
The Induction of Long-Term Potentiation by Medial Septum Activation under Urethane Anesthesia Can Alter Gene Expression in the Hippocampus
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Loss of Cholinergic Neurons in the Medial Septum Does Not Lead to Transcriptomic Changes in the Dorsal Hippocampus under Basal Conditions
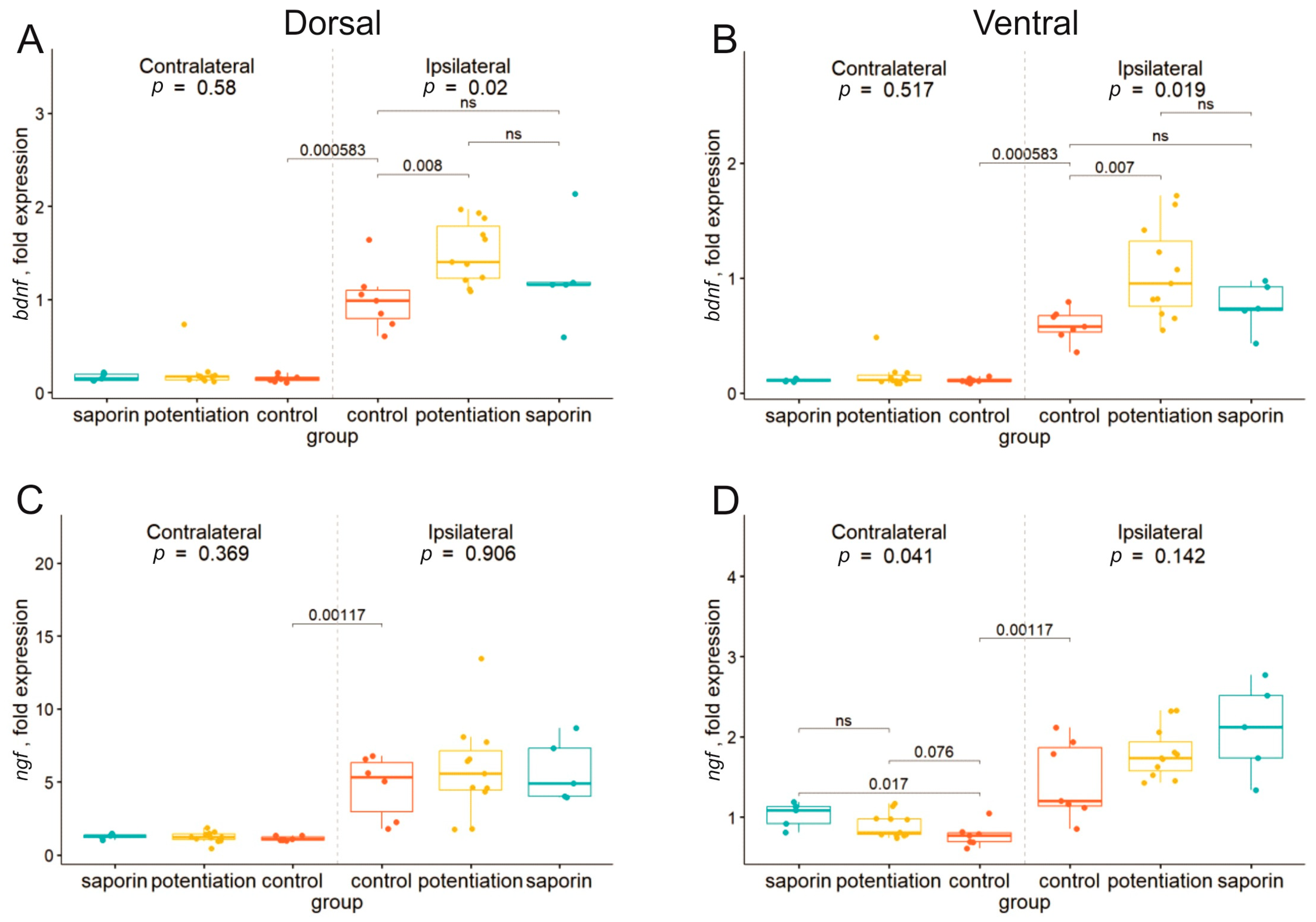
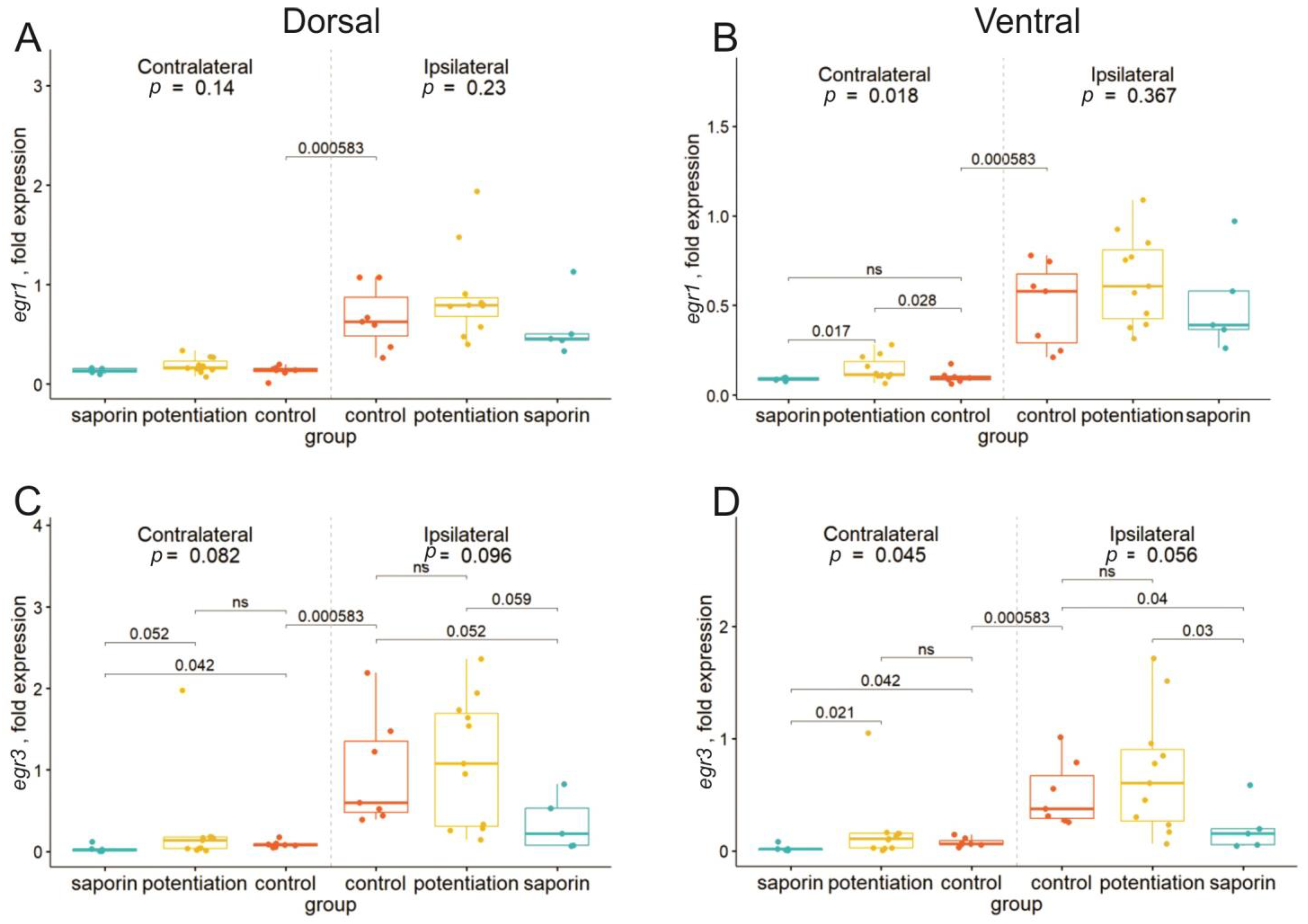
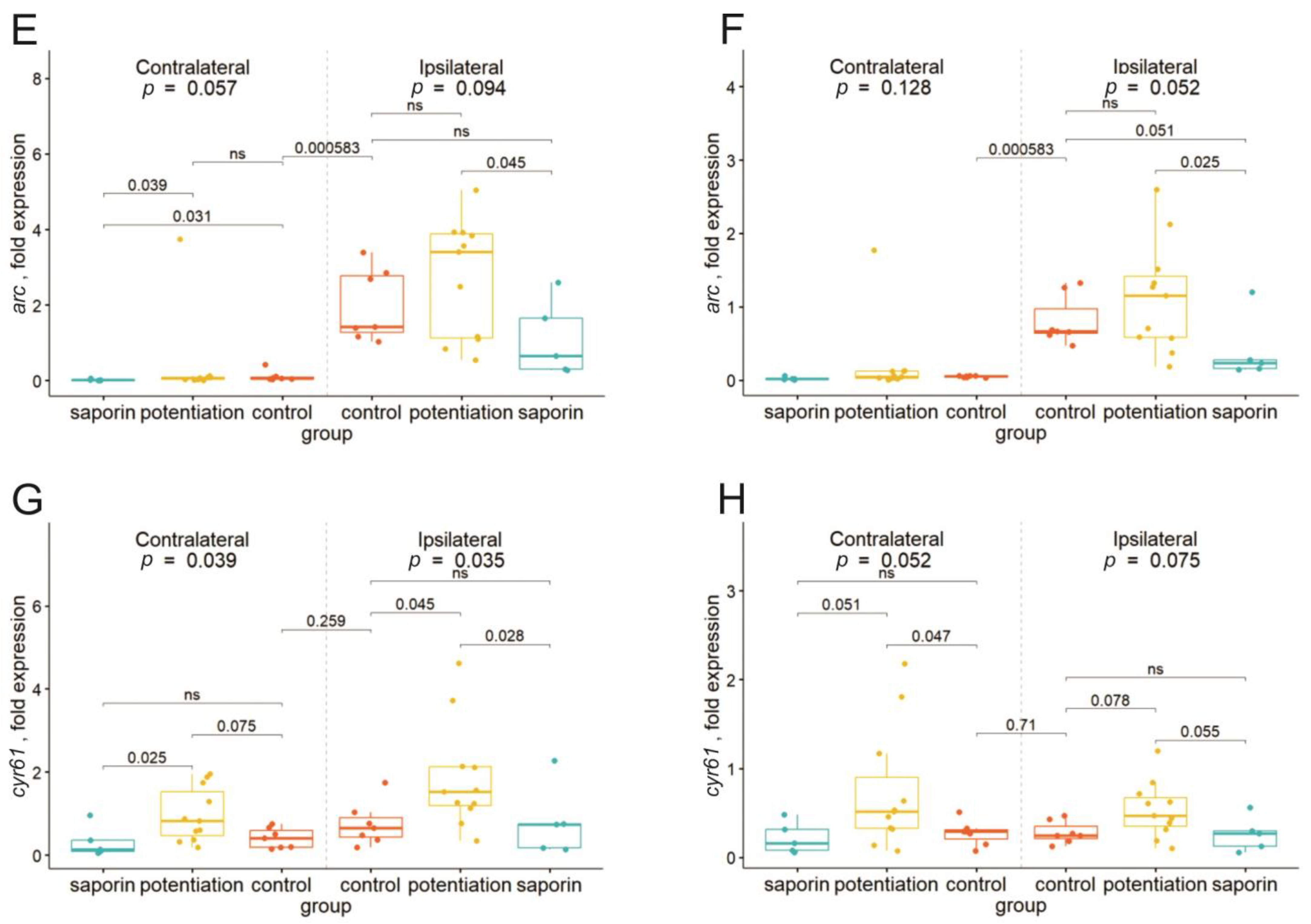
2.2. Degeneration of Cholinergic Neurons in the dMSA Prevents Expression of Some Early Genes after the Induction of Long-Term Potentiation
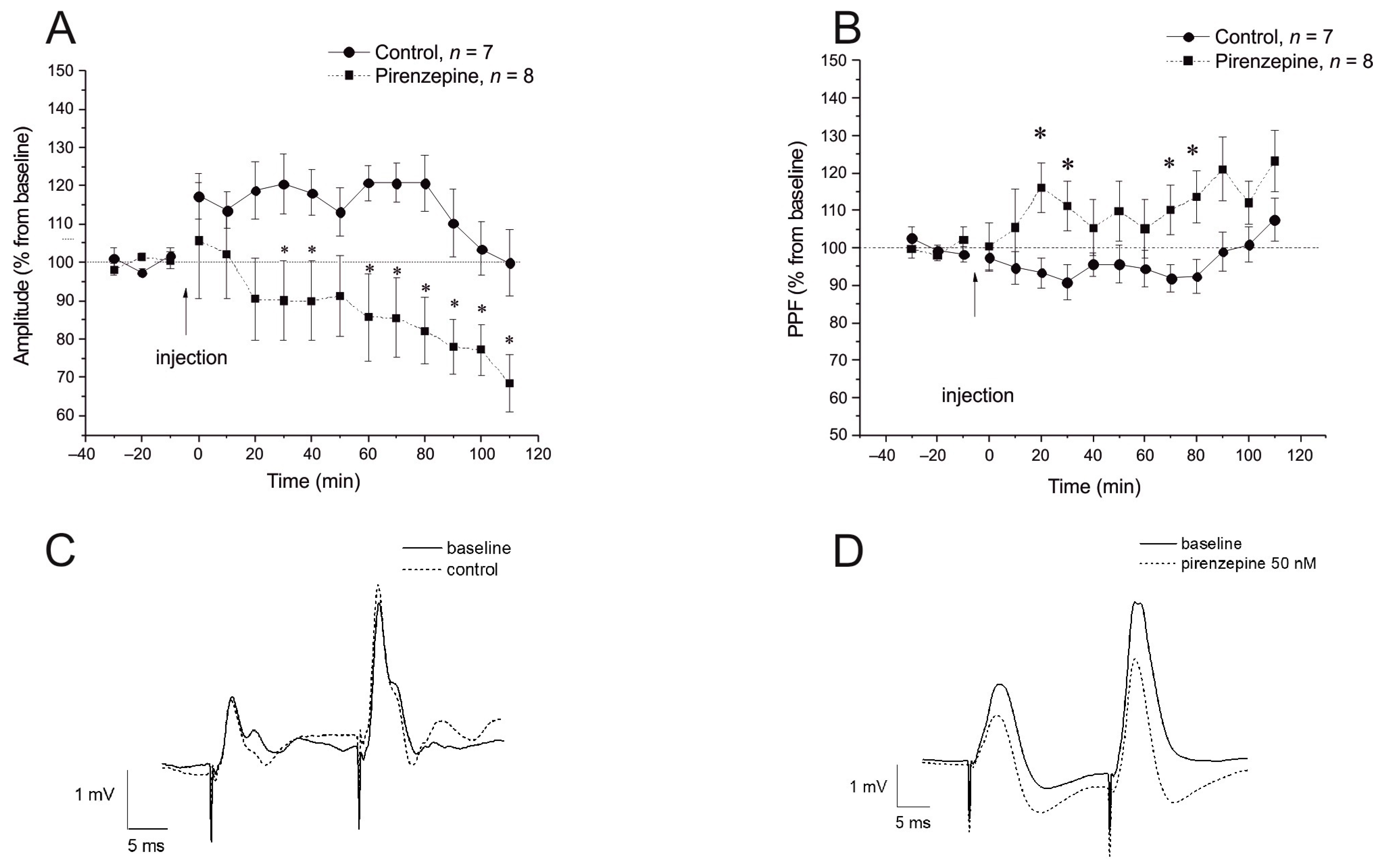
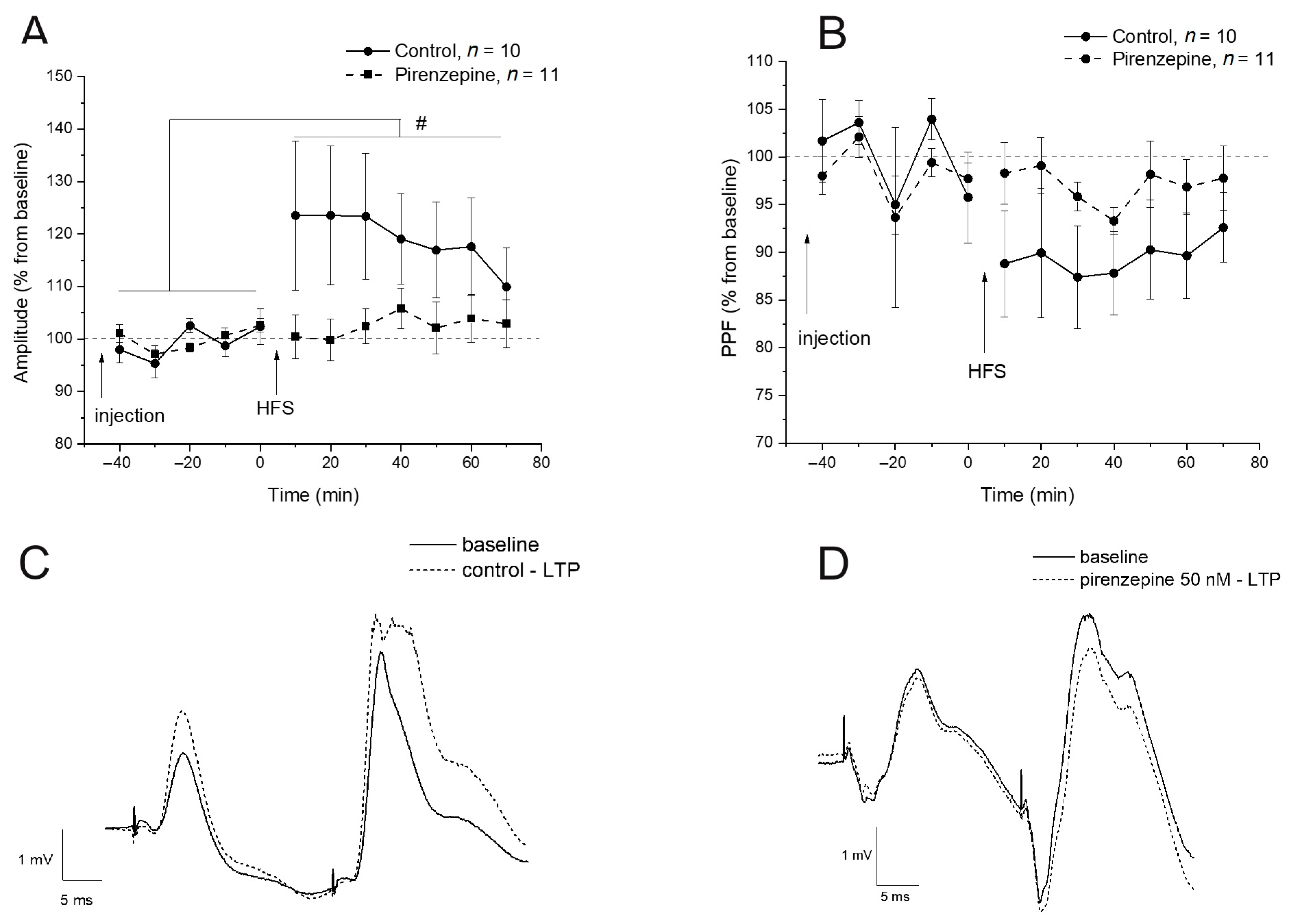
2.3. Inhibition of the M1 Muscarinic Receptors Alters the fEPSP Characteristics but Does Not Affect the Expression of Early Genes
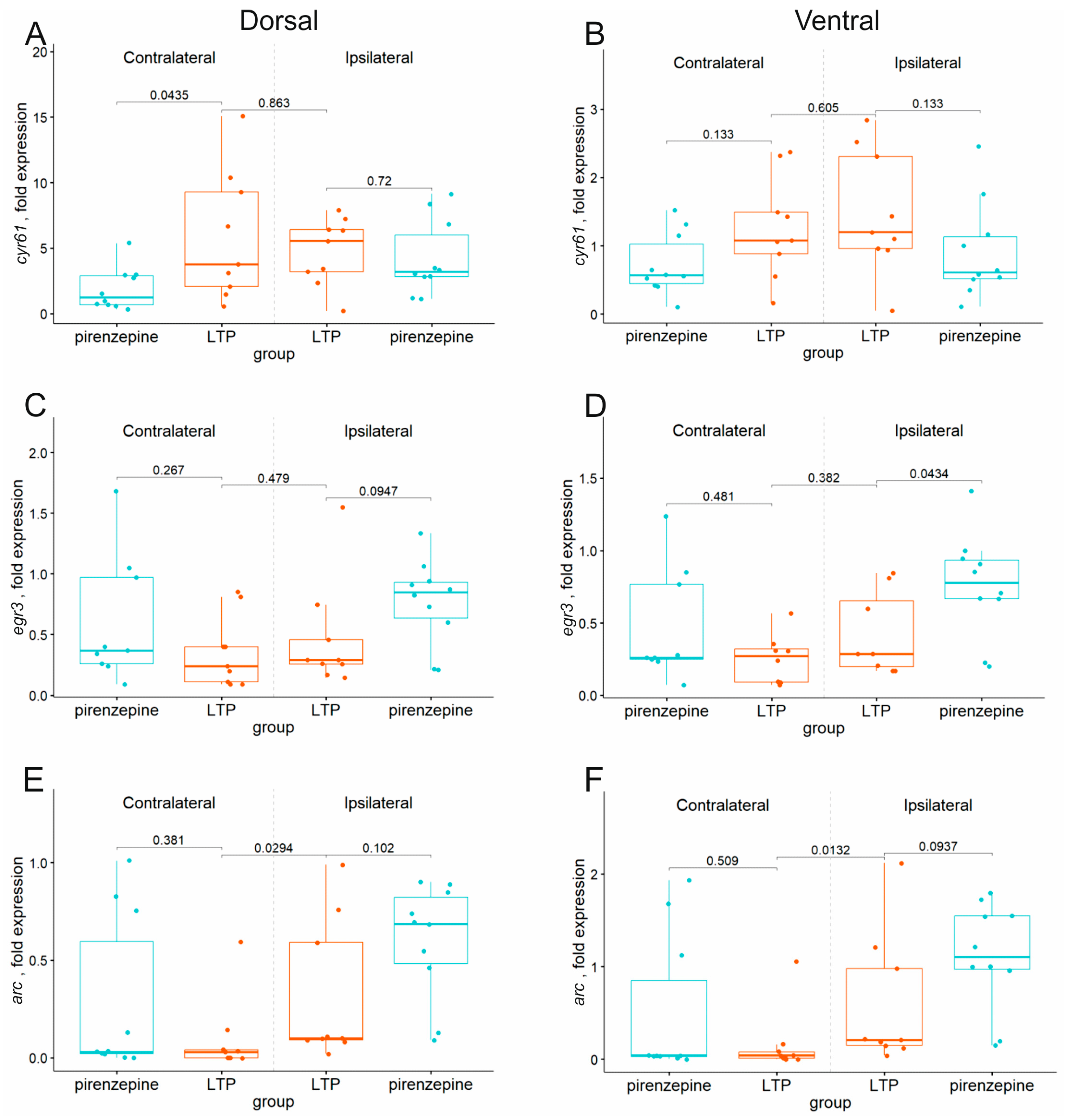
2.4. Inhibition of M1 Muscarinic Receptors Prevents LTP Induction and Induces Selective Suppression of the cyr61 Gene in the Dorsal Hippocampus
3. Discussion
4. Materials and Methods
4.1. 192IgG-Saporin Injection
4.2. RNAseq
4.3. RNA Isolation and Reverse Transcription
4.4. qPCR
4.5. Electrophysiology Recording and Drug Injection
4.6. Immunohistochemistry
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hampel, H.; Mesulam, M.-M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, A.S.; Vergallo, A.; Cavedo, E.; Snyder, P.J.; et al. The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain 2018, 141, 1917–1933. [Google Scholar] [CrossRef]
- Liu, A.K.L.; Lim, E.J.; Ahmed, I.; Chang, R.C.-C.; Pearce, R.K.B.; Gentleman, S.M. Review: Revisiting the human cholinergic nucleus of the diagonal band of Broca. Neuropathol. Appl. Neurobiol. 2018, 44, 647–662. [Google Scholar] [CrossRef] [PubMed]
- Janisiewicz, A.M.; Jackson, O.; Firoz, E.F.; Baxter, M.G. Environment-spatial conditional learning in rats with selective lesions of medial septal cholinergic neurons. Hippocampus 2004, 14, 265–273. [Google Scholar] [CrossRef]
- Paban, V.; Farioli, F.; Romier, B.; Chambon, C.; Alescio-Lautier, B. Gene expression profile in rat hippocampus with and without memory deficit. Neurobiol. Learn. Mem. 2010, 94, 42–56. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.U.; Lee, J.E.; Lee, S.E.; Chang, W.S.; Kim, S.J.; Chang, J.W. Improvements in memory after medial septum stimulation are associated with changes in hippocampal cholinergic activity and neurogenesis. Biomed Res. Int. 2014, 2014, 568587. [Google Scholar] [CrossRef]
- Knox, D.; Keller, S.M. Cholinergic neuronal lesions in the medial septum and vertical limb of the diagonal bands of Broca induce contextual fear memory generalization and impair acquisition of fear extinction. Hippocampus 2016, 26, 718–726. [Google Scholar] [CrossRef]
- Dobryakova, Y.V.; Kasianov, A.; Zaichenko, M.I.; Stepanichev, M.Y.; Chesnokova, E.A.; Kolosov, P.M.; Markevich, V.A.; Bolshakov, A.P. Intracerebroventricular Administration of 192IgG-Saporin Alters Expression of Microglia-Associated Genes in the Dorsal But Not Ventral Hippocampus. Front. Mol. Neurosci. 2018, 10, 429. [Google Scholar] [CrossRef]
- Auerbach, J.M.; Segal, M. Muscarinic receptors mediating depression and long-term potentiation in rat hippocampus. J. Physiol. 1996, 492 Pt 2, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Ovsepian, S.V.; Anwyl, R.; Rowan, M.J. Endogenous acetylcholine lowers the threshold for long-term potentiation induction in the CA1 area through muscarinic receptor activation: In vivo study. Eur. J. Neurosci. 2004, 20, 1267–1275. [Google Scholar] [CrossRef]
- Markevich, V.; Scorsa, A.M.; Dawe, G.S.; Stephenson, J.D. Cholinergic facilitation and inhibition of long-term potentiation of CA1 in the urethane-anaesthetized rats. Brain Res. 1997, 754, 95–102. [Google Scholar] [CrossRef]
- Dobryakova, Y.V.; Stepanichev, M.Y.; Markevich, V.A.; Bolshakov, A.P. Long-term potentiation in the hippocampal CA3 to CA1 synapses may be induced in vivo by activation of septal cholinergic inputs. Int. J. Neurosci. 2020, 132, 490–496. [Google Scholar] [CrossRef]
- Spivak, Y.S.; Karan, A.A.; Dobryakova, Y.V.; Medvedeva, T.M.; Markevich, V.A.; Bolshakov, A.P. Deep Brain Stimulation of the Medial Septal Area Can Modulate Gene Expression in the Hippocampus of Rats under Urethane Anesthesia. Int. J. Mol. Sci. 2022, 23, 6034. [Google Scholar] [CrossRef] [PubMed]
- Yap, E.L.; Greenberg, M.E. Activity-Regulated Transcription: Bridging the Gap between Neural Activity and Behavior. Neuron 2018, 100, 330–348. [Google Scholar] [CrossRef] [PubMed]
- Okuno, H. Regulation and function of immediate-early genes in the brain: Beyond neuronal activity markers. Neurosci. Res. 2011, 69, 175–186. [Google Scholar] [CrossRef]
- Volobueva, M.N.; Dobryakova, Y.V.; Manolova, A.O.; Stepanichev, M.Y.; Kvichansky, A.A.; Gulyaeva, N.V.; Markevich, V.A.; Bolshakov, A.P. Intracerebroventricular Administration of 192IgG-Saporin Alters the State of Microglia in the Neocortex. Neurochem. J. 2020, 14, 37–42. [Google Scholar] [CrossRef]
- Lindefors, N.; Ernfors, P.; Falkenberg, T.; Persson, H. Septal cholinergic afferents regulate expression of brain-derived neurotrophic factor and beta-nerve growth factor mRNA in rat hippocampus. Exp. Brain Res. 1992, 88, 78–90. [Google Scholar] [CrossRef]
- Von Der Kammer, H.; Albrecht, C.; Mayhaus, M.; Hoffmann, B.; Stanke, G.; Nitsch, R.M. Identification of genes regulated by muscarinic acetylcholine receptors: Application of an improved and statistically comprehensive mRNA differential display technique. Nucleic Acids Res. 1999, 27, 2211–2218. [Google Scholar] [CrossRef]
- Albrecht, C.; von der Kammer, H.; Mayhaus, M.; Klaudiny, J.; Schweitzer, M.; Nitsch, R.M. Muscarinic Acetylcholine Receptors Induce the Expression of the Immediate Early Growth Regulatory Gene CYR61. J. Biol. Chem. 2000, 275, 28929–28936. [Google Scholar] [CrossRef] [PubMed]
- von der Kammer, H.; Demiralay, C.; Andresen, B.; Albrecht, C.; Mayhaus, M.; Nitsch, R.M. Regulation of gene expression by muscarinic acetylcholine receptors. Biochem. Soc. Symp. 2001, 1, 131–140. [Google Scholar] [CrossRef]
- Luo, L.; Chen, W.H.; Wang, M.; Zhu, D.M.; She, J.Q.; Ruan, D.Y. Modulation of long-term potentiation by individual subtypes of muscarinic acetylcholine receptor in the rat dentate gyrus. Hippocampus 2008, 18, 989–995. [Google Scholar] [CrossRef]
- Buchanan, K.A.; Petrovic, M.M.; Chamberlain, S.E.L.; Marrion, N.V.; Mellor, J.R. Article Facilitation of Long-Term Potentiation by Muscarinic M 1 Receptors Is Mediated by Inhibition of SK Channels. Neuron 2010, 68, 948–963. [Google Scholar] [CrossRef]
- Burgard, E.C.; Sarvey, J.M. Muscarinic receptor activation facilitates the induction of long-term potentiation (LTP) in the rat dentate gyrus. Neurosci. Lett. 1990, 116, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Levey, A.I.; Edmunds, S.M.; Heilman, C.J.; Desmond, T.J.; Frey, K.A. Localization of muscarinic M3 receptor protein and M3 receptor binding in rat brain. Neuroscience 1994, 63, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Dannenberg, H.; Young, K.; Hasselmo, M. Modulation of Hippocampal Circuits by Muscarinic and Nicotinic Receptors. Front. Neural Circuits 2017, 11, 102. [Google Scholar] [CrossRef]
- French, P.J.; O’Connor, V.; Jones, M.W.; Davis, S.; Errington, M.L.; Voss, K.; Truchet, B.; Wotjak, C.; Stean, T.; Doyère, V.; et al. Subfield-specific immediate early gene expression associated with hippocampal long-term potentiation in vivo. Eur. J. Neurosci. 2001, 13, 968–976. [Google Scholar] [CrossRef]
- Worley, P.F.; Bhat, R.V.; Baraban, J.M.; Erickson, C.A.; McNaughton, B.L.; Barnes, C.A. Thresholds for synaptic activation of transcription factors in hippocampus: Correlation with long-term enhancement. J. Neurosci. 1993, 13, 4776–4786. [Google Scholar] [CrossRef]
- Dragunow, M.; Beilharz, E.; Mason, B.; Lawlor, P.; Abraham, W.; Gluckman, P. Brain-derived neurotrophic factor expression after long-term potentiation. Neurosci. Lett. 1993, 160, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Abraham, W.C.; Mason, S.E.; Demmer, J.; Williams, J.M.; Richardson, C.L.; Tate, W.P.; Lawlor, P.A.; Dragunow, M. Correlations between immediate early gene induction and the persistence of long-term potentiation. Neuroscience 1993, 56, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Dragunow, M.; Abraham, W.C.; Goulding, M.; Mason, S.E.; Robertson, H.A.; Faull, R.L.M. Long-term potentiation and the induction of c-fos mRNA and proteins in the dentate gyrus of unanesthetized rats. Neurosci. Lett. 1989, 101, 274–280. [Google Scholar] [CrossRef]
- Vila-Pueyo, M.; Cuenca-León, E.; Queirós, A.C.; Kulis, M.; Sintas, C.; Cormand, B.; Martín-Subero, J.I.; Pozo-Rosich, P.; Fernàndez-Castillo, N.; Macaya, A. Genome-wide DNA methylation analysis in an antimigraine-treated preclinical model of cortical spreading depolarization. Cephalalgia 2023, 43. [Google Scholar] [CrossRef]
- Cámara, M.S.; Martín Bujanda, M.; Mendioroz Iriarte, M. Epigenetic changes in headache. Neurologia 2021, 36. [Google Scholar] [CrossRef]
- Müller, C.; Remy, S. Septo–hippocampal interaction. Cell Tissue Res. 2018, 373, 565–575. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Nagy, A.J.; Barcsai, L.; Li, Q.; Ohsawa, M.; Mizuseki, K.; Berényi, A. The Medial Septum as a Potential Target for Treating Brain Disorders Associated with Oscillopathies. Front. Neural Circuits 2021, 15, 701080. [Google Scholar] [CrossRef] [PubMed]
- Santiago, L.J.; Abrol, R. Understanding G Protein Selectivity of Muscarinic Acetylcholine Receptors Using Computational Methods. Int. J. Mol. Sci. 2019, 20, 5290. [Google Scholar] [CrossRef]
- Aisaka, K.; Kihara, T.; Koike, M.; Kuroki, M.; Ishihara, T. Effect of yohimbine on urethane-induced hyperglycemia in rats. Jpn. J. Pharmacol. 1989, 49, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Carruba, M.O.; Bondiolotti, G.; Picotti, G.B.; Catteruccia, N.; Da Prada, M. Effects of diethyl ether, halothane, ketamine and urethane on sympathetic activity in the rat. Eur. J. Pharmacol. 1987, 134, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Faraguna, U.; Nelson, A.; Vyazovskiy, V.V.; Cirelli, C.; Tononi, G. Unilateral cortical spreading depression affects sleep need and induces molecular and electrophysiological signs of synaptic potentiation in vivo. Cereb. Cortex 2010, 20, 2939–2947. [Google Scholar] [CrossRef]
- Hughes, P.; Dragunow, M. Activation of pirenzepine-sensitive muscarinic receptors induces a specific pattern of immediate-early gene expression in rat brain neurons. Mol. Brain Res. 1994, 24, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Hughes, P.; Dragunow, M. Muscarinic receptor-mediated induction of Fos protein in rat brain. Neurosci. Lett. 1993, 150, 122–126. [Google Scholar] [CrossRef]
- Duclot, F.; Kabbaj, M. The Role of Early Growth Response 1 (EGR1) in Brain Plasticity and Neuropsychiatric Disorders. Front. Behav. Neurosci. 2017, 11, 35. [Google Scholar] [CrossRef]
- Draguhn, A.; Keller, M.; Reichinnek, S. Coordinated Network Activity in the Hippocampus. Front. Neurol. Neurosci. 2014, 34, 26–35. [Google Scholar] [PubMed]
- Wiley, R.G. Toxin-induced death of neurotrophin-sensitive neurons. Methods Mol. Biol. 2001, 169, 217–222. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Cunningham, F.; Allen, J.E.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Bennett, R.; et al. Ensembl 2022. Nucleic Acids Res. 2022, 50, D988–D995. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. The R package Rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. 2019, 47, e47. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Elsevier: Amsterdam, The Netherlands, 2007; ISBN 9780123741219. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dobryakova, Y.V.; Gerasimov, K.; Spivak, Y.S.; Korotkova, T.; Koryagina, A.; Deryabina, A.; Markevich, V.A.; Bolshakov, A.P. The Induction of Long-Term Potentiation by Medial Septum Activation under Urethane Anesthesia Can Alter Gene Expression in the Hippocampus. Int. J. Mol. Sci. 2023, 24, 12970. https://doi.org/10.3390/ijms241612970
Dobryakova YV, Gerasimov K, Spivak YS, Korotkova T, Koryagina A, Deryabina A, Markevich VA, Bolshakov AP. The Induction of Long-Term Potentiation by Medial Septum Activation under Urethane Anesthesia Can Alter Gene Expression in the Hippocampus. International Journal of Molecular Sciences. 2023; 24(16):12970. https://doi.org/10.3390/ijms241612970
Chicago/Turabian StyleDobryakova, Yulia V., Konstantin Gerasimov, Yulia S. Spivak, Tinna Korotkova, Alena Koryagina, Angelina Deryabina, Vladimir A. Markevich, and Alexey P. Bolshakov. 2023. "The Induction of Long-Term Potentiation by Medial Septum Activation under Urethane Anesthesia Can Alter Gene Expression in the Hippocampus" International Journal of Molecular Sciences 24, no. 16: 12970. https://doi.org/10.3390/ijms241612970