

Restraint Stress-Induced Immunosuppression Is Associated with Concurrent Macrophage Pyroptosis Cell Death in Mice

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
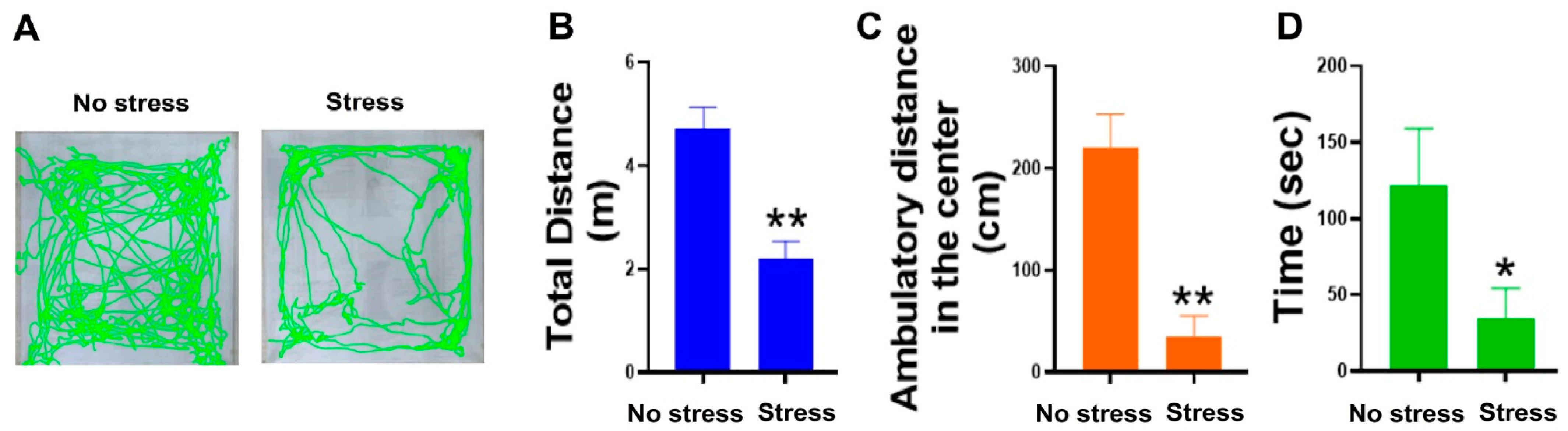
2.1. Restraint Stress Resulted in the Development of Anxiety-like Behaviors in C57Bl/6J Mice
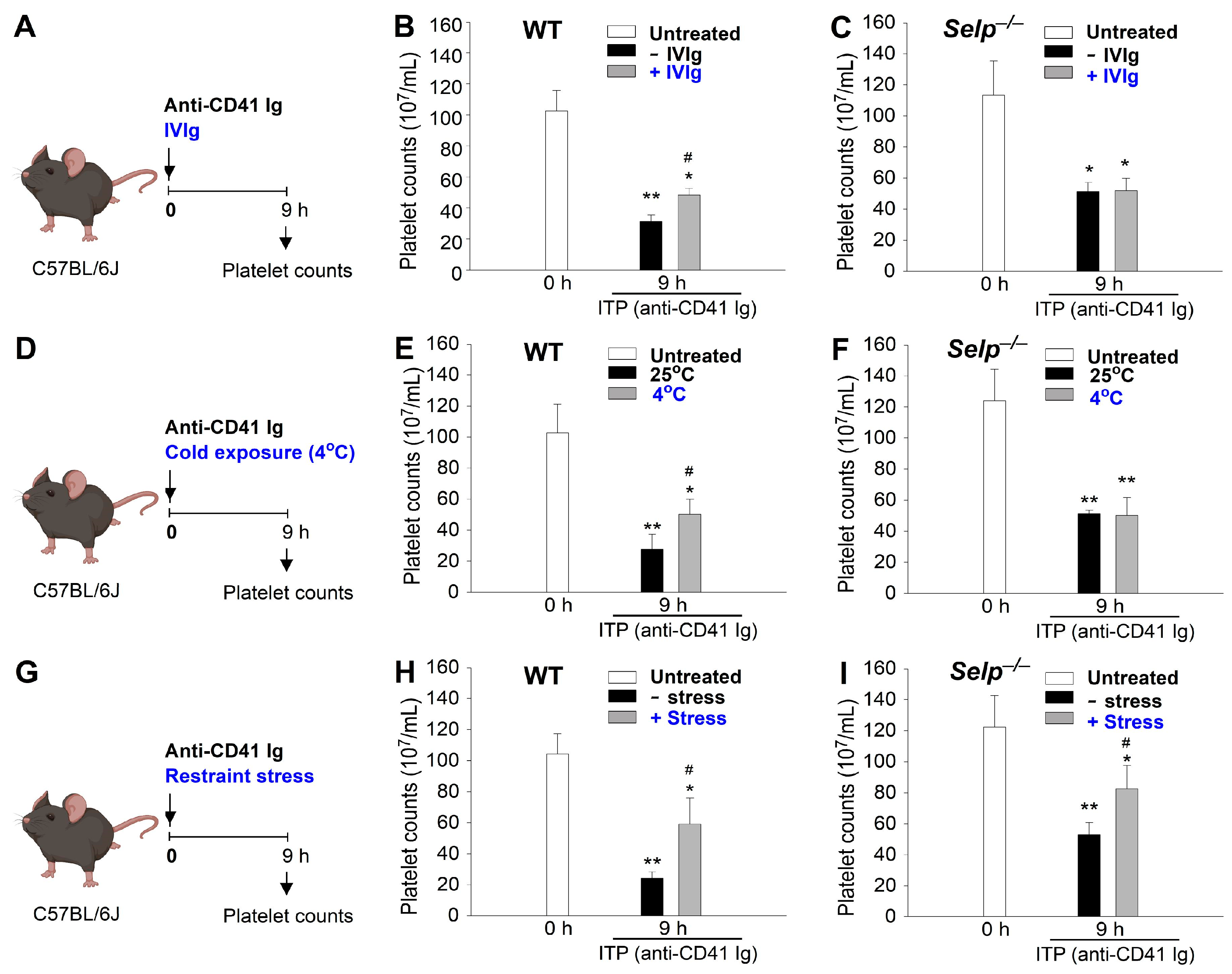
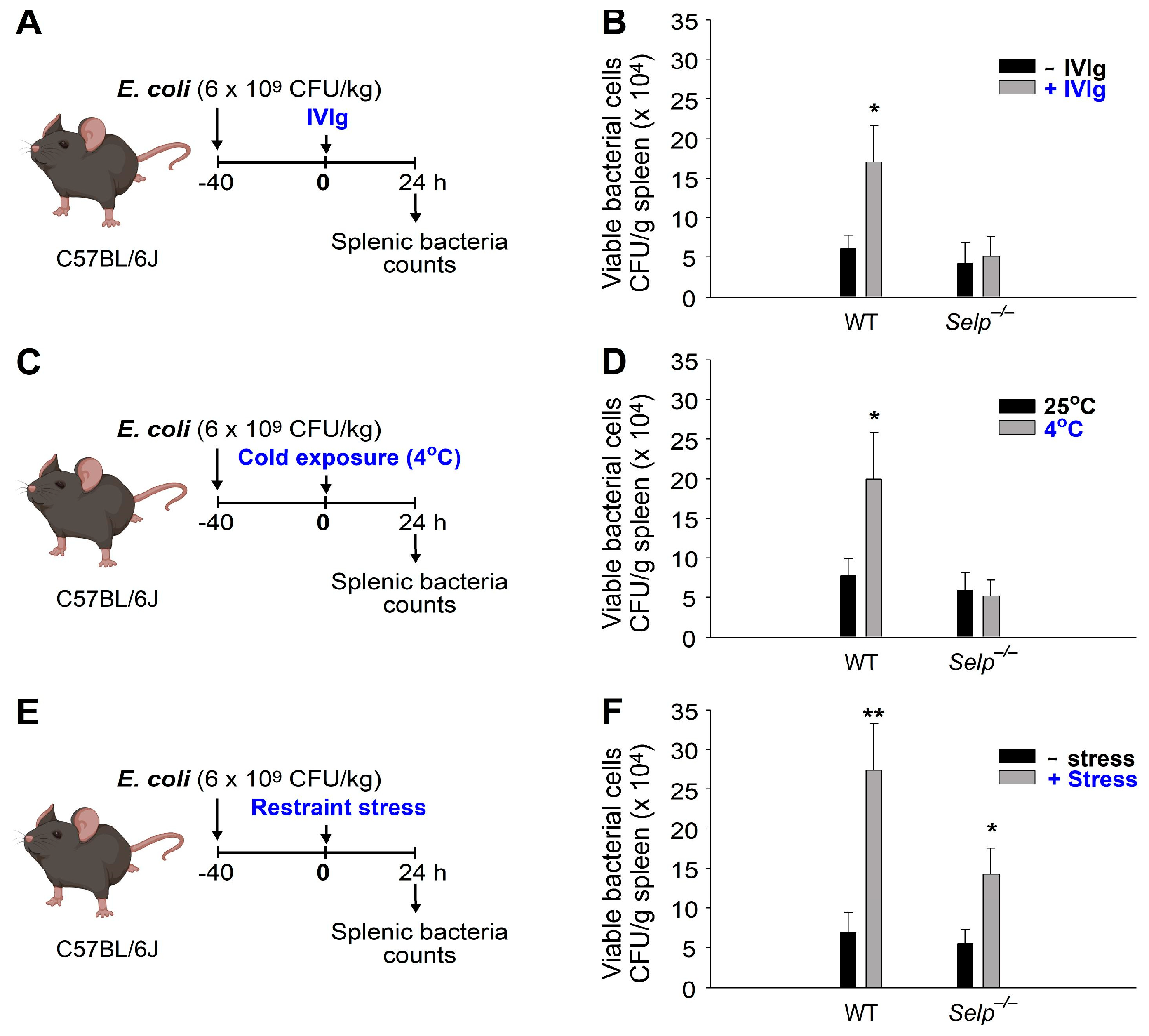
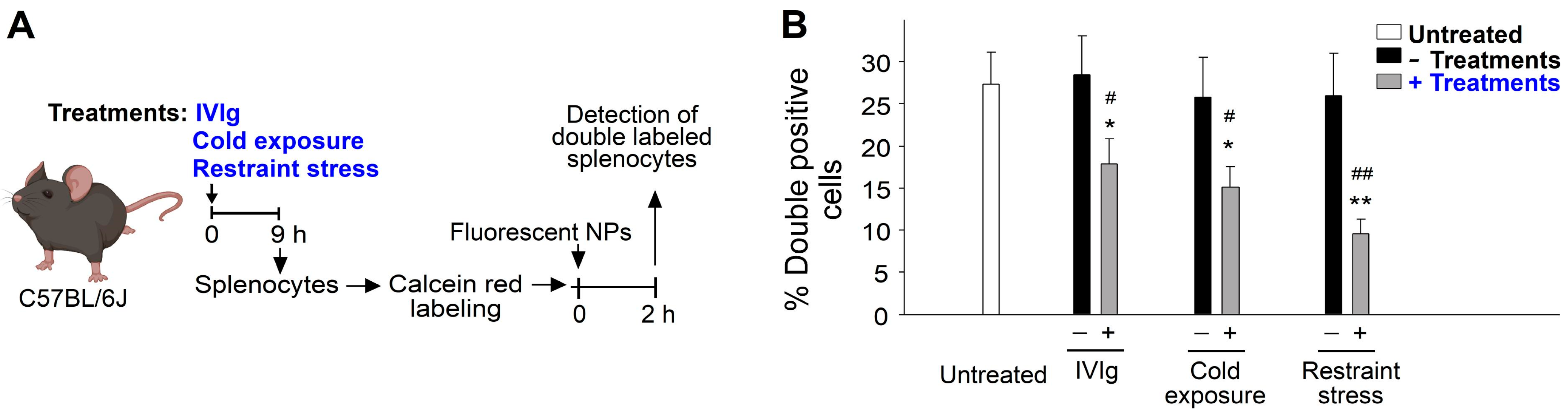
2.2. The Mechanism of Immunosuppression Differs between Restraint Stress, Intravenous Immunoglobulin (IVIg), and Ambient Cold Exposure in Mice
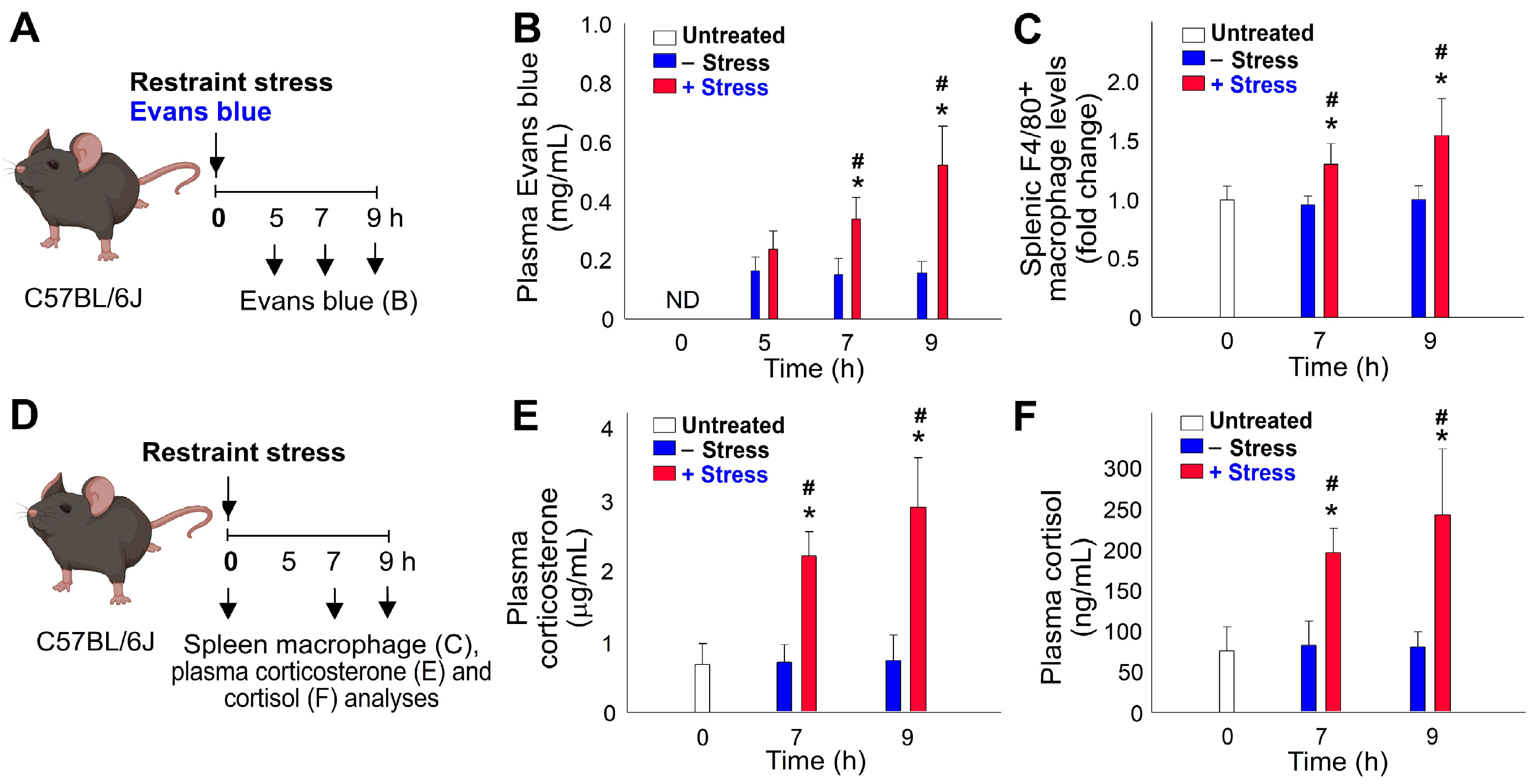
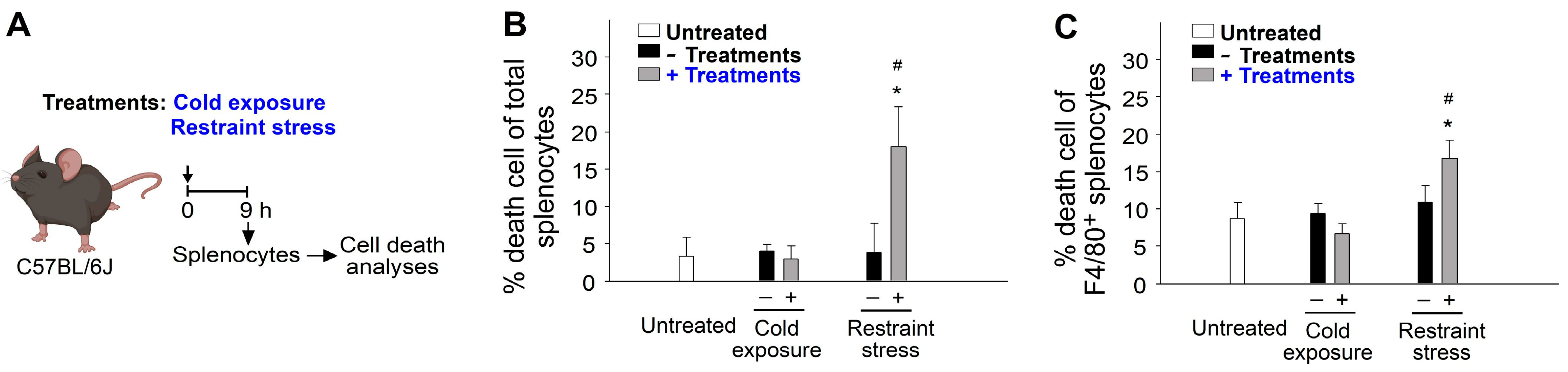
2.3. Restraint-Stress-Induced Immunosuppression Is Associated with Reduced Phagocyte Function and Increased Cell Death in Mice
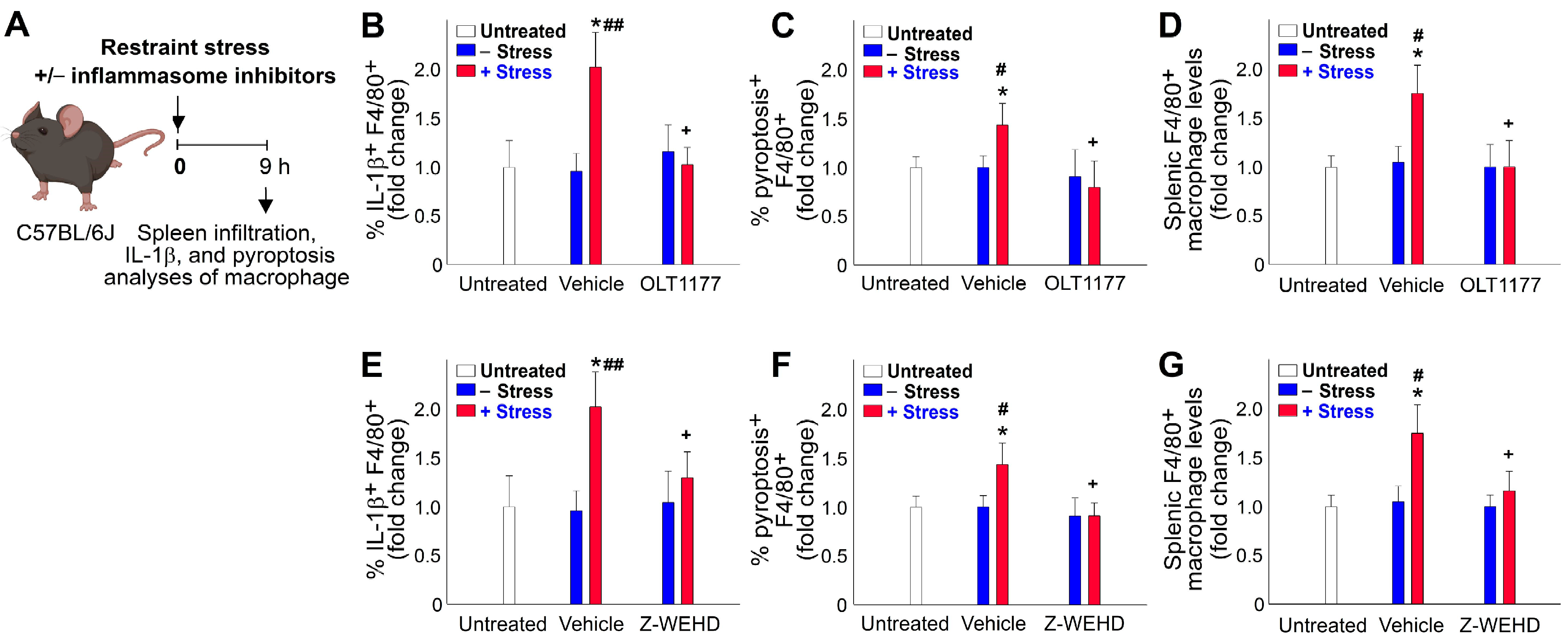
2.4. Restraint-Stress-Induced Pyroptosis of Spleen Macrophage
3. Discussion
4. Materials and Methods
4.1. Laboratory Mice
4.2. Open Field Test (OFT) Behavior Test
4.3. Restraint Stress and Restraint-Stress-Induced GI Leakage
4.4. Induction and Reversal of ITP
4.5. Analysis of Bacterial Clearance
4.6. Measurement of Phagocyte Functions and Cell Death Using Flow Cytometry
4.7. Measurements of Corticosterone and Cortisol
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Segerstrom, S.C.; Miller, G.E. Psychological stress and the human immune system: A meta-analytic study of 30 years of inquiry. Psychol. Bull. 2004, 130, 601–630. [Google Scholar] [CrossRef]
- Marshall, G.D., Jr. The adverse effects of psychological stress on immunoregulatory balance: Applications to human inflammatory diseases. Immunol. Allergy Clin. N. Am. 2011, 31, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Chatterjee, U.; MacDonald, C.R.; Repasky, E.A.; Halbreich, U. Psychosocial stress and immunosuppression in cancer: What can we learn from new research? BJPsych Adv. 2021, 27, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Bains, J.S.; Sharkey, K.A. Stress and immunity—The circuit makes the difference. Nat. Immunol. 2022, 23, 1137–1139. [Google Scholar] [CrossRef]
- Poller, W.C.; Downey, J.; Mooslechner, A.A.; Khan, N.; Li, L.; Chan, C.T.; McAlpine, C.S.; Xu, C.; Kahles, F.; He, S.; et al. Brain motor and fear circuits regulate leukocytes during acute stress. Nature 2022, 607, 578–584. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.C.; Fogaca, M.V.; Aguiar, D.C.; Guimaraes, F.S. Animal models of anxiety disorders and stress. Rev. Bras. Psiquiatr. 2013, 35 (Suppl. 2), S101–S111. [Google Scholar] [CrossRef]
- Pare, W.P.; Glavin, G.B. Restraint stress in biomedical research: A review. Neurosci. Biobehav. Rev. 1986, 10, 339–370. [Google Scholar] [CrossRef]
- Glavin, G.B.; Pare, W.P.; Sandbak, T.; Bakke, H.K.; Murison, R. Restraint stress in biomedical research: An update. Neurosci. Biobehav. Rev. 1994, 18, 223–249. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, S.; Liu, Y.; Ma, J.; Li, W.; Xu, X.; Wang, Y.; Luo, Y.; Cheng, K.; Zhuang, R. Acute Cold Water-Immersion Restraint Stress Induces Intestinal Injury and Reduces the Diversity of Gut Microbiota in Mice. Front. Cell. Infect. Microbiol. 2021, 11, 706849. [Google Scholar] [CrossRef]
- Rudak, P.T.; Choi, J.; Parkins, K.M.; Summers, K.L.; Jackson, D.N.; Foster, P.J.; Skaro, A.I.; Leslie, K.; McAlister, V.C.; Kuchroo, V.K.; et al. Chronic stress physically spares but functionally impairs innate-like invariant T cells. Cell Rep. 2021, 35, 108979. [Google Scholar] [CrossRef]
- Cao, M.; Huang, W.; Chen, Y.; Li, G.; Liu, N.; Wu, Y.; Wang, G.; Li, Q.; Kong, D.; Xue, T.; et al. Chronic restraint stress promotes the mobilization and recruitment of myeloid-derived suppressor cells through beta-adrenergic-activated CXCL5-CXCR2-Erk signaling cascades. Int. J. Cancer 2021, 149, 460–472. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Kishihara, K.; Wang, B.; Mizobe, K.; Kubo, C.; Nomoto, K. Restraint stress-induced immunosuppression by inhibiting leukocyte migration and Th1 cytokine expression during the intraperitoneal infection of Listeria monocytogenes. J. Neuroimmunol. 1998, 92, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.; Miao, J.; Hanley, G.; Stuart, C.; Sun, X.; Chen, T.; Yin, D. Chronic restraint stress promotes immune suppression through toll-like receptor 4-mediated phosphoinositide 3-kinase signaling. J. Neuroimmunol. 2008, 204, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Chuang, D.J.; Pethaperumal, S.; Siwakoti, B.; Chien, H.J.; Cheng, C.F.; Hung, S.C.; Lien, T.S.; Sun, D.S.; Chang, H.H. Activating Transcription Factor 3 Protects against Restraint Stress-Induced Gastrointestinal Injury in Mice. Cells 2021, 10, 3530. [Google Scholar] [CrossRef]
- Pethaperumal, S.; Hung, S.C.; Lien, T.S.; Sun, D.S.; Chang, H.H. P-Selectin is a Critical Factor for Platelet-Mediated Protection on Restraint Stress-Induced Gastrointestinal Injury in Mice. Int. J. Mol. Sci. 2022, 23, 1909. [Google Scholar] [CrossRef]
- Sun, D.S.; Lien, T.S.; Chang, H.H. Restraint stress-associated gastrointestinal injury and implications from the Evans-blue fed restraint stress mouse model. Tzu Chi Med. J. 2023, 35. in press. [Google Scholar]
- Siwakoti, B.; Lien, T.S.; Lin, Y.Y.; Pethaperumal, S.; Hung, S.C.; Sun, D.S.; Cheng, C.F.; Chang, H.H. The Role of Activating Transcription Factor 3 in Metformin’s Alleviation of Gastrointestinal Injury Induced by Restraint Stress in Mice. Int. J. Mol. Sci. 2023, 24, 10995. [Google Scholar] [CrossRef]
- Chan, H.; Huang, H.S.; Sun, D.S.; Lee, C.J.; Lien, T.S.; Chang, H.H. TRPM8 and RAAS-mediated hypertension is critical for cold-induced immunosuppression in mice. Oncotarget 2018, 9, 12781–12795. [Google Scholar] [CrossRef]
- Huang, H.S.; Sun, D.S.; Lien, T.S.; Chang, H.H. Dendritic cells modulate platelet activity in IVIg-mediated amelioration of ITP in mice. Blood 2010, 116, 5002–5009. [Google Scholar] [CrossRef]
- Zhang, R.L.; Lo, H.H.; Lei, C.; Ip, N.; Chen, J.; Law, B.Y. Current pharmacological intervention and development of targeting IVIG resistance in Kawasaki disease. Curr. Opin. Pharmacol. 2020, 54, 72–81. [Google Scholar] [CrossRef]
- Hansen, R.J.; Balthasar, J.P. Mechanisms of IVIG action in immune thrombocytopenic purpura. Clin. Lab. 2004, 50, 133–140. [Google Scholar]
- Cajamarca-Baron, J.; Buitrago-Bohorquez, J.; Mendoza Orozco, J.E.; Segura, O.; Guavita-Navarro, D.; Gallego-Cardona, L.; Cubides, H.; Arredondo, A.M.; Escobar, A.; Rojas-Villarraga, A. Efficacy and safety of intravenous immunoglobulin in patients with lupus nephritis: A systematic review of the literature. Autoimmun. Rev. 2022, 21, 103182. [Google Scholar] [CrossRef]
- Cornblath, D.R.; van Doorn, P.A.; Hartung, H.P.; Merkies, I.S.J.; Katzberg, H.D.; Hinterberger, D.; Clodi, E.; ProCID Investigators. Safety and Tolerability of Intravenous Immunoglobulin in Chronic Inflammatory Demyelinating Polyneuropathy: Results of the ProCID Study. Drug Saf. 2023, 46, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pasnoor, M.; Bril, V.; Levine, T.; Trivedi, J.; Silvestri, N.J.; Phadnis, M.; Katzberg, H.D.; Saperstein, D.S.; Wolfe, G.I.; Herbelin, L.; et al. Phase 2 trial in acetylcholine receptor antibody-positive myasthenia gravis of transition from intravenous to subcutaneous immunoglobulin: The MGSCIg study. Eur. J. Neurol. 2023, 30, 1417–1424. [Google Scholar] [CrossRef]
- Marcec, R.; Dodig, V.M.; Radanovic, I.; Likic, R. Intravenous immunoglobulin (IVIg) therapy in hospitalised adult COVID-19 patients: A systematic review and meta-analysis. Rev. Med. Virol. 2022, 32, e2397. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Y.; Lu, L.; Li, X.; Wu, Y.; Yang, Y.; Li, T.; Cao, W. Benefits of high-dose intravenous immunoglobulin on mortality in patients with severe COVID-19: An updated systematic review and meta-analysis. Front. Immunol. 2023, 14, 1116738. [Google Scholar] [CrossRef]
- Lai, C.C.; Chen, W.C.; Chen, C.Y.; Wei, Y.F. The effect of intravenous immunoglobulins on the outcomes of patients with COVID-19: A systematic review and meta-analysis of randomized controlled trials. Expert Rev. Anti-Infective Ther. 2022, 20, 1333–1340. [Google Scholar] [CrossRef] [PubMed]
- Kolahchi, Z.; Sohrabi, H.; Ekrami Nasab, S.; Jelodari Mamaghani, H.; Keyfari Alamdari, M.; Rezaei, N. Potential therapeutic approach of intravenous immunoglobulin against COVID-19. Allergy Asthma Clin. Immunol. 2021, 17, 105. [Google Scholar] [CrossRef]
- Flores-Oria, C.A.; Saturno, E.; Ramanathan, S.; Martinez Castillo, D.J.; Kumar, R.; Ferrer, N.; Mossaad, A.; Tellez, M.E.; Jon, C.; Waters, S.C.; et al. Intravenous immunoglobulin as adjuvant therapy for COVID-19: A case report and literature review. SAGE Open Med. Case Rep. 2021, 9, 2050313X211029699. [Google Scholar] [CrossRef]
- Manganotti, P.; Garascia, G.; Furlanis, G.; Buoite Stella, A. Efficacy of intravenous immunoglobulin (IVIg) on COVID-19-related neurological disorders over the last 2 years: An up-to-date narrative review. Front. Neurosci. 2023, 17, 1159929. [Google Scholar] [CrossRef]
- Bayry, J.; Ahmed, E.A.; Toscano-Rivero, D.; Vonniessen, N.; Genest, G.; Cohen, C.G.; Dembele, M.; Kaveri, S.V.; Mazer, B.D. Intravenous Immunoglobulin: Mechanism of Action in Autoimmune and Inflammatory Conditions. J. Allergy Clin. Immunol. Pract. 2023, 11, 1688–1697. [Google Scholar] [CrossRef] [PubMed]
- Seeling, M.; Pohnl, M.; Kara, S.; Horstmann, N.; Riemer, C.; Wohner, M.; Liang, C.; Bruckner, C.; Eiring, P.; Werner, A.; et al. Immunoglobulin G-dependent inhibition of inflammatory bone remodeling requires pattern recognition receptor Dectin-1. Immunity 2023, 56, 1046–1063.e1047. [Google Scholar] [CrossRef] [PubMed]
- Kindgen-Milles, D.; Feldt, T.; Jensen, B.E.O.; Dimski, T.; Brandenburger, T. Why the application of IVIG might be beneficial in patients with COVID-19. Lancet Respir. Med. 2022, 10, e15. [Google Scholar] [CrossRef]
- Tripathi, P. Immunosuppression in Patients with Diabetes Mellitus. In Immunosuppression and Immunomodulation; Tyagi, R.K., Sharma, P., Sharma, P., Eds.; IntechOpen: London, UK, 2022. [Google Scholar] [CrossRef]
- Berbudi, A.; Rahmadika, N.; Tjahjadi, A.I.; Ruslami, R. Type 2 Diabetes and its Impact on the Immune System. Curr. Diabetes Rev. 2020, 16, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Ferracini, M.; Martins, J.O.; Campos, M.R.; Anger, D.B.; Jancar, S. Impaired phagocytosis by alveolar macrophages from diabetic rats is related to the deficient coupling of LTs to the Fc gamma R signaling cascade. Mol. Immunol. 2010, 47, 1974–1980. [Google Scholar] [CrossRef]
- Pido-Lopez, J.; Andre, R.; Benjamin, A.C.; Ali, N.; Farag, S.; Tabrizi, S.J.; Bates, G.P. In vivo neutralization of the protagonist role of macrophages during the chronic inflammatory stage of Huntington’s disease. Sci. Rep. 2018, 8, 11447. [Google Scholar] [CrossRef]
- Hu, Z.; Zhan, J.; Pei, G.; Zeng, R. Depletion of macrophages with clodronate liposomes partially attenuates renal fibrosis on AKI-CKD transition. Ren. Fail. 2023, 45, 2149412. [Google Scholar] [CrossRef]
- Waltl, I.; Kaufer, C.; Broer, S.; Chhatbar, C.; Ghita, L.; Gerhauser, I.; Anjum, M.; Kalinke, U.; Loscher, W. Macrophage depletion by liposome-encapsulated clodronate suppresses seizures but not hippocampal damage after acute viral encephalitis. Neurobiol. Dis. 2018, 110, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Danenberg, H.D.; Fishbein, I.; Gao, J.; Monkkonen, J.; Reich, R.; Gati, I.; Moerman, E.; Golomb, G. Macrophage depletion by clodronate-containing liposomes reduces neointimal formation after balloon injury in rats and rabbits. Circulation 2002, 106, 599–605. [Google Scholar] [CrossRef]
- Lien, T.S.; Sun, D.S.; Wu, C.Y.; Chang, H.H. Exposure to Dengue Envelope Protein Domain III Induces Nlrp3 Inflammasome-Dependent Endothelial Dysfunction and Hemorrhage in Mice. Front. Immunol. 2021, 12, 617251. [Google Scholar] [CrossRef]
- Lien, T.S.; Sun, D.S.; Hung, S.C.; Wu, W.S.; Chang, H.H. Dengue Virus Envelope Protein Domain III Induces Nlrp3 Inflammasome-Dependent NETosis-Mediated Inflammation in Mice. Front. Immunol. 2021, 12, 618577. [Google Scholar] [CrossRef] [PubMed]
- Lien, T.S.; Chan, H.; Sun, D.S.; Wu, J.C.; Lin, Y.Y.; Lin, G.L.; Chang, H.H. Exposure of Platelets to Dengue Virus and Envelope Protein Domain III Induces Nlrp3 Inflammasome-Dependent Platelet Cell Death and Thrombocytopenia in Mice. Front. Immunol. 2021, 12, 616394. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Kang, R.; Berghe, T.V.; Vandenabeele, P.; Kroemer, G. The molecular machinery of regulated cell death. Cell Res. 2019, 29, 347–364. [Google Scholar] [CrossRef] [PubMed]
- Bertheloot, D.; Latz, E.; Franklin, B.S. Necroptosis, pyroptosis and apoptosis: An intricate game of cell death. Cell Mol. Immunol. 2021, 18, 1106–1121. [Google Scholar] [CrossRef]
- Jin, X.; Ma, Y.; Liu, D.; Huang, Y. Role of pyroptosis in the pathogenesis and treatment of diseases. MedComm 2023, 4, e249. [Google Scholar] [CrossRef]
- Hung, S.C.; Ke, L.C.; Lien, T.S.; Huang, H.S.; Sun, D.S.; Cheng, C.L.; Chang, H.H. Nanodiamond-Induced Thrombocytopenia in Mice Involve P-Selectin-Dependent Nlrp3 Inflammasome-Mediated Platelet Aggregation, Pyroptosis and Apoptosis. Front. Immunol. 2022, 13, 806686. [Google Scholar] [CrossRef]
- Blevins, H.M.; Xu, Y.; Biby, S.; Zhang, S. The NLRP3 Inflammasome Pathway: A Review of Mechanisms and Inhibitors for the Treatment of Inflammatory Diseases. Front. Aging Neurosci. 2022, 14, 879021. [Google Scholar] [CrossRef]
- La-Vu, M.; Tobias, B.C.; Schuette, P.J.; Adhikari, A. To Approach or Avoid: An Introductory Overview of the Study of Anxiety Using Rodent Assays. Front. Behav. Neurosci. 2020, 14, 145. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to measure locomotor and anxiety-like behavior in mice. J. Vis. Exp. 2015, 96, e52434. [Google Scholar] [CrossRef]
- Shoji, H.; Miyakawa, T. Differential effects of stress exposure via two types of restraint apparatuses on behavior and plasma corticosterone level in inbred male BALB/cAJcl mice. Neuropsychopharmacol. Rep. 2020, 40, 73–84. [Google Scholar] [CrossRef]
- Ma, M.; Chang, X.; Wu, H. Animal models of stress and stress-related neurocircuits: A comprehensive review. Stress Brain 2021, 1, 108–127. [Google Scholar] [CrossRef]
- Sanchez-Marin, L.; Flores-Lopez, M.; Gavito, A.L.; Suarez, J.; Pavon-Moron, F.J.; de Fonseca, F.R.; Serrano, A. Repeated Restraint Stress and Binge Alcohol during Adolescence Induce Long-Term Effects on Anxiety-like Behavior and the Expression of the Endocannabinoid System in Male Rats. Biomedicines 2022, 10, 593. [Google Scholar] [CrossRef] [PubMed]
- Bak, J.; Bobula, B.; Hess, G. Restraint Stress and Repeated Corticosterone Administration Differentially Affect Neuronal Excitability, Synaptic Transmission and 5-HT(7) Receptor Reactivity in the Dorsal Raphe Nucleus of Young Adult Male Rats. Int. J. Mol. Sci. 2022, 23, 4303. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.J.; Wang, Z.; Jin, S.W.; Liu, X.M.; Liu, Y.G.; Zhang, S.S.; Pan, R.L.; Jiang, N.; Liao, Y.H.; Yan, M.Z.; et al. Melatonin-related dysfunction in chronic restraint stress triggers sleep disorders in mice. Front. Pharmacol. 2023, 14, 1210393. [Google Scholar] [CrossRef]
- Xu, Y.X.; Liu, G.Y.; Ji, Z.Z.; Li, Y.Y.; Wang, Y.L.; Wu, X.Y.; Liu, J.L.; Ma, D.X.; Zhong, M.K.; Gao, C.B.; et al. Restraint stress induced anxiety and sleep in mice. Front. Psychiatry 2023, 14, 1090420. [Google Scholar] [CrossRef]
- Mao, Y.; Xu, Y.; Yuan, X. Validity of chronic restraint stress for modeling anhedonic-like behavior in rodents: A systematic review and meta-analysis. J. Int. Med. Res. 2022, 50, 3000605221075816. [Google Scholar] [CrossRef]
- Chaoui, N.; Anarghou, H.; Laaroussi, M.; Essaidi, O.; Najimi, M.; Chigr, F. Long lasting effect of acute restraint stress on behavior and brain anti-oxidative status. AIMS Neurosci. 2022, 9, 57–75. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Peng, Z.; Jiang, L.; Zhang, P.; Yang, P.; Yuan, Z.; Cheng, J. Dlg1 deletion in microglia ameliorates chronic restraint stress induced mice depression-like behavior. Front. Pharmacol. 2023, 14, 1124845. [Google Scholar] [CrossRef]
- Afridi, R.; Suk, K. Microglial Responses to Stress-Induced Depression: Causes and Consequences. Cells 2023, 12, 1521. [Google Scholar] [CrossRef]
- Lai, Z.K.; Yin, Y.Y.; Yan, J.Z.; Wei, Q.Q.; Wang, B.; Li, Y.F.; Zhang, L.M.; Wang, Y.L. Inulin-type oligosaccharides of Morinda officinalis exerted antidepressant effects by reducing hippocampal inflammation. Metab. Brain Dis. 2023, 38, 2065–2075. [Google Scholar] [CrossRef]
- Jiang, X.; Yi, S.; Liu, Q.; Su, D.; Li, L.; Xiao, C.; Zhang, J. Asperosaponin VI ameliorates the CMS-induced depressive-like behaviors by inducing a neuroprotective microglial phenotype in hippocampus via PPAR-gamma pathway. J. Neuroinflammation 2022, 19, 115. [Google Scholar] [CrossRef]
- Haem, S.C. A single 1 g/kg dose of intravenous immunoglobulin is a safe and effective treatment for immune thrombocytopenia; results of the first HaemSTAR ‘Flash-Mob’ retrospective study incorporating 961 patients. Br. J. Haematol. 2022, 196, 433–437. [Google Scholar] [CrossRef]
- Song, F.; Al-Samkari, H. Management of Adult Patients with Immune Thrombocytopenia (ITP): A Review on Current Guidance and Experience from Clinical Practice. J. Blood Med. 2021, 12, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Norris, P.A.A.; Segel, G.B.; Burack, W.R.; Sachs, U.J.; Lissenberg-Thunnissen, S.N.; Vidarsson, G.; Bayat, B.; Cserti-Gazdewich, C.M.; Callum, J.; Lin, Y.; et al. FcgammaRI and FcgammaRIII on splenic macrophages mediate phagocytosis of anti-glycoprotein IIb/IIIa autoantibody-opsonized platelets in immune thrombocytopenia. Haematologica 2021, 106, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Swinkels, M.; Rijkers, M.; Voorberg, J.; Vidarsson, G.; Leebeek, F.W.G.; Jansen, A.J.G. Emerging Concepts in Immune Thrombocytopenia. Front. Immunol. 2018, 9, 880. [Google Scholar] [CrossRef] [PubMed]
- Kuwana, M.; Okazaki, Y.; Ikeda, Y. Splenic macrophages maintain the anti-platelet autoimmune response via uptake of opsonized platelets in patients with immune thrombocytopenic purpura. J. Thromb. Haemost. 2009, 7, 322–329. [Google Scholar] [CrossRef]
- Hirayama, D.; Iida, T.; Nakase, H. The Phagocytic Function of Macrophage-Enforcing Innate Immunity and Tissue Homeostasis. Int. J. Mol. Sci. 2017, 19, 92. [Google Scholar] [CrossRef]
- Pidwill, G.R.; Gibson, J.F.; Cole, J.; Renshaw, S.A.; Foster, S.J. The Role of Macrophages in Staphylococcus aureus Infection. Front. Immunol. 2020, 11, 620339. [Google Scholar] [CrossRef]
- Jaganathan, D.; Bruscia, E.M.; Kopp, B.T. Emerging Concepts in Defective Macrophage Phagocytosis in Cystic Fibrosis. Int. J. Mol. Sci. 2022, 23, 7750. [Google Scholar] [CrossRef]
- Kau, J.H.; Sun, D.S.; Huang, H.S.; Lien, T.S.; Huang, H.H.; Lin, H.C.; Chang, H.H. Sublethal doses of anthrax lethal toxin on the suppression of macrophage phagocytosis. PLoS ONE 2010, 5, e14289. [Google Scholar] [CrossRef]
- Chauhan, D.; Vande Walle, L.; Lamkanfi, M. Therapeutic modulation of inflammasome pathways. Immunol. Rev. 2020, 297, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Pachathundikandi, S.K.; Blaser, N.; Bruns, H.; Backert, S. Helicobacter pylori Avoids the Critical Activation of NLRP3 Inflammasome-Mediated Production of Oncogenic Mature IL-1beta in Human Immune Cells. Cancers 2020, 12, 803. [Google Scholar] [CrossRef] [PubMed]
- Ershaid, N.; Sharon, Y.; Doron, H.; Raz, Y.; Shani, O.; Cohen, N.; Monteran, L.; Leider-Trejo, L.; Ben-Shmuel, A.; Yassin, M.; et al. NLRP3 inflammasome in fibroblasts links tissue damage with inflammation in breast cancer progression and metastasis. Nat. Commun. 2019, 10, 4375. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Woodruff, M.; Zhang, Y.; Miao, J.; Hanley, G.; Stuart, C.; Zeng, X.; Prabhakar, S.; Moorman, J.; Zhao, B.; et al. Toll-like receptor 4 mediates chronic restraint stress-induced immune suppression. J. Neuroimmunol. 2008, 194, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.Q.; Hong, X.D.; Chen, T.S.; Luo, P.F.; Xiao, S.C. Effects of caspase-1 inhibitor VX765 on cold-restraint stress-induced acute gastric ulcer in mice. Chin. J. Burn. 2017, 33, 688–693. [Google Scholar] [CrossRef]
- Sun, D.S.; Ho, P.H.; Chang, H.H. Soluble P-selectin rescues viper venom-induced mortality through anti-inflammatory properties and PSGL-1 pathway-mediated correction of hemostasis. Sci. Rep. 2016, 6, 35868. [Google Scholar] [CrossRef]
- Sun, D.S.; Chang, Y.W.; Kau, J.H.; Huang, H.H.; Ho, P.H.; Tzeng, Y.J.; Chang, H.H. Soluble P-selectin rescues mice from anthrax lethal toxin-induced mortality through PSGL-1 pathway-mediated correction of hemostasis. Virulence 2017, 8, 1216–1228. [Google Scholar] [CrossRef]
- Chen, J.L.; Cheng, T.T.; Huang, C.C.; Chang, H.H.; Lam, C.F. Dual phenotypic characteristics of P-selectin in a mouse model of hemorrhagic shock and hepatectomy. Heliyon 2023, 9, e18627. [Google Scholar] [CrossRef]
- Zimprich, A.; Garrett, L.; Deussing, J.M.; Wotjak, C.T.; Fuchs, H.; Gailus-Durner, V.; de Angelis, M.H.; Wurst, W.; Holter, S.M. A robust and reliable non-invasive test for stress responsivity in mice. Front. Behav. Neurosci. 2014, 8, 125. [Google Scholar] [CrossRef]
- Chu, X.; Zhou, Y.; Hu, Z.; Lou, J.; Song, W.; Li, J.; Liang, X.; Chen, C.; Wang, S.; Yang, B.; et al. 24-hour-restraint stress induces long-term depressive-like phenotypes in mice. Sci. Rep. 2016, 6, 32935. [Google Scholar] [CrossRef]
- Samuelsson, A.; Towers, T.L.; Ravetch, J.V. Anti-inflammatory activity of IVIG mediated through the inhibitory Fc receptor. Science 2001, 291, 484–486. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.S.; King, C.C.; Huang, H.S.; Shih, Y.L.; Lee, C.C.; Tsai, W.J.; Yu, C.C.; Chang, H.H. Antiplatelet autoantibodies elicited by dengue virus non-structural protein 1 cause thrombocytopenia and mortality in mice. J. Thromb. Haemost. 2007, 5, 2291–2299. [Google Scholar] [CrossRef] [PubMed]
- Lien, T.S.; Sun, D.S.; Chang, C.M.; Wu, C.Y.; Dai, M.S.; Chan, H.; Wu, W.S.; Su, S.H.; Lin, Y.Y.; Chang, H.H. Dengue virus and antiplatelet autoantibodies synergistically induce haemorrhage through Nlrp3-inflammasome and FcgammaRIII. Thromb. Haemost. 2015, 113, 1060–1070. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Wong, M.S.; Chen, C.W.; Hsieh, C.C.; Hung, S.C.; Sun, D.S.; Chang, H.H. Antibacterial property of Ag nanoparticle-impregnated N-doped titania films under visible light. Sci. Rep. 2015, 5, 11978. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.S.; Tseng, Y.H.; Wu, W.S.; Wong, M.S.; Chang, H.H. Visible Light-Responsive Platinum-Containing Titania Nanoparticle-Mediated Photocatalysis Induces Nucleotide Insertion, Deletion and Substitution Mutations. Nanomaterials 2016, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.F.; Lin, G.L.; Chu, S.C.; Chen, C.C.; Liou, Y.S.; Chang, H.H.; Sun, D.S. AQP0 is a novel surface marker for deciphering abnormal erythropoiesis. Stem Cell Res. Ther. 2021, 12, 274. [Google Scholar] [CrossRef]
- Chen, T.L.; Chiang, Y.W.; Lin, G.L.; Chang, H.H.; Lien, T.S.; Sheh, M.H.; Sun, D.S. Different effects of granulocyte colony-stimulating factor and erythropoietin on erythropoiesis. Stem Cell Res. Ther. 2018, 9, 119. [Google Scholar] [CrossRef]
- Liao, G.; Simone, J.; Simon, S.R. Paracrine downregulation of Fc gamma RIII in human monocyte-derived macrophages induced by phagocytosis of nonopsonized particles. Blood 1994, 83, 2294–2304. [Google Scholar] [CrossRef]
- Lien, T.S.; Sun, D.S.; Wu, W.S.; Chang, H.H. Simulation of Hemorrhage Pathogenesis in Mice through Dual Stimulation with Dengue Envelope Protein Domain III-Coated Nanoparticles and Antiplatelet Antibody. Int. J. Mol. Sci. 2023, 24, 9270. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.-C.; Munalisa, R.; Lee, H.-Y.; Lien, T.-S.; Chan, H.; Hung, S.-C.; Sun, D.-S.; Cheng, C.-F.; Chang, H.-H. Restraint Stress-Induced Immunosuppression Is Associated with Concurrent Macrophage Pyroptosis Cell Death in Mice. Int. J. Mol. Sci. 2023, 24, 12877. https://doi.org/10.3390/ijms241612877
Li C-C, Munalisa R, Lee H-Y, Lien T-S, Chan H, Hung S-C, Sun D-S, Cheng C-F, Chang H-H. Restraint Stress-Induced Immunosuppression Is Associated with Concurrent Macrophage Pyroptosis Cell Death in Mice. International Journal of Molecular Sciences. 2023; 24(16):12877. https://doi.org/10.3390/ijms241612877
Chicago/Turabian StyleLi, Chi-Cheng, Rina Munalisa, Hsuan-Yun Lee, Te-Sheng Lien, Hao Chan, Shih-Che Hung, Der-Shan Sun, Ching-Feng Cheng, and Hsin-Hou Chang. 2023. "Restraint Stress-Induced Immunosuppression Is Associated with Concurrent Macrophage Pyroptosis Cell Death in Mice" International Journal of Molecular Sciences 24, no. 16: 12877. https://doi.org/10.3390/ijms241612877