
Transcriptomic Response of Differentiating Porcine Myotubes to Thermal Stress and Donor Piglet Age

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
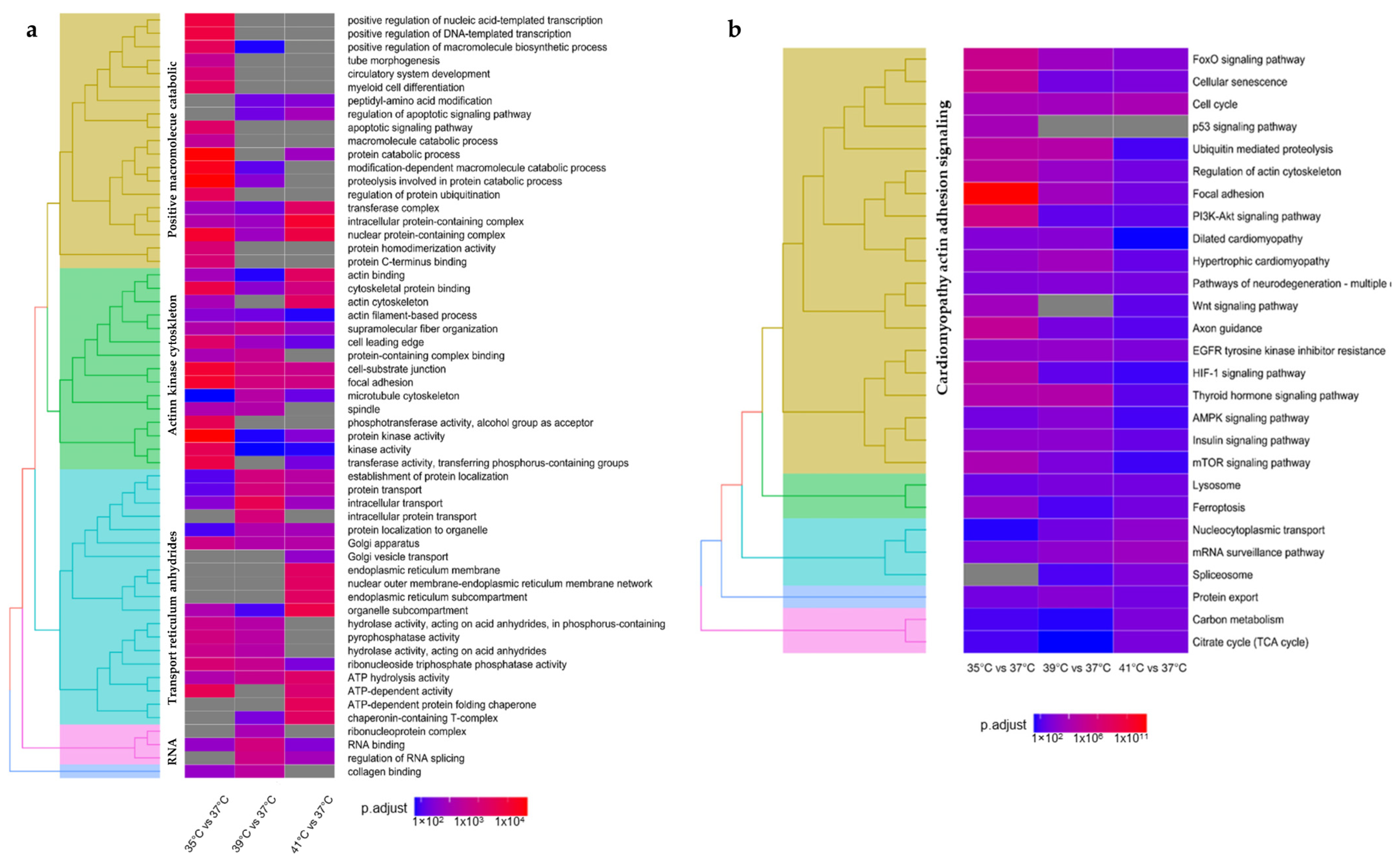
2.1. Transcriptome Analysis in Response to Culture Temperatures
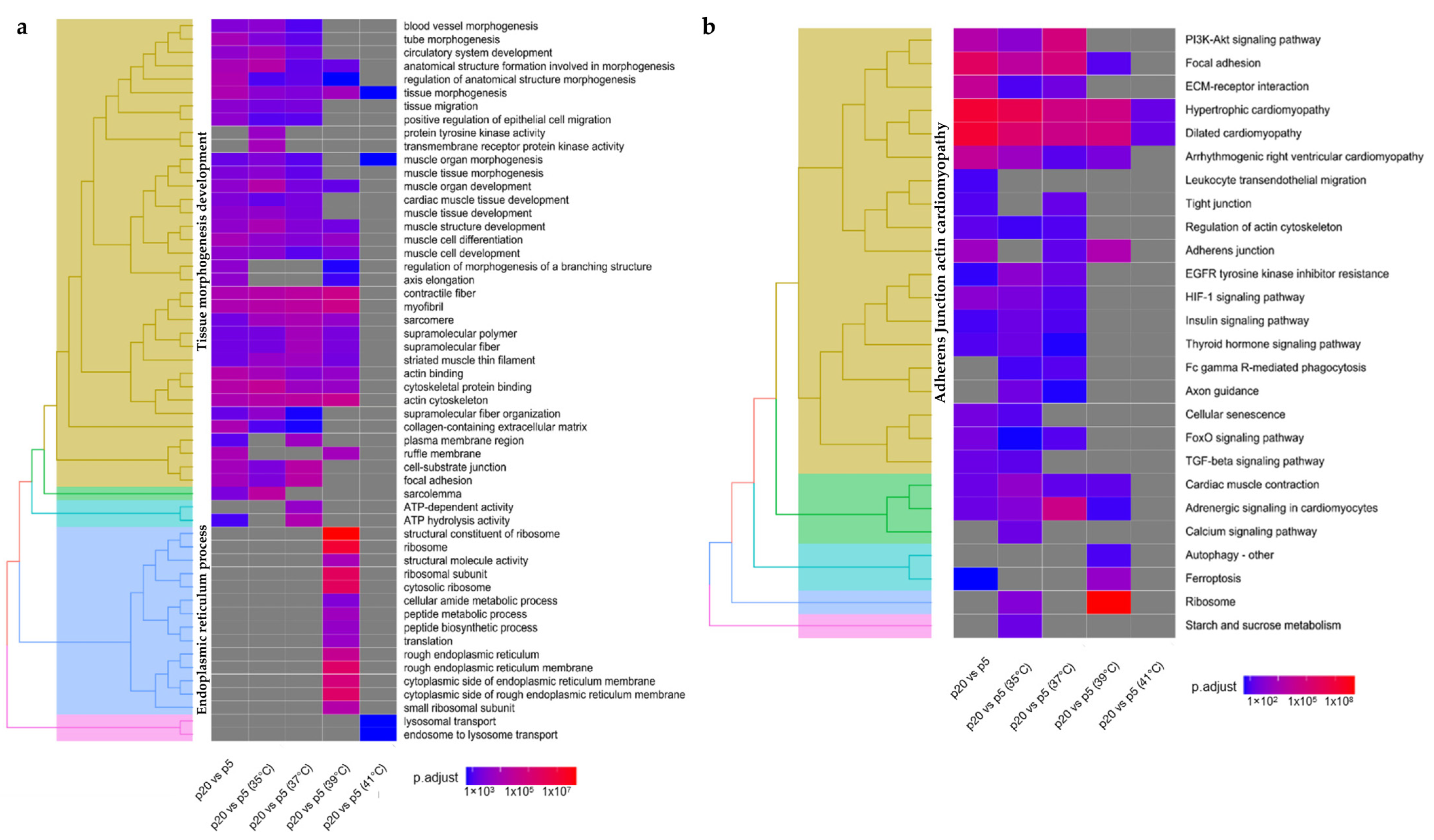
2.2. Transcriptome Analysis in Response to the Age of Donor Piglets to Culture Temperatures
2.3. GSH/GSSG Ratio in Response to Experimental Temperature Conditions
2.4. Metabolic Flux Assay
3. Discussion
3.1. Transcriptome Regulation in Response to Cold Stress
3.2. Transcriptome Regulation in Response to Heat Stress
3.3. Transcriptome Regulation in the Different Developmental Stages of Donor Piglets at Different Experimental Temperatures
3.4. Effects of Thermal Stress on Metabolic Flux and GSH/GSSG Ratio
4. Materials and Methods
4.1. Cell Culture
4.2. RNA Isolation and Microarray Analysis
4.3. Analysis of Differentially Expressed Genes (DEGs)
4.4. Functional Annotation of Gene Ontology (GO) and Analysis of Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment
4.5. Validation of Microarray Results
4.6. Mitochondrial Bioenergetics Assay and Ratio of Reduced/Oxidized Glutathione
4.7. Data Visualisation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thornton, P.; Nelson, G.; Mayberry, D.; Herrero, M. Impacts of Heat Stress on Global Cattle Production during the 21st Century: A Modelling Study. Lancet Planet Health 2022, 6, e192–e201. [Google Scholar] [CrossRef] [PubMed]
- D’Allaire, S.; Drolet, R.; Brodeur, D. Sow Mortality Associated with High Ambient Temperatures. Can. Vet. J. 1996, 37, 237–239. [Google Scholar] [PubMed]
- Pearce, S.C.; Mani, V.; Boddicker, R.L.; Johnson, J.S.; Weber, T.E.; Ross, J.W.; Baumgard, L.H.; Gabler, N.K. Heat Stress Reduces Barrier Function and Alters Intestinal Metabolism in Growing Pigs1. J. Anim. Sci. 2012, 90, 257–259. [Google Scholar] [CrossRef]
- Robbins, L.A.; Green-Miller, A.R.; Johnson, J.S.; Gaskill, B.N. Early Life Thermal Stress: Impacts on Future Temperature Preference in Weaned Pigs (3 to 15 Kg). J. Anim. Sci. 2020, 98, skaa327. [Google Scholar] [CrossRef]
- Yu, J.; Chen, S.; Zeng, Z.; Xing, S.; Chen, D.; Yu, B.; He, J.; Huang, Z.; Luo, Y.; Zheng, P.; et al. Effects of Cold Exposure on Performance and Skeletal Muscle Fiber in Weaned Piglets. Animals 2021, 11, 2148. [Google Scholar] [CrossRef]
- Blondin, D.P.; Haman, F. Shivering and Nonshivering Thermogenesis in Skeletal Muscles. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 156, pp. 153–173. [Google Scholar]
- Nowack, J.; Giroud, S.; Arnold, W.; Ruf, T. Muscle Non-Shivering Thermogenesis and Its Role in the Evolution of Endothermy. Front. Physiol. 2017, 8, 889. [Google Scholar] [CrossRef]
- Berthon, D.; Herpin, P.; Bertin, R.; De Marco, F.; le Dividich, J. Metabolic Changes Associated with Sustained 48-Hr Shivering Thermogenesis in the Newborn Pig. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1996, 114, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.-M.; Haman, F. Fuel Selection in Shivering Humans. Acta Physiol. Scand. 2005, 184, 319–329. [Google Scholar] [CrossRef]
- Lefaucheur, L.; Ecolan, P.; Lossec, G.; Gabillard, J.C.; Butler-Browne, G.S.; Herpin, P. Influence of Early Postnatal Cold Exposure on Myofiber Maturation in Pig Skeletal Muscle. J. Muscle Res. Cell Motil. 2001, 22, 439–452. [Google Scholar] [CrossRef]
- Langlois, S.; Cowan, K.N. Regulation of Skeletal Muscle Myoblast Differentiation and Proliferation by Pannexins. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2016; Volume 925, pp. 57–73. [Google Scholar]
- Asfour, H.A.; Allouh, M.Z.; Said, R.S. Myogenic Regulatory Factors: The Orchestrators of Myogenesis after 30 Years of Discovery. Exp. Biol. Med. 2018, 243, 118. [Google Scholar] [CrossRef]
- Hernández-Hernández, J.M.; García-González, E.G.; Brun, C.E.; Rudnicki, M.A. The Myogenic Regulatory Factors, Determinants of Muscle Development, Cell Identity and Regeneration. Semin. Cell Dev. Biol. 2017, 72, 10–18. [Google Scholar] [CrossRef]
- Metzger, K.; Kalbe, C.; Siengdee, P.; Ponsuksili, S. The Effects of Temperature and Donor Piglet Age on the Transcriptomic Profile and Energy Metabolism of Myoblasts. Front. Physiol. 2022, 13, 979283. [Google Scholar] [CrossRef] [PubMed]
- Metzger, K.; Tuchscherer, A.; Palin, M.F.; Ponsuksili, S.; Kalbe, C. Establishment and Validation of Cell Pools Using Primary Muscle Cells Derived from Satellite Cells of Pig Skeletal Muscle. In Vitro Cell Dev. Biol. Anim. 2020, 56, 193. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Valko, M. Thermodynamics of Free Radical Reactions and the Redox Environment of a Cell. In ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2011; Volume 1083, pp. 71–82. [Google Scholar]
- Zhu, P.; Oe, T.; Blair, I.A. Determination of Cellular Redox Status by Stable Isotope Dilution Liquid Chromatography/Mass Spectrometry Analysis of Glutathione and Glutathione Disulfide. Rapid Commun. Mass Spectrom. 2008, 22, 432–440. [Google Scholar] [CrossRef]
- Sajjanar, B.; Siengdee, P.; Trakooljul, N.; Liu, X.; Kalbe, C.; Wimmers, K.; Ponsuksili, S. Cross-Talk between Energy Metabolism and Epigenetics during Temperature Stress Response in C2C12 Myoblasts. Int. J. Hyperth. 2019, 36, 776–784. [Google Scholar] [CrossRef]
- Rauw, W.M.; de Mercado de la Peña, E.; Gomez-Raya, L.; García Cortés, L.A.; Ciruelos, J.J.; Gómez Izquierdo, E. Impact of Environmental Temperature on Production Traits in Pigs. Sci. Rep. 2020, 10, 2106. [Google Scholar] [CrossRef]
- Seelenbinder, K.M.; Zhao, L.D.; Hanigan, M.D.; Hulver, M.W.; McMillan, R.P.; Baumgard, L.H.; Selsby, J.T.; Ross, J.W.; Gabler, N.K.; Rhoads, R.P. Effects of Heat Stress during Porcine Reproductive and Respiratory Syndrome Virus Infection on Metabolic Responses in Growing Pigs. J. Anim. Sci. 2018, 96, 1375–1387. [Google Scholar] [CrossRef]
- Pant, M.; Bal, N.C.; Periasamy, M. Sarcolipin: A Key Thermogenic and Metabolic Regulator in Skeletal Muscle. Trends Endocrinol. Metab. 2016, 27, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Rehfeldt, C.; Lefaucheur, L.; Block, J.; Stabenow, B.; Pfuhl, R.; Otten, W.; Metges, C.C.; Kalbe, C. Limited and Excess Protein Intake of Pregnant Gilts Differently Affects Body Composition and Cellularity of Skeletal Muscle and Subcutaneous Adipose Tissue of Newborn and Weanling Piglets. Eur. J. Nutr. 2012, 51, 151–165. [Google Scholar] [CrossRef]
- Ren, D.; Song, J.; Liu, R.; Zeng, X.; Yan, X.; Zhang, Q.; Yuan, X. Molecular and Biomechanical Adaptations to Mechanical Stretch in Cultured Myotubes. Front. Physiol. 2021, 12, 689492. [Google Scholar] [CrossRef]
- Fujita, J. Cold Shock Response in Mammalian Cells JMMB Symposium. J. Mol. Microbiol. Biotechnol 1999, 1, 243–255. [Google Scholar] [PubMed]
- Sonna, L.A.; Fujita, J.; Gaffin, S.L.; Lilly, C.M. Invited Review: Effects of Heat and Cold Stress on Mammalian Gene Expression. J. Appl. Physiol. 2002, 92, 1725–1742. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, J.A.; Mertens, P.R. Cold Shock Proteins: From Cellular Mechanisms to Pathophysiology and Disease. Cell Commun. Signal. 2018, 16, 63. [Google Scholar] [CrossRef]
- Fuse, T.; Yamada, K.; Asai, K.; Kato, T.; Nakanishi, M. Heat Shock-Mediated Cell Cycle Arrest Is Accompanied by Induction of P21 CKI. Biochem. Biophys. Res. Commun. 1996, 225, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Al-Fageeh, M.B.; Smales, C.M. Control and Regulation of the Cellular Responses to Cold Shock: The Responses in Yeast and Mammalian Systems. Biochem. J. 2006, 397, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Liebscher, I.; Ackley, B.; Araç, D.; Ariestanti, D.M.; Aust, G.; Bae, B.; Bista, B.R.; Bridges, J.P.; Duman, J.G.; Engel, F.B.; et al. New Functions and Signaling Mechanisms for the Class of Adhesion G Protein-Coupled Receptors. Ann. N. Y. Acad. Sci. 2014, 1333, 43–64. [Google Scholar] [CrossRef] [PubMed]
- Risha, M.A.; Ali, A.; Siengdee, P.; Trakooljul, N.; Haack, F.; Dannenberger, D.; Wimmers, K.; Ponsuksili, S. Wnt Signaling Related Transcripts and Their Relationship to Energy Metabolism in C2C12 Myoblasts under Temperature Stress. PeerJ 2021, 9, e11625. [Google Scholar] [CrossRef]
- Cuella-Martin, R.; Oliveira, C.; Lockstone, H.E.; Snellenberg, S.; Grolmusova, N.; Chapman, J.R. 53BP1 Integrates DNA Repair and P53-Dependent Cell Fate Decisions via Distinct Mechanisms. Mol. Cell 2016, 64, 51–64. [Google Scholar] [CrossRef]
- Ohnishi, T.; Wang, X.; Ohnishi, K.; Takahashi, A. P53-Dependent Induction of WAF1 by Cold Shock in Human Glioblastoma Cells. Oncogene 1998, 16, 1507–1511. [Google Scholar] [CrossRef]
- Żwirowski, S.; Kłosowska, A.; Obuchowski, I.; Nillegoda, N.B.; Piróg, A.; Ziętkiewicz, S.; Bukau, B.; Mogk, A.; Liberek, K. Hsp70 Displaces Small Heat Shock Proteins from Aggregates to Initiate Protein Refolding. EMBO J. 2017, 36, 783–796. [Google Scholar] [CrossRef]
- Ávila-Gómez, P.; Vieites-Prado, A.; Dopico-López, A.; Bashir, S.; Fernández-Susavila, H.; Gubern, C.; Pérez-Mato, M.; Correa-Paz, C.; Iglesias-Rey, R.; Sobrino, T.; et al. Cold Stress Protein RBM3 Responds to Hypothermia and Is Associated with Good Stroke Outcome. Brain Commun. 2020, 2, fcaa078. [Google Scholar] [CrossRef]
- Martínez-Salas, E.; de Quinto, S.L.; Ramos, R.; Fernández-Miragall, O. IRES Elements: Features of the RNA Structure Contributing to Their Activity. Biochimie 2002, 84, 755–763. [Google Scholar] [CrossRef]
- Park, I.-H.; Chen, J. Mammalian Target of Rapamycin (MTOR) Signaling Is Required for a Late-Stage Fusion Process during Skeletal Myotube Maturation. J. Biol. Chem. 2005, 280, 32009–32017. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Kang, R.; Tang, D. Ferroptosis by Lipid Peroxidation: The Tip of the Iceberg? Front. Cell Dev. Biol. 2021, 9, 646890. [Google Scholar] [CrossRef]
- Hattori, K.; Ishikawa, H.; Sakauchi, C.; Takayanagi, S.; Naguro, I.; Ichijo, H. Cold Stress-induced Ferroptosis Involves the ASK1-p38 Pathway. EMBO Rep. 2017, 18, 2067–2078. [Google Scholar] [CrossRef]
- Nakamura, T.; Ogawa, M.; Kojima, K.; Takayanagi, S.; Ishihara, S.; Hattori, K.; Naguro, I.; Ichijo, H. The Mitochondrial Ca2+ Uptake Regulator, MICU1, Is Involved in Cold Stress-induced Ferroptosis. EMBO Rep. 2021, 22, e51532. [Google Scholar] [CrossRef] [PubMed]
- Gungor, B.; Gombos, I.; Crul, T.; Ayaydin, F.; Szabó, L.; Török, Z.; Mátés, L.; Vígh, L.; Horváth, I. Rac1 Participates in Thermally Induced Alterations of the Cytoskeleton, Cell Morphology and Lipid Rafts, and Regulates the Expression of Heat Shock Proteins in B16F10 Melanoma Cells. PLoS ONE 2014, 9, e89136. [Google Scholar] [CrossRef] [PubMed]
- van Hoof, A.; Wagner, E.J. A Brief Survey of MRNA Surveillance. Trends Biochem. Sci. 2011, 36, 585–592. [Google Scholar] [CrossRef]
- Tiago, T.; Hummel, B.; Morelli, F.F.; Basile, V.; Vinet, J.; Galli, V.; Mediani, L.; Antoniani, F.; Pomella, S.; Cassandri, M.; et al. Small Heat-Shock Protein HSPB3 Promotes Myogenesis by Regulating the Lamin B Receptor. Cell Death Dis. 2021, 12, 452. [Google Scholar] [CrossRef]
- Xu, L.; Lin, X.; Zheng, Y.; Zhou, H. Silencing of Heat Shock Protein 27 Increases the Radiosensitivity of Non-small Cell Lung Carcinoma Cells. Mol. Med. Rep. 2019, 20, 613–621. [Google Scholar] [CrossRef]
- Kusuluri, D.K.; Güler, B.E.; Knapp, B.; Horn, N.; Boldt, K.; Ueffing, M.; Aust, G.; Wolfrum, U. Adhesion G Protein-Coupled Receptor VLGR1/ADGRV1 Regulates Cell Spreading and Migration by Mechanosensing at Focal Adhesions. iScience 2021, 24, 102283. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Feng, Z.; Levine, A.J. The Regulation of Multiple P53 Stress Responses Is Mediated through MDM2. Genes Cancer 2012, 3, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Muns, R.; Nuntapaitoon, M.; Tummaruk, P. Non-Infectious Causes of Pre-Weaning Mortality in Piglets. Livest. Sci. 2016, 184, 46–57. [Google Scholar] [CrossRef]
- Baxter, E.M.; Jarvis, S.; D’Eath, R.B.; Ross, D.W.; Robson, S.K.; Farish, M.; Nevison, I.M.; Lawrence, A.B.; Edwards, S.A. Investigating the Behavioural and Physiological Indicators of Neonatal Survival in Pigs. Theriogenology 2008, 69, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Maskal, J.M.; Duttlinger, A.W.; Kpodo, K.R.; McConn, B.R.; Byrd, C.J.; Richert, B.T.; Marchant-Forde, J.N.; Lay, D.C.; Perry, S.D.; Lucy, M.C.; et al. Evaluation and Mitigation of the Effects of in Utero Heat Stress on Piglet Growth Performance, Postabsorptive Metabolism, and Stress Response Following Weaning and Transport. J. Anim. Sci. 2020, 98, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Villanueva-García, D.; Mota-Rojas, D.; Martínez-Burnes, J.; Olmos-Hernández, A.; Mora-Medina, P.; Salmerón, C.; Gómez, J.; Boscato, L.; Gutiérrez-Pérez, O.; Cruz, V.; et al. Hypothermia in Newly Born Piglets: Mechanisms of Thermoregulation and Pathophysiology of Death. J. Anim. Behav. Biometeorol. 2021, 9, 2101. [Google Scholar] [CrossRef]
- He, T.; He, L.; Gao, E.; Hu, J.; Zang, J.; Wang, C.; Zhao, J.; Ma, X. Fat Deposition Deficiency Is Critical for the High Mortality of Pre-Weanling Newborn Piglets. J. Anim. Sci. Biotechnol. 2018, 9, 66. [Google Scholar] [CrossRef]
- Rothwell, N.J.; Stock, M.J. Insulin and Thermogenesis. Int. J. Obes. 1988, 12, 93–102. [Google Scholar] [PubMed]
- Hefco, E.; Krulich, L.; Illner, P.; Larsen, P.R. Effect of Acute Exposure to Cold on the Activity of the Hypothalamic-Pituitary-Thyroid System. Endocrinology 1975, 97, 1185–1195. [Google Scholar] [CrossRef]
- Tsibulnikov, S.; Maslov, L.; Voronkov, N.; Oeltgen, P. Thyroid Hormones and the Mechanisms of Adaptation to Cold. Hormones 2020, 19, 329–339. [Google Scholar] [CrossRef]
- Yarlagadda, A.; Clayton, A.H. Thermoregulation and the Role of Calcium Signalling in Neurotransmission. Psychiatry 2008, 5, 51–54. [Google Scholar]
- McCormick, J.J.; Dokladny, K.; Moseley, P.L.; Kenny, G.P. Autophagy and Heat: A Potential Role for Heat Therapy to Improve Autophagic Function in Health and Disease. J. Appl. Physiol. 2021, 130, 1–9. [Google Scholar] [CrossRef]
- Williams, C.M.; McCue, M.D.; Sunny, N.E.; Szejner-Sigal, A.; Morgan, T.J.; Allison, D.B.; Hahn, D.A. Cold Adaptation Increases Rates of Nutrient Flow and Metabolic Plasticity during Cold Exposure in Drosophila Melanogaster. Proc. R. Soc. B Biol. Sci. 2016, 283, 20161317. [Google Scholar] [CrossRef]
- Britton, S.; Alber, M.; Cannon, W.R. Enzyme Activities Predicted by Metabolite Concentrations and Solvent Capacity in the Cell. J. R. Soc. Interface 2020, 17, 20200656. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.; Mitchell, J.B.; McPherson, S. The Effects of Glutathione Depletion on Thermotolerance and Heat Stress Protein Synthesis. Br. J. Cancer 1984, 49, 753–758. [Google Scholar] [CrossRef]
- Mitchell, J.; Russo, A.; Kinsella, T.; Glatstein, E. Glutathione Elevation during Thermotolerance Induction and Thermosensitization by Glutathione Depletion. Cancer Res. 1983, 43, 987–991. [Google Scholar]
- Zhao, Q.L.; Fujiwara, Y.; Kondo, T. Mechanism of Cell Death Induction by Nitroxide and Hyperthermia. Free Radic. Biol. Med. 2006, 40, 1131–1143. [Google Scholar] [CrossRef]
- Shrieve, D.C.; Li, G.C.; Astromoff, A.; Harris, J.W. Cellular Glutathione, Thermal Sensitivity, and Thermotolerance in Chinese Hamster Fibroblasts and Their Heat-Resistant Variants. Cancer Res. 1986, 46, 1684–1687. [Google Scholar] [PubMed]
- Naito, H.; Powers, S.K.; Demirel, H.A.; Sugiura, T.; Dodd, S.L.; Aoki, J. Heat Stress Attenuates Skeletal Muscle Atrophy in Hindlimb-Unweighted Rats. J. Appl. Physiol. 2000, 88, 359–363. [Google Scholar] [CrossRef]
- Hu, H.; Dai, S.; Li, J.; Wen, A.; Bai, X. Glutamine Improves Heat Stress-Induced Oxidative Damage in the Broiler Thigh Muscle by Activating the Nuclear Factor Erythroid 2-Related 2/Kelch-like ECH-Associated Protein 1 Signaling Pathway. Poult. Sci. 2020, 99, 1454–1461. [Google Scholar] [CrossRef] [PubMed]
- Macho, A.; Hirsch, T.; Marzo, I.; Marchetti, P.; Dallaporta, B.; Susin, S.A.; Zamzami, N.; Kroemer, G. Glutathione Depletion Is an Early and Calcium Elevation Is a Late Event of Thymocyte Apoptosis. J. Immunol. 1997, 158, 4612–4619. [Google Scholar] [CrossRef]
- Armstrong, J.S.; Steinauer, K.K.; Hornung, B.; Irish, J.M.; Lecane, P.; Birrell, G.W.; Peehl, D.M.; Knox, S.J. Role of Glutathione Depletion and Reactive Oxygen Species Generation in Apoptotic Signaling in a Human B Lymphoma Cell Line. Cell Death Differ. 2002, 9, 252–263. [Google Scholar] [CrossRef]
- Slimen, I.B.; Najar, T.; Ghram, A.; Dabbebi, H.; Ben Mrad, M.; Abdrabbah, M. Reactive Oxygen Species, Heat Stress and Oxidative-Induced Mitochondrial Damage. A Review. Int. J. Hyperth. 2014, 30, 513–523. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First Line Defence Antioxidants-Superoxide Dismutase (SOD), Catalase (CAT) and Glutathione Peroxidase (GPX): Their Fundamental Role in the Entire Antioxidant Defence Grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Ardite, E.; Barbera, J.A.; Roca, J.; Fernández-Checa, J.C. Glutathione Depletion Impairs Myogenic Differentiation of Murine Skeletal Muscle C2C12 Cells through Sustained NF-ΚB Activation. Am. J. Pathol. 2004, 165, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Rajasekaran, N.S.; Shelar, S.B.; Jones, D.P.; Hoidal, J.R. Reductive Stress Impairs Myogenic Differentiation. Redox Biol. 2020, 34, 101492. [Google Scholar] [CrossRef] [PubMed]
- Marí, M.; Morales, A.; Colell, A.; García-Ruiz, C.; Fernández-Checa, J.C. Mitochondrial Glutathione, a Key Survival Antioxidant. Antioxid. Redox Signal. 2009, 11, 2685–2700. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. ClusterProfiler 4.0: A Universal Enrichment Tool for Interpreting Omics Data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Sarais, F.; Montero, R.; Ostermann, S.; Rebl, A.; Köllner, B.; Goldammer, T. The Early Immune Response of Lymphoid and Myeloid Head-Kidney Cells of Rainbow Trout (Oncorhynchus mykiss) Stimulated with Aeromonas Salmonicida. Fishes 2022, 7, 12. [Google Scholar] [CrossRef]
- Rebl, A.; Rebl, H.; Verleih, M.; Haupt, S.; Köbis, J.M.; Goldammer, T.; Seyfert, H.M. At Least Two Genes Encode Many Variants of Irak3 in Rainbow Trout, but Neither the Full-Length Factor nor Its Variants Interfere Directly with the TLR-Mediated Stimulation of Inflammation. Front. Immunol. 2019, 10, 2246. [Google Scholar] [CrossRef]
- Galili, T.; O’Callaghan, A.; Sidi, J.; Sievert, C. Heatmaply: An R Package for Creating Interactive Cluster Heatmaps for Online Publishing. Bioinformatics 2018, 34, 1600–1602. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Pant, G.; Bhavnasi, Y.K.; Blanchard, S.G.; Brouwer, C. Pathview Web: User Friendly Pathway Visualization and Data Integration. Nucleic Acids Res. 2017, 45, W501–W508. [Google Scholar] [CrossRef] [PubMed]
- Blighe, K.; Rana, S.; Lewis, M. EnhancedVolcano: Publication-Ready Volcano Plots with Enhanced Colouring and Labeling, Version 1.18.0. 2023. Available online: https://bioconductor.org/packages/release/bioc/html/EnhancedVolcano.html (accessed on 1 July 2023). [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING V11: Protein–Protein Association Networks with Increased Coverage, Supporting Functional Discovery in Genome-Wide Experimental Datasets. Nucleic Acids Res. 2019, 47, D607. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarais, F.; Metzger, K.; Hadlich, F.; Kalbe, C.; Ponsuksili, S. Transcriptomic Response of Differentiating Porcine Myotubes to Thermal Stress and Donor Piglet Age. Int. J. Mol. Sci. 2023, 24, 13599. https://doi.org/10.3390/ijms241713599
Sarais F, Metzger K, Hadlich F, Kalbe C, Ponsuksili S. Transcriptomic Response of Differentiating Porcine Myotubes to Thermal Stress and Donor Piglet Age. International Journal of Molecular Sciences. 2023; 24(17):13599. https://doi.org/10.3390/ijms241713599
Chicago/Turabian StyleSarais, Fabio, Katharina Metzger, Frieder Hadlich, Claudia Kalbe, and Siriluck Ponsuksili. 2023. "Transcriptomic Response of Differentiating Porcine Myotubes to Thermal Stress and Donor Piglet Age" International Journal of Molecular Sciences 24, no. 17: 13599. https://doi.org/10.3390/ijms241713599