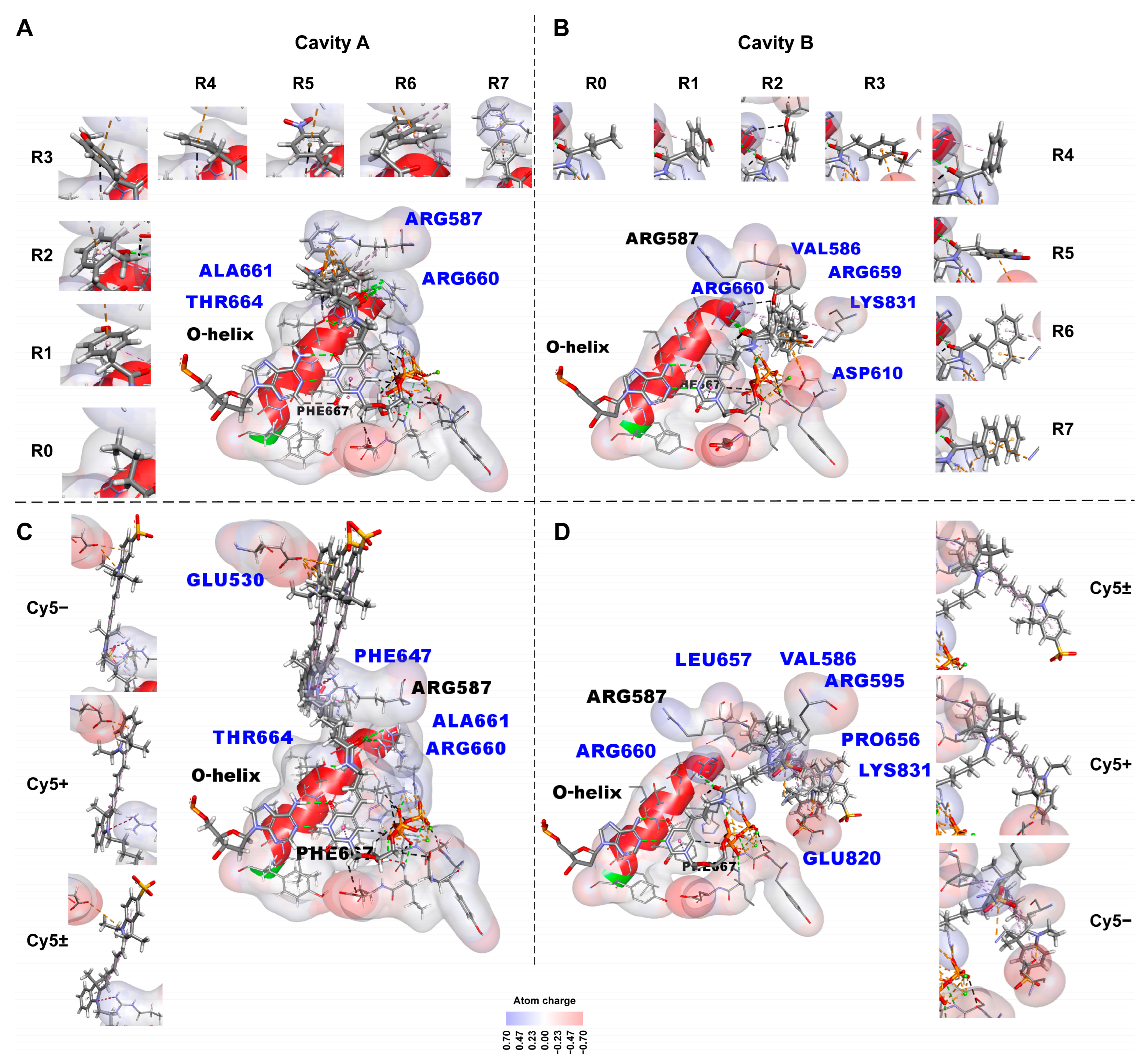
2.2.1. Non-Covalent Interactions between KlenTaq Polymerase and the Substituents Containing Small Functional Groups R0–R7
- hydrogen,
- carbon–hydrogen,
- π–donor hydrogen,
- π–cation,
- π–anion, and
- π–alkyl bonds between the chemical groups of the substituents and the chemical groups of the amino acid residues of the KlenTaq polymerase obtained using the Discovery Studio program. These interactions are listed in Table S2. Table S2 also presents the mean number of non-covalent interactions between each substituent, and KlenTaq polymerase averaged over two types of localization (via cavity A and cavity B).
- -
- Hydrogen bonds by green dotted lines;
- -
- Carbon and π–donor hydrogen bonds by black dotted lines;
- -
- Electrostatic interactions, including π–cationic and π–anionic, by orange dotted lines;
- -
- Hydrophobic π–alkyl interactions by purple dotted lines;
- -
- Electrostatic attractive charge bonds by orange dotted lines;
- -
- π–lone pair bond by green dotted line;
- -
- π–sulfur bond by yellow dotted line;
- -
- Hydrophobic π–π stacking bond is denoted by a pink dotted line, alkyl bonds by purple dotted lines, and the π–sigma bond by a dark purple dotted line. The bonds can be seen more in the Discovery Studio program [49] using dsv files with these 3D structures in the supplementary data. An unfavorable positive–positive interaction between the nitrogen atom close to the linker indolyl ring of the substituent and the nitrogen atom of Arg587 is marked by a red dotted line in C.
2.2.2. Non-Covalent Interactions between KlenTaq Polymerase and Bulky Substituents
- hydrogen,
- electrostatic (attractive charge bonds, π–anion bonds),
- π–lone pair bond,
- π–sulfur bond, and
- hydrophobic bonds (π–π-stacked, π–sigma, alkyl, and π–alkyl bonds), which are also listed in Table S3. Table S3 also presents the mean numbers of non-covalent bonds between the chemical groups of the substituent, and the chemical groups of the KlenTaq polymerase amino acid residues averaged over two types of localization (via cavity A and cavity B) for each substituent.
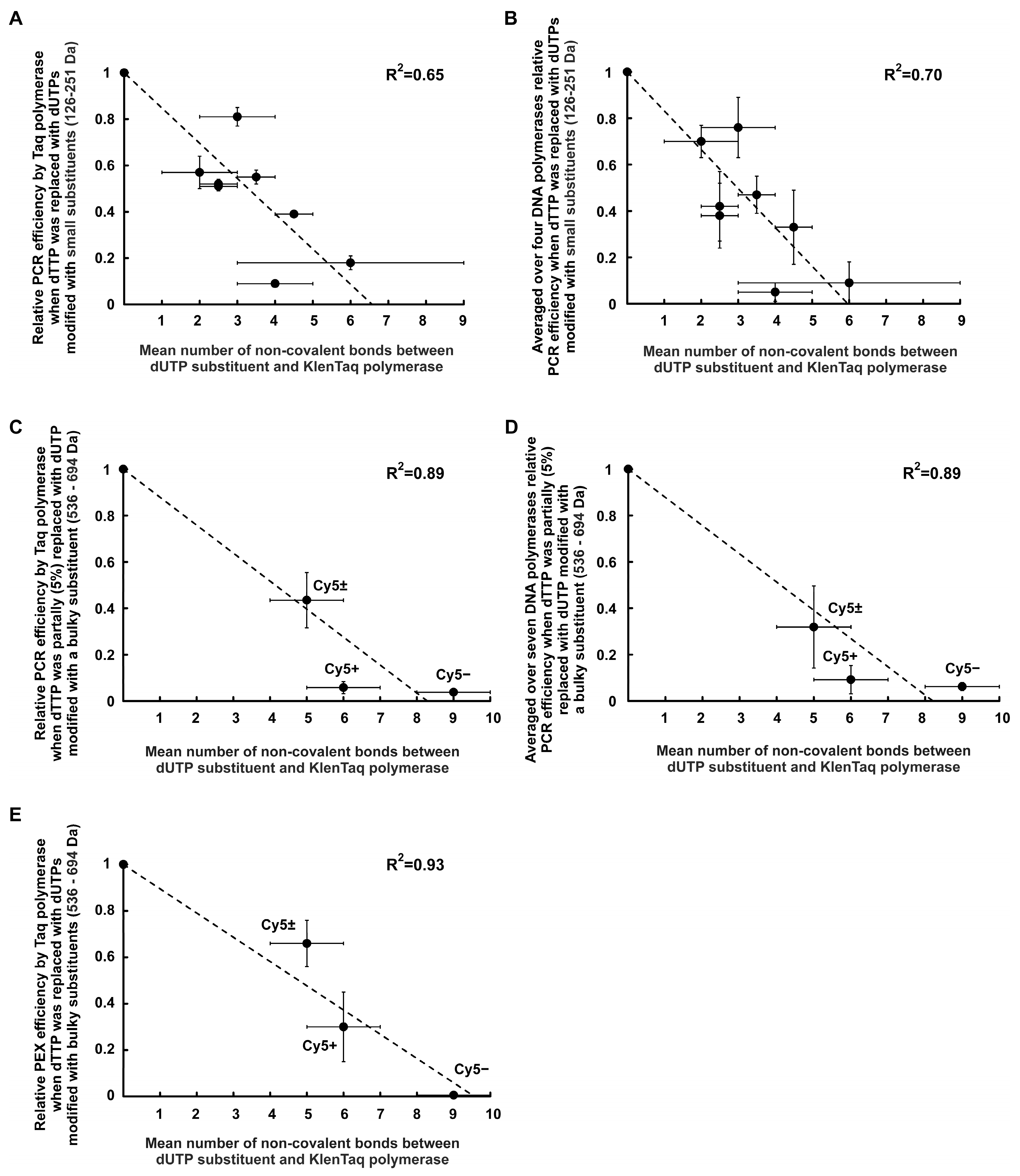
2.2.3. Negative Correlation between PCR Efficiency and the Number of Non-Covalent Bonds between the dUTP C5-Substituents and DNA Polymerase
Correlations for dUTPs C5-Modified with R0–R7 Functional Groups
Correlations for dUTPs C5-Modified with the Bulky Substituents
- For dU(Cy5±)TP:
- relative PCR efficiency—0.44 ± 0.12; mean number of non-covalent bonds—(5 ± 1);
- For dU(Cy5+)TP:
- relative PCR efficiency—0.06 ± 0.03; mean number of non-covalent bonds—(6 ± 1);
- For dU(Cy5–)TP:
- relative PCR efficiency—0.04 ± 0.01; mean number of non-covalent bonds—(9 ± 1).
2.2.4. Negative Correlation between PEX Efficiency and the Number of Non-Covalent Bonds Formed by dUTP C5-Substituents with the Taq Polymerase
2.2.5. The Similarity of the Local Environments of the dUTP C5-Substituents Localized in the Active Center of Different Polymerases
2.2.6. Analysis of the Linker Parts of the dUTP C5-Substituents in Non-Covalent Interactions with Taq Polymerase
2.2.7. Analysis of the Functional Groups R0–R7 of the dUTP C5-Substituents in Non-Covalent Interactions with Taq Polymerase
2.2.8. Analysis of Bulky dUTP C5-Substituents in Non-Covalent Interactions with Taq Polymerase
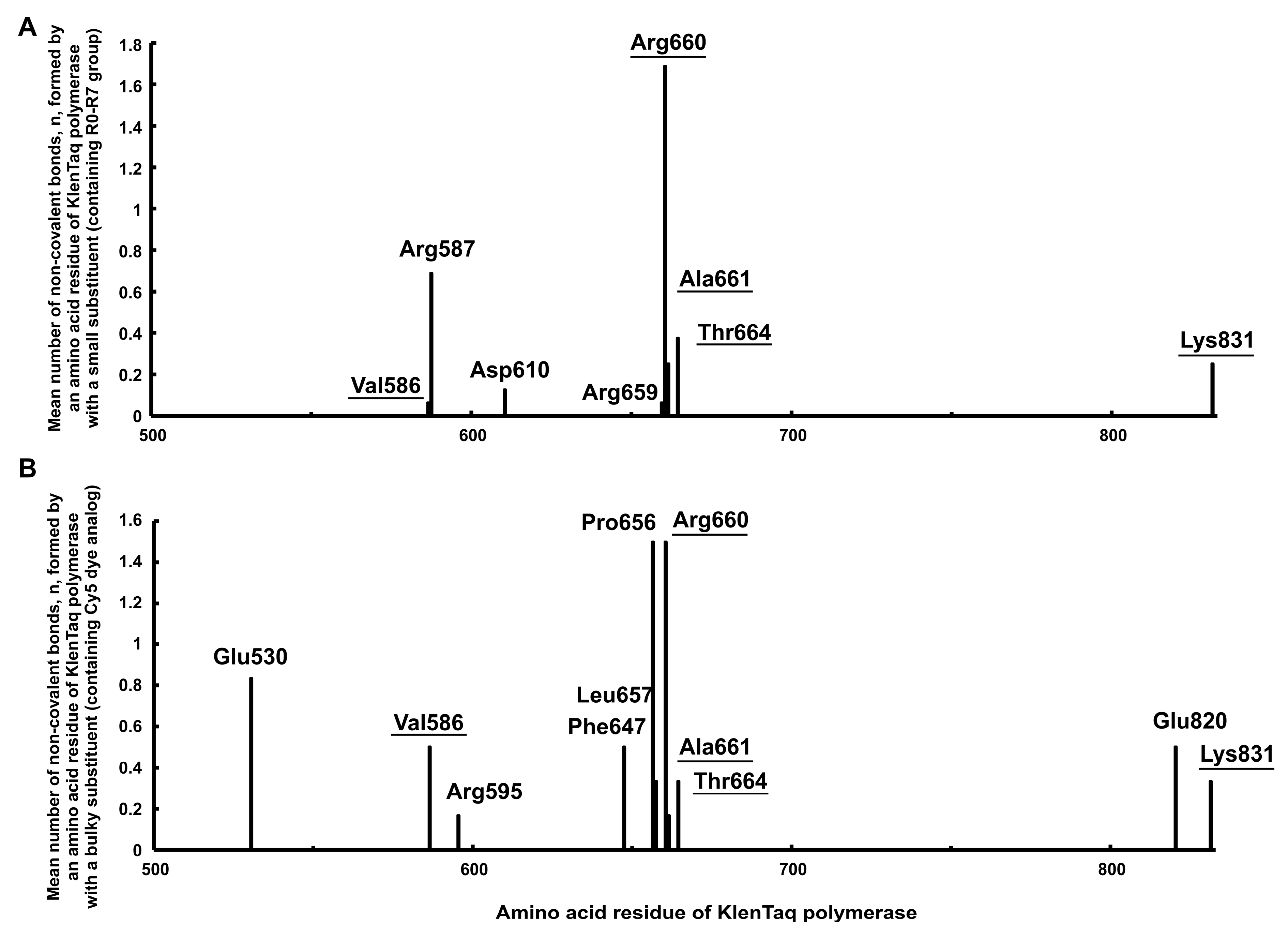
2.2.9. Analysis of Amino acid Residues Capable of Forming Non-Covalent Bonds with the dUTP-Attached Substituents
3. Discussion
3.1. Negative Correlation between the PCR Efficiency and the Number of Non-Covalent Bonds between the dUTP C5-Substituents and DNA Polymerase
3.2. The Similarity of the Local Environments for dNTP Substituents Localized in the Active Centers of Different Polymerases
3.3. The Role of Non-Covalent Interactions between Low Molecular Weight Substituents of the Modified dNTPs and the DNA Polymerase in Enzymatic Incorporation Efficiency
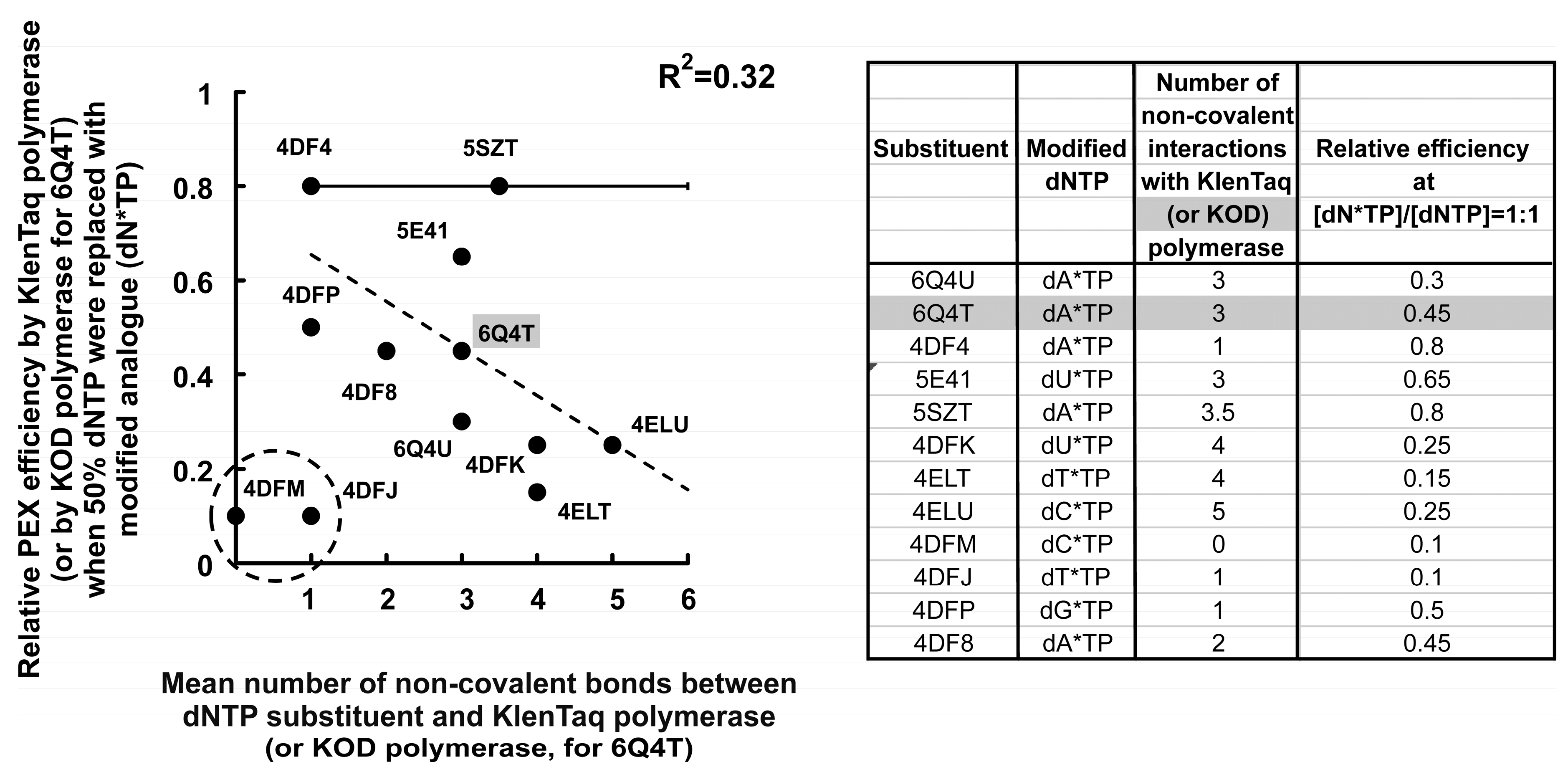
3.3.1. Incorporation Efficiency of dTspinMP and dTdendMP and X-ray Structural Data Obtained by Marx A. et al. Analyzed in the Discovery Studio Program
3.3.2. Competitive Incorporation of the Modified dNTPs and the X-ray Structural Data for KlenTaq Polymerase–DNA–(Modified dNTP) Complexes
3.4. Analysis of Amino Acid Residues of the KlenTaq Polymerase Capable of Forming Non-Covalent Bonds with the dUTP C5-Substituents
4. Materials and Methods
4.1. dUTPs Modified at C5-Position with Bulky or Small Aromatic Substituents
4.2. The DNA Template and Primers
4.3. PCR and Electrophoresis
- Thermus aquaticus (Taq);
- Thermus thermophilus (Tth);
- Pyrococcus furiosus (Pfu) DNA polymerases (Sileks, Badenweiler, Germany) as well as
- Thermococcus litoralis (Vent);
- Pyrococcus GB-D strain (Deep Vent) DNA polymerases and forms of the native DNA polymerases;
- Thermococcus litoralis (Vent (exo-)); and
- Pyrococcus GB-D strain (Deep Vent (exo-)) that had been genetically engineered to eliminate the 3′→5′ proofreading exonuclease activity (New England Biolabs, Ipswich, UK).
4.4. Quantitative Analysis of Electrophoretic Bands Containing PCR-Amplified Full-Length DNA Fragments
4.5. Computer Modeling of 3D Structures of Triple Complexes Consisting of KlenTaq Polymerase–DNA–(Modified dUTP)
4.6. KlenTaq Polymerase Amino Acid Residues’ Ability to Form Non-Covalent Interactions with the dUTP Substituents Attached at the C5 Position of the Pyrimidine Ring Localized in the Active Center of the Enzyme
5. Conclusions
- (i)
- The PCR efficiency in the presence of dUTPs modified with low molecular weight substituents (126–694 Da) decreases with an increase in the number of non-covalent bonds between the substituents and the DNA polymerase (about 15% decrease per one extra non-covalent bond). The decrease in the efficiency of PCR can be explained by the necessity of spending extra energy to overcome these additional bonds in order to release the polymerase molecule for further moving along the DNA primer/template after the formation of a phosphodiester bond;
- (ii)
- The number of non-covalent bonds between the dNTP substituents and polymerase amino acid residues can be considered as a variable parameter for regulation of enzyme activity. This number of non-covalent bonds can be estimated by modeling with the help of the highly efficient Discovery Studio software and can be useful in future design and synthesis of dNTP derivatives with new polymerase-specific properties;
- (iii)
- The obtained results demonstrate the similarity of all the tested DNA polymerases of the A and B families in the ability to use a substrate that mimics dTTP—the dUTP modified with a certain substituent. One can assume that, despite the varieties in the amino acid chains forming the active centers of different polymerases, the chemical groups of the amino acid residues facing the spaces of the active centers as well as those facing the adjacent regions (possibly forming cavities similar to the cavities A and B of the KlenTaq polymerase) form similar local environments not only for dNTPs (which is well known), but also for their substituents.
- (iv)
- The ten amino acid residues of Taq polymerase (Glu530, Val586, Arg595, Asp610, Phe647, Pro656, Leu657, Arg659, Glu820, and Lys831) are first predicted as capable of forming non-covalent bonds with dNTP substituents. The abovementioned amino acid residues can be considered the useful targets in directed mutagenesis to create mutant polymerases with desirable specificity to the modified dNTP substituents.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Walsh, J.M.; Beuning, P.J. Synthetic Nucleotides as Probes of DNA Polymerase Specificity. J. Nucleic Acids. 2012, 2012, 530963. [Google Scholar] [CrossRef]
- Hollenstein, M. Nucleoside triphosphates—building blocks for the modification of nucleic acids. Molecules 2012, 17, 13569–13591. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.-J.; Yang, W.; Tsai, M.-D. How DNA polymerases catalyse replication and repair with contrasting fidelity. Nat. Rev. Chem. 2017, 1, 0068. [Google Scholar] [CrossRef]
- Perlíková, P.; Hocek, M. Pyrrolo[2,3-d]pyrimidine (7-deazapurine) as a privileged scaffold in design of antitumor and antiviral nucleosides. Med. Res. Rev. 2017, 37, 1429–1460. [Google Scholar] [CrossRef] [PubMed]
- Tokarenko, A.; Lišková, B.; Smoleń, S.; Táborská, N.; Tichý, M.; Gurská, S.; Perlíková, P.; Frydrych, I.; Tloušt’ová, E.; Znojek, P.; et al. Synthesis and Cytotoxic and Antiviral Profiling of Pyrrolo- and Furo-Fused 7-Deazapurine Ribonucleosides. J Med Chem. 2018, 61, 9347–9359. [Google Scholar] [CrossRef]
- Tichý, M.; Pohl, R.; Xu, H.Y.; Chen, Y.L.; Yokokawa, F.; Shi, P.Y.; Hocek, M. Synthesis and antiviral activity of 4,6-disubstituted pyrimido[4,5-b]indole ribonucleosides. Bioorg Med Chem. 2012, 20, 6123–6133. [Google Scholar] [CrossRef]
- Fleuti, M.; Bártová, K.; Slavětínská, L.P.; Tloušt’ová, E.; Tichý, M.; Gurská, S.; Pavliš, P.; Džubák, P.; Hajdúch, M.; Hocek, M. Synthesis and Biological Profiling of Pyrazolo-Fused 7-Deazapurine Nucleosides. J. Org. Chem. 2020, 85, 10539–10551. [Google Scholar] [CrossRef]
- Gold, L.; Ayers, D.; Bertino, J.; Bock, C.; Bock, A.; Brody, E.N.; Carter, J.; Dalby, A.B.; Eaton, B.E.; Fitzwater, T.; et al. Aptamer-based multiplexed proteomic technology for biomarker discovery. PLoS ONE 2010, 5, e15004. [Google Scholar] [CrossRef]
- Kuwahara, M.; Sugimoto, N. Molecular evolution of functional nucleic acids with chemical modifications. Molecules. 2010, 15, 5423–5444. [Google Scholar] [CrossRef]
- Gold, L.; Walker, J.J.; Wilcox, S.K.; Williams, S. Advances in human proteomics at high scale with the SOMAscan proteomics platform. N. Biotechnol. 2012, 29, 543–549. [Google Scholar] [CrossRef]
- Famulok, M.; Mayer, G. Aptamers and SELEX in Chemistry & Biology. Chem. Biol. 2014, 21, 1055–1058. [Google Scholar] [PubMed]
- Drolet, D.W.; Green, L.S.; Gold, L.; Janjic, N. Fit for the Eye: Aptamers in Ocular Disorders. Nucleic Acid. Ther. 2016, 26, 127–146. [Google Scholar] [CrossRef]
- Gawande, B.N.; Rohloff, J.C.; Carter, J.D.; von Carlowitz, I.; Zhang, C.; Schneider, D.J.; Janjic, N. Selection of DNA aptamers with two modified bases. Proc. Natl. Acad. Sci. USA 2017, 114, 2898–2903. [Google Scholar] [CrossRef] [PubMed]
- Ladju, R.B.; Pascut, D.; Massi, M.N.; Tiribelli, C.; Sukowati, C.H. Aptamer: A potential oligonucleotide nanomedicine in the diagnosis and treatment of hepatocellular carcinoma. Oncotarget 2017, 9, 2951–2961. [Google Scholar] [CrossRef]
- Zhu, G.; Chen, X. Aptamer-based targeted therapy. Adv. Drug. Deliv. Rev. 2018, 134, 65–78. [Google Scholar] [CrossRef]
- Rose, K.M.; Alves Ferreira-Bravo, I.; Li, M.; Craigie, R.; Ditzler, M.A.; Holliger, P.; DeStefano, J.J. Selection of 2′-Deoxy-2′-Fluoroarabino Nucleic Acid (FANA) Aptamers That Bind HIV-1 Integrase with Picomolar Affinity. ACS Chem. Biol. 2019, 14, 2166–2175. [Google Scholar] [CrossRef]
- Minagawa, H.; Kataoka, Y.; Kuwahara, M.; Horii, K.; Shiratori, I.; Waga, I. A high affinity modified DNA aptamer containing base-appended bases for human β-defensin. Anal. Biochem. 2020, 594, 113627. [Google Scholar] [CrossRef] [PubMed]
- Minagawa, H.; Kataoka, Y.; Fujita, H.; Kuwahara, M.; Horii, K.; Shiratori, I.; Waga, I. Modified DNA Aptamers for C-Reactive Protein and Lactate Dehydrogenase-5 with Sub-Nanomolar Affinities. Int. J. Mol. Sci. 2020, 21, 2683. [Google Scholar] [CrossRef]
- Schmitz, A.; Weber, A.; Bayin, M.; Breuers, S.; Fieberg, V.; Famulok, M.; Mayer, G. A SARS-CoV-2 Spike Binding DNA Aptamer that Inhibits Pseudovirus Infection by an RBD-Independent Mechanism. Angew. Chem. Weinheim. Bergstr. Ger. 2021, 133, 10367–10373. [Google Scholar] [CrossRef]
- McKenzie, L.K.; El-Khoury, R.; Thorpe, J.D.; Damha, M.J.; Hollenstein, M. Recent progress in non-native nucleic acid modifications. Chem. Soc. Rev. 2021, 50, 5126–5164. [Google Scholar] [CrossRef]
- Jager, S.; Rasched, G.; Kornreich-Leshem, H.; Engeser, M.; Thum, O.; Famulok, M. A versatile toolbox for variable DNA functionalization at high density. J. Am. Chem. Soc. 2005, 127, 15071–15082. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, M.; Nagashima, J.; Hasegawa, M.; Tamura, T.; Kitagata, R.; Hanawa, K.; Hososhima, S.; Kasamatsu, T.; Ozaki, H.; Sawai, H. Systematic characterization of 2′-deoxynucleoside-5′-triphosphate analogs as substrates for DNA polymerases by polymerase chain reaction and kinetic studies on enzymatic production of modified DNA. Nucl. Acids Res. 2006, 34, 5383–5394. [Google Scholar] [CrossRef]
- Vaught, J.D.; Bock, C.; Carter, J.; Fitzwater, T.; Otis, M.; Schneider, D.; Rolando, J.; Waugh, S.; Wilcox, S.K.; Eaton, B.E. Expanding the chemistry of DNA for in vitro selection. J. Am. Chem. Soc. 2010, 132, 4141–4151. [Google Scholar] [CrossRef] [PubMed]
- Hocek, M. Synthesis of base-modified 2′-deoxyribonucleoside triphosphates and their use in enzymatic synthesis of modified DNA for applications in bioanalysis and chemical biology. J. Org. Chem. 2014, 79, 9914–9921. [Google Scholar] [CrossRef] [PubMed]
- Hollenstein, M. Generation of long, fully modified, and serum-resistant oligonucleotides by rolling circle amplification. Org. Biomol. Chem. 2015, 13, 9820–9824. [Google Scholar] [CrossRef] [PubMed]
- Ménová, P.; Cahová, H.; Vrábel, M.; Hocek, M. Synthesis of Base-Modified dNTPs Through Cross-Coupling Reactions and Their Polymerase Incorporation to DNA. Methods Mol. Biol. 2019, 1973, 39–57. [Google Scholar]
- Röthlisberger, P.; Levi-Acobas, F.; Leumann, C.J.; Hollenstein, M. Enzymatic synthesis of biphenyl-DNA oligonucleotides. Bioorg. Med. Chem. 2020, 28, 115487. [Google Scholar] [CrossRef] [PubMed]
- Zasedateleva, O.A.; Surzhikov, S.A.; Shershov, V.E.; Miftakhov, R.A.; Yurasov, D.A.; Kuznetsova, V.E.; Chudinov, A.V. PCR incorporation of dUMPs modified with aromatic hydrocarbon substituents of different hydrophilicities: Synthesis of C5-modified dUTPs and PCR studies using Taq, Tth, Vent (exo-) and Deep Vent (exo-) polymerases. Bioorg. Chem. 2020, 99, 103829. [Google Scholar] [CrossRef] [PubMed]
- Ondruš, M.; Sýkorová, V.; Bednárová, L.; Pohl, R.; Hocek, M. Enzymatic synthesis of hypermodified DNA polymers for sequence-specific display of four different hydrophobic groups. Nucleic. Acids Res. 2020, 48, 11982–11993. [Google Scholar] [CrossRef]
- Ollis, D.L.; Brick, P.; Hamlin, R.; Xuong, N.G.; Steitz, T.A. Structure of the large fragment of Escherichia coli DNA polymerase I complexed with TMP. Nature 1985, 313, 762–766. [Google Scholar] [CrossRef]
- Kim, Y.; Eom, S.H.; Wang, J.; Lee, D.S.; Suh, S.W.; Steitz, T.A. Crystal structure of Thermus aquaticus DNA polymerase. Nature 1995, 376, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Korolev, S.; Waksman, G. Crystal structures of open and closed forms of binary and ternary complexes of the large fragment of Thermus aquaticus DNA polymerase I: Structural basis for nucleotide incorporation. EMBO J. 1998, 17, 7514–7525. [Google Scholar] [CrossRef]
- Li, Y.; Mitaxov, V.; Waksman, G. Structure-based design of Taq DNA polymerases with improved properties of dideoxynucleotide incorporation. Proc. Natl. Acad. Sci. USA 1999, 96, 9491–9496. [Google Scholar] [CrossRef] [PubMed]
- Steitz, T.A. DNA Polymerases: Structural Diversity and Common Mechanisms. J Biol Chem. 1999, 274, 17395–17398. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, P.J.; Waksman, G. Structure and mechanism of DNA polymerases. In Advances in protein chemistry; Squire, J.M., Parry, D.A.D., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; Volume 71, pp. 401–440. [Google Scholar]
- Obeid, S.; Baccaroa, A.; Welteb, W.; Diederichsb, K.; Marx, A. Structural basis for the synthesis of nucleobase modified DNA by Thermus aquaticus DNA polymerase. Proc. Natl. Acad. Sci. USA 2010, 107, 21327–21331. [Google Scholar] [CrossRef] [PubMed]
- Bergen, K.; Steck, A.-L.; Strütt, S.; Baccaro, A.; Welte, W.; Diederichs, K.; Marx, A. Structures of KlenTaq DNA polymerase caught while incorporating C5-modified pyrimidine and C7-modified 7-deazapurine nucleoside triphosphates. J. Am. Chem. Soc. 2012, 134, 11840–11843. [Google Scholar] [CrossRef]
- Obeid, S.; Busskamp, H.; Welte, W.; Diederichs, K.; Marx, A. Interactions of non-polar and “Click-able” nucleotides in the confines of a DNA polymerase active site. Chem. Commun. 2012, 48, 8320–8322. [Google Scholar] [CrossRef]
- Hottin, A.; Marx, A. Structural Insights into the processing of nucleobase-modified nucleotides by DNA polymerases. Acc. Chem. Res. 2016, 49, 418–427. [Google Scholar] [CrossRef]
- Hottin, A.; Betz, K.; Diederichs, K.; Marx, A. Structural Basis for the KlenTaq DNA Polymerase Catalysed Incorporation of Alkene- versus Alkyne-Modified Nucleotides. Chemistry 2017, 23, 2109–2118. [Google Scholar] [CrossRef]
- Kropp, H.M.; Durr, S.L.; Peter, C.; Diederichs, K.; Marx, A. Snapshots of a modified nucleotide moving through the confines of a DNA polymerase. Proc. Natl. Acad. Sci. USA 2018, 115, 9992–9997. [Google Scholar] [CrossRef]
- Cahová, H.; Panattoni, A.; Kielkowski, P.; Jindřich Fanfrlík, J.; Hocek, M. 5-Substituted Pyrimidine and 7-Substituted 7-Deazapurine dNTPs as Substrates for DNA Polymerases in Competitive Primer Extension in the Presence of Natural dNTPs. ACS Chem. Biol. 2016, 11, 3165–3171. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, N.; Jemth, A.S.; Brown, A.; Crampton, N.; Dear, P.; Holliger, P. CyDNA: Synthesis and replication of highly Cy-dye substituted DNA by an evolved polymerase. J. Am. Chem. Soc. 2010, 132, 5096–5104. [Google Scholar] [CrossRef]
- Wynne, S.A.; Pinheiro, V.B.; Holliger, P.; Leslie, A.G. Structures of an apo and a binary complex of an evolved archeal B family DNA polymerase capable of synthesizing highly Cy-dye labelled DNA. PLoS ONE 2013, 8, e70892. [Google Scholar] [CrossRef] [PubMed]
- Ivancová, I.; Leone, D.L.; Hocek, M. Reactive modifications of DNA nucleobases for labelling, bioconjugations, and cross-linking. Curr. Opin. Chem. Biol. 2019, 52, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Kuba, M.; Kraus, T.; Pohl, R.; Hocek, M. Nucleotide-Bearing Benzylidene-Tetrahydroxanthylium Near-IR Fluorophore for Sensing DNA Replication, Secondary Structures and Interactions. Chemistry 2020, 26, 11950–11954. [Google Scholar] [CrossRef]
- Zasedateleva, O.A.; Vasiliskov, V.A.; Surzhikov, S.A.; Kuznetsova, V.E.; Shershov, V.E.; Guseinov, T.O.; Smirnov, I.P.; Yurasov, R.A.; Spitsyn, M.A.; Chudinov, A.V. dUTPs conjugated with zwitterionic Cy3 or Cy5 fluorophore analogues are effective substrates for DNA amplification and labelling by Taq polymerase. Nucleic. Acids. Res. 2018, 46, e73. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic. Acids Res. 2000, 28, 235–242. Available online: https://www.rcsb.org (accessed on 31 July 2023). [CrossRef]
- BIOVIA, Dassault Systèmes, Discovery Studio, 2020, San Diego: Dassault Systèmes. 2020. Available online: https://discover.3ds.com/discovery-studio-visualizer-download (accessed on 31 July 2023).
- Shershov, V.E.; Kuznetsova, V.E.; Lysov, Y.P.; Guseinov, T.O.; Barsky, V.E.; Spitsyn, M.A.; Zasedateleva, O.A.; Vasiliskov, V.A.; Surzhikov, S.A.; Zasedatelev, A.S.; et al. The effect of chromophore charge on the incorporation efficiency of fluorescence-labeled nucleotides catalyzed by Taq DNA polymerase in template synthesis. Biophysiks 2015, 60, 1013–1015. [Google Scholar]
- Aschenbrenner, J.; Marx, A. DNA polymerases and biotechnological applications. Curr. Opin. Biotechnol. 2017, 48, 187–195. [Google Scholar] [CrossRef]
- Kropp, H.M.; Diederichs, K.; Marx, A. The Structure of an Archaeal B-Family DNA Polymerase in Complex with a Chemically Modified Nucleotide. Angew Chem. Int. Ed. Engl. 2019, 58, 5457–5461. [Google Scholar] [CrossRef]
- Mader, O.; Reiner, K.; Egelhaaf, H.J.; Fischer, R.; Brock, R. Structure property analysis of pentamethine indocyanine dyes: Identification of a new dye for life science applications. Bioconjug. Chem. 2004, 15, 70–78. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zasedateleva, O.A.; Surzhikov, S.A.; Kuznetsova, V.E.; Shershov, V.E.; Barsky, V.E.; Zasedatelev, A.S.; Chudinov, A.V. Non-Covalent Interactions between dUTP C5-Substituents and DNA Polymerase Decrease PCR Efficiency. Int. J. Mol. Sci. 2023, 24, 13643. https://doi.org/10.3390/ijms241713643
Zasedateleva OA, Surzhikov SA, Kuznetsova VE, Shershov VE, Barsky VE, Zasedatelev AS, Chudinov AV. Non-Covalent Interactions between dUTP C5-Substituents and DNA Polymerase Decrease PCR Efficiency. International Journal of Molecular Sciences. 2023; 24(17):13643. https://doi.org/10.3390/ijms241713643
Chicago/Turabian StyleZasedateleva, Olga A., Sergey A. Surzhikov, Viktoriya E. Kuznetsova, Valeriy E. Shershov, Victor E. Barsky, Alexander S. Zasedatelev, and Alexander V. Chudinov. 2023. "Non-Covalent Interactions between dUTP C5-Substituents and DNA Polymerase Decrease PCR Efficiency" International Journal of Molecular Sciences 24, no. 17: 13643. https://doi.org/10.3390/ijms241713643