
Microglia and p38 MAPK Inhibitors Suppress Development of Mechanical Allodynia in Both Sexes in a Mouse Model of Antiretroviral-Induced Neuropathic Pain

Abstract
:
1. Introduction
2. Results
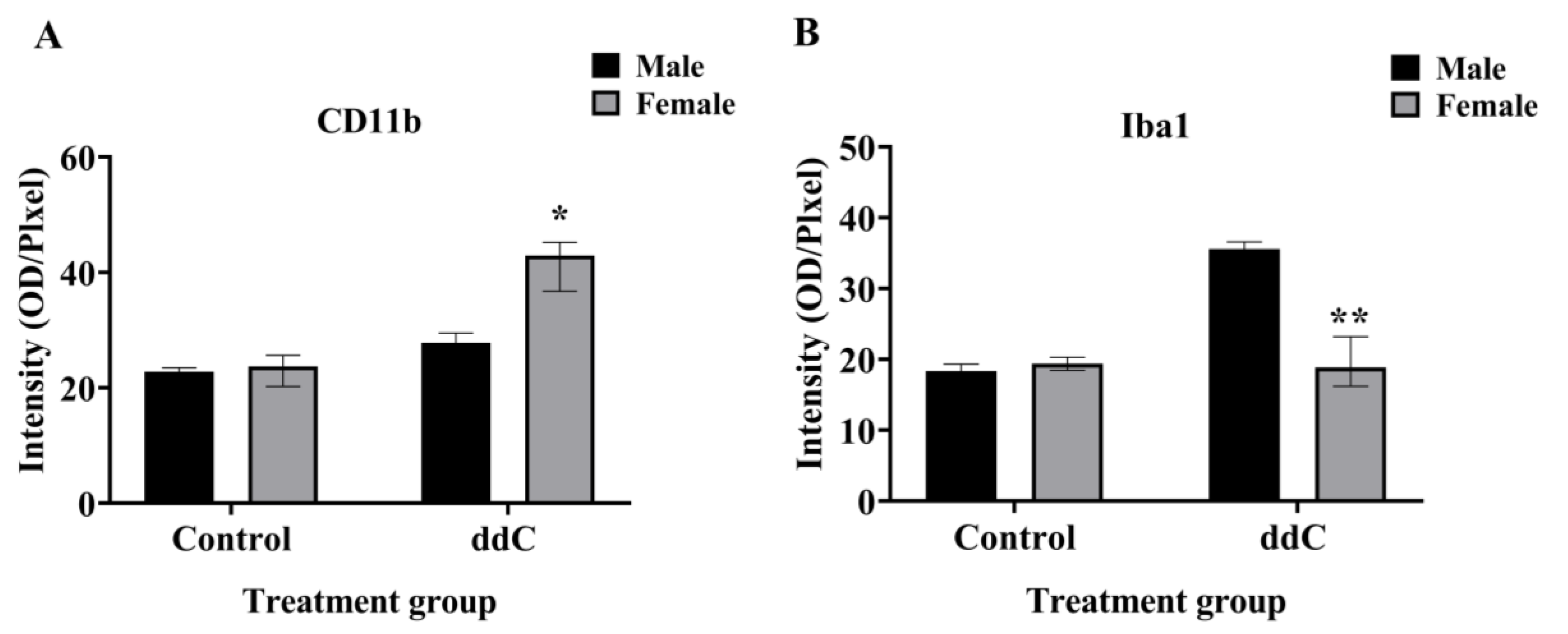
2.1. ddC Induced Microglia Reaction on the Lumbar Segment of the Spinal Cord of Both Male and Female Mice
2.2. ddC Induced p38 MAPK Activation on the Lumbar Segment of the Spinal Cord of Both Male and Female Mice
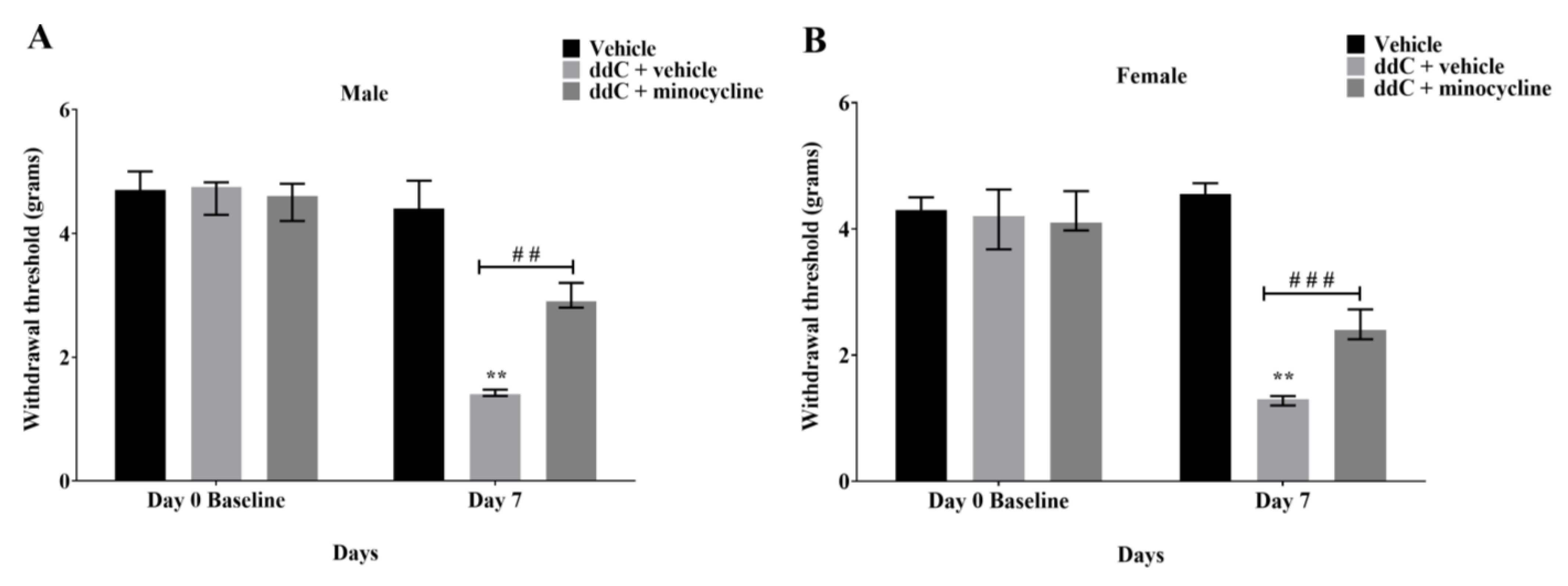
2.3. Minocycline Suppressed ddC-Induced Mechanical Allodynia in Both Male and Female Mice
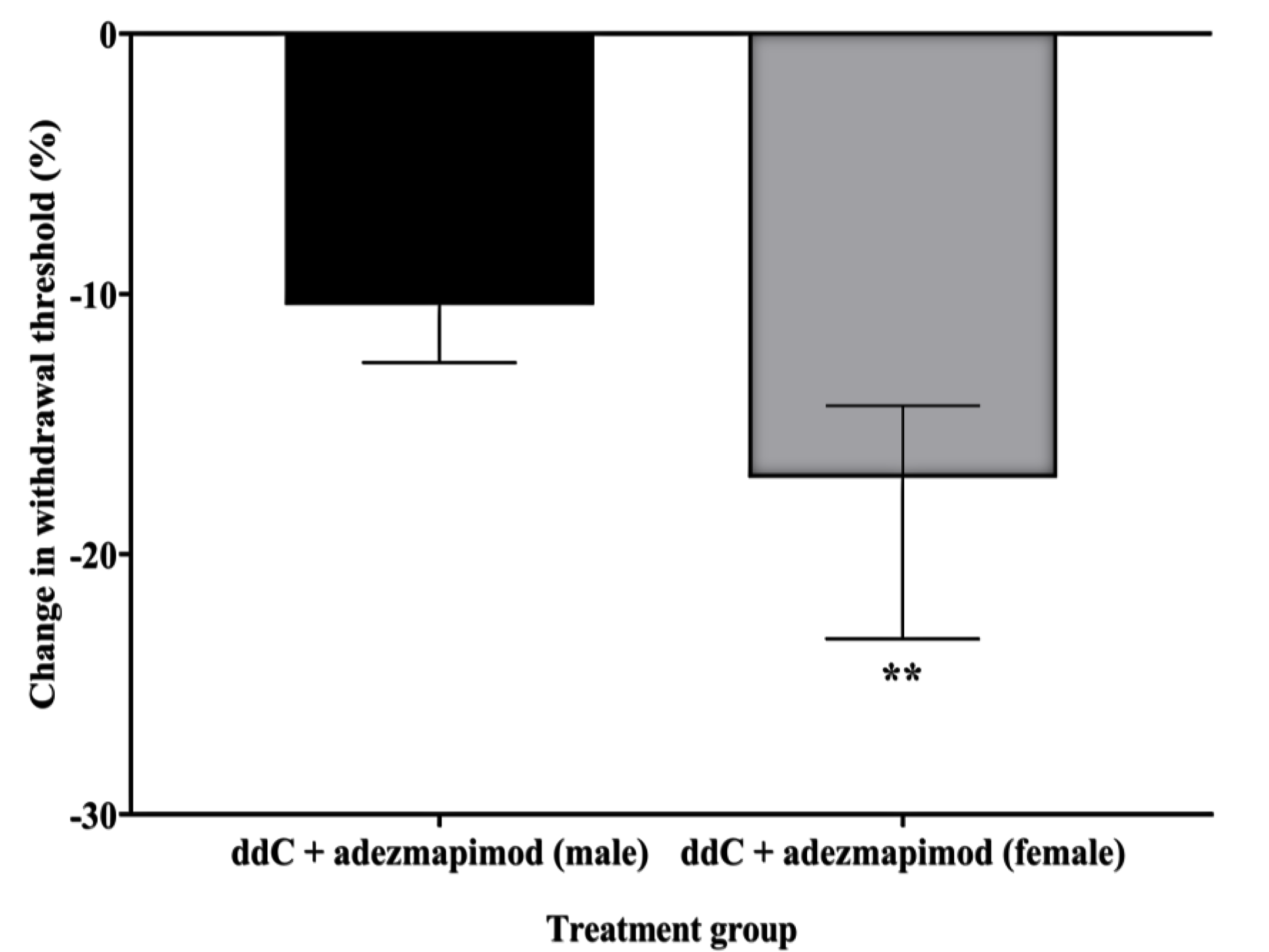
2.4. Inhibition of p38 MAPK Prevented ddC-Induced Mechanical Allodynia in Both Male and Female Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drug Preparation and Administration
4.3. Assessment of Mechanical Allodynia
4.4. Tissue Fixation, Processing, and Paraffine Embedding
4.5. Immunofluorescence
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scholz, J.; Finnerup, N.B.; Attal, N.; Aziz, Q.; Baron, R.; Bennett, M.I.; Benoliel, R.; Cohen, M.; Cruccu, G.; Davis, K.D.; et al. The IASP classification of chronic pain for ICD-11: Chronic neuropathic pain. Pain 2019, 160, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Ji, R.R.; Xu, Z.Z.; Gao, Y.J. Emerging targets in neuroinflammation-driven chronic pain. Nat. Rev. Drug Discov. 2014, 13, 533–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Hecke, O.; Austin, S.K.; Khan, R.A.; Smith, B.H.; Torrance, N. Neuropathic pain in the general population: A systematic review of epidemiological studies. Pain 2014, 155, 654–662. [Google Scholar] [CrossRef]
- Gabbai, A.A.; Castelo, A.; Oliveira, A.S. HIV peripheral neuropathy. Handb. Clin. Neurol. 2013, 115, 515–529. [Google Scholar] [CrossRef] [PubMed]
- Saag, M.S.; Gandhi, R.T.; Hoy, J.F.; Landovitz, R.J.; Thompson, M.A.; Sax, P.E.; Smith, D.M.; Benson, C.A.; Buchbinder, S.P.; Del Rio, C.; et al. Antiretroviral Drugs for Treatment and Prevention of HIV Infection in Adults: 2020 Recommendations of the International Antiviral Society-USA Panel. JAMA 2020, 324, 1651–1669. [Google Scholar] [CrossRef]
- Dalakas, M.C. Peripheral neuropathy and antiretroviral drugs. J. Peripher. Nerv. Syst. 2001, 6, 14–20. [Google Scholar] [CrossRef]
- Alhadlaq, M.W.; Masocha, W. Sex Differences in the Expression of Neuroimmune Molecules in the Spinal Cord of a Mouse Model of Antiretroviral-Induced Neuropathic Pain. Biomedicines 2023, 11, 875. [Google Scholar] [CrossRef] [PubMed]
- Aly, E.; Khajah, M.A.; Masocha, W. β-Caryophyllene, a CB2-Receptor-Selective Phytocannabinoid, Suppresses Mechanical Allodynia in a Mouse Model of Antiretroviral-Induced Neuropathic Pain. Molecules 2019, 25, 106. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, M.; Shigemoto-Mogami, Y.; Koizumi, S.; Mizokoshi, A.; Kohsaka, S.; Salter, M.W.; Inoue, K. P2X4 receptors induced in spinal microglia gate tactile allodynia after nerve injury. Nature 2003, 424, 778–783. [Google Scholar] [CrossRef]
- Zhuang, Z.Y.; Kawasaki, Y.; Tan, P.H.; Wen, Y.R.; Huang, J.; Ji, R.R. Role of the CX3CR1/p38 MAPK pathway in spinal microglia for the development of neuropathic pain following nerve injury-induced cleavage of fractalkine. Brain Behav. Immun. 2007, 21, 642–651. [Google Scholar] [CrossRef] [Green Version]
- Sorge, R.E.; LaCroix-Fralish, M.L.; Tuttle, A.H.; Sotocinal, S.G.; Austin, J.S.; Ritchie, J.; Chanda, M.L.; Graham, A.C.; Topham, L.; Beggs, S.; et al. Spinal cord Toll-like receptor 4 mediates inflammatory and neuropathic hypersensitivity in male but not female mice. J. Neurosci. 2011, 31, 15450–15454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beggs, S.; Trang, T.; Salter, M.W. P2X4R+ microglia drive neuropathic pain. Nat. Neurosci. 2012, 15, 1068–1073. [Google Scholar] [CrossRef] [PubMed]
- Narita, M.; Yoshida, T.; Nakajima, M.; Narita, M.; Miyatake, M.; Takagi, T.; Yajima, Y.; Suzuki, T. Direct evidence for spinal cord microglia in the development of a neuropathic pain-like state in mice. J. Neurochem. 2006, 97, 1337–1348. [Google Scholar] [CrossRef]
- Guasti, L.; Richardson, D.; Jhaveri, M.; Eldeeb, K.; Barrett, D.; Elphick, M.R.; Alexander, S.P.; Kendall, D.; Michael, G.J.; Chapman, V. Minocycline treatment inhibits microglial activation and alters spinal levels of endocannabinoids in a rat model of neuropathic pain. Mol. Pain 2009, 5, 35. [Google Scholar] [CrossRef]
- Mika, J.; Osikowicz, M.; Rojewska, E.; Korostynski, M.; Wawrzczak-Bargiela, A.; Przewlocki, R.; Przewlocka, B. Differential activation of spinal microglial and astroglial cells in a mouse model of peripheral neuropathic pain. Eur. J. Pharmacol. 2009, 623, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Ledeboer, A.; Sloane, E.M.; Milligan, E.D.; Frank, M.G.; Mahony, J.H.; Maier, S.F.; Watkins, L.R. Minocycline attenuates mechanical allodynia and proinflammatory cytokine expression in rat models of pain facilitation. Pain 2005, 115, 71–83. [Google Scholar] [CrossRef]
- Raghavendra, V.; Tanga, F.; DeLeo, J.A. Inhibition of microglial activation attenuates the development but not existing hypersensitivity in a rat model of neuropathy. J. Pharmacol. Exp. Ther. 2003, 306, 624–630. [Google Scholar] [CrossRef] [Green Version]
- Masocha, W. Paclitaxel-induced hyposensitivity to nociceptive chemical stimulation in mice can be prevented by treatment with minocycline. Sci. Rep. 2014, 4, 6719. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.X.; Zhuang, Z.Y.; Woolf, C.J.; Ji, R.R. p38 mitogen-activated protein kinase is activated after a spinal nerve ligation in spinal cord microglia and dorsal root ganglion neurons and contributes to the generation of neuropathic pain. J. Neurosci. 2003, 23, 4017–4022. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, M.; Mizokoshi, A.; Shigemoto-Mogami, Y.; Koizumi, S.; Inoue, K. Activation of p38 mitogen-activated protein kinase in spinal hyperactive microglia contributes to pain hypersensitivity following peripheral nerve injury. Glia 2004, 45, 89–95. [Google Scholar] [CrossRef]
- Wen, Y.R.; Suter, M.R.; Ji, R.R.; Yeh, G.C.; Wu, Y.S.; Wang, K.C.; Kohno, T.; Sun, W.Z.; Wang, C.C. Activation of p38 mitogen-activated protein kinase in spinal microglia contributes to incision-induced mechanical allodynia. Anesthesiology 2009, 110, 155–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, A.K.; D’Aquisto, F.; Gentry, C.; Marchand, F.; McMahon, S.B.; Malcangio, M. Rapid co-release of interleukin 1beta and caspase 1 in spinal cord inflammation. J. Neurochem. 2006, 99, 868–880. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.R.; Suter, M.R. p38 MAPK, microglial signaling, and neuropathic pain. Mol. Pain 2007, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Sukhtankar, D.; Okun, A.; Chandramouli, A.; Nelson, M.A.; Vanderah, T.W.; Cress, A.E.; Porreca, F.; King, T. Inhibition of p38-MAPK signaling pathway attenuates breast cancer induced bone pain and disease progression in a murine model of cancer-induced bone pain. Mol. Pain 2011, 7, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taves, S.; Berta, T.; Liu, D.L.; Gan, S.; Chen, G.; Kim, Y.H.; Van de Ven, T.; Laufer, S.; Ji, R.R. Spinal inhibition of p38 MAP kinase reduces inflammatory and neuropathic pain in male but not female mice: Sex-dependent microglial signaling in the spinal cord. Brain Behav. Immun. 2016, 55, 70–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorge, R.E.; Mapplebeck, J.C.; Rosen, S.; Beggs, S.; Taves, S.; Alexander, J.K.; Martin, L.J.; Austin, J.S.; Sotocinal, S.G.; Chen, D.; et al. Different immune cells mediate mechanical pain hypersensitivity in male and female mice. Nat. Neurosci. 2015, 18, 1081–1083. [Google Scholar] [CrossRef] [Green Version]
- Beery, A.K.; Zucker, I. Sex bias in neuroscience and biomedical research. Neurosci. Biobehav. Rev. 2011, 35, 565–572. [Google Scholar] [CrossRef] [Green Version]
- Torrance, N.; Smith, B.H.; Bennett, M.I.; Lee, A.J. The epidemiology of chronic pain of predominantly neuropathic origin. Results from a general population survey. J. Pain 2006, 7, 281–289. [Google Scholar] [CrossRef]
- Fillingim, R.B.; King, C.D.; Ribeiro-Dasilva, M.C.; Rahim-Williams, B.; Riley, J.L., 3rd. Sex, gender, and pain: A review of recent clinical and experimental findings. J. Pain 2009, 10, 447–485. [Google Scholar] [CrossRef] [Green Version]
- Mapplebeck, J.C.S.; Dalgarno, R.; Tu, Y.; Moriarty, O.; Beggs, S.; Kwok, C.H.T.; Halievski, K.; Assi, S.; Mogil, J.S.; Trang, T.; et al. Microglial P2X4R-evoked pain hypersensitivity is sexually dimorphic in rats. Pain 2018, 159, 1752–1763. [Google Scholar] [CrossRef]
- Mapplebeck, J.C.S.; Beggs, S.; Salter, M.W. Sex differences in pain: A tale of two immune cells. Pain 2016, 157 (Suppl. S1), S2–S6. [Google Scholar] [CrossRef]
- Rosen, S.; Ham, B.; Mogil, J.S. Sex differences in neuroimmunity and pain. J. Neurosci. Res. 2017, 95, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Prinz, M.; Masuda, T.; Wheeler, M.A.; Quintana, F.J. Microglia and Central Nervous System-Associated Macrophages-From Origin to Disease Modulation. Annu. Rev. Immunol. 2021, 39, 251–277. [Google Scholar] [CrossRef]
- Dance, A. Why the sexes don’t feel pain the same way. Nature 2019, 567, 448–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, G.; Robinson, F.; Beers Gibson, T.; Xu, B.E.; Karandikar, M.; Berman, K.; Cobb, M.H. Mitogen-activated protein (MAP) kinase pathways: Regulation and physiological functions. Endocr. Rev. 2001, 22, 153–183. [Google Scholar] [CrossRef] [Green Version]
- Nazarian, S.; Akhondi, H. Minocycline. In StatPearls; Ineligible Companies: Treasure Island, FL, USA, 2023. [Google Scholar]
- Garrido-Mesa, N.; Zarzuelo, A.; Galvez, J. Minocycline: Far beyond an antibiotic. Br. J. Pharmacol. 2013, 169, 337–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Q.; Xiong, J.; Yuan, Y.; Ruan, Z.; Zhang, Y.; Chai, B.; Li, L.; Cai, S.; Xiao, J.; Wu, Y.; et al. Minocycline improves the functional recovery after traumatic brain injury via inhibition of aquaporin-4. Int. J. Biol. Sci. 2022, 18, 441–458. [Google Scholar] [CrossRef]
- Strickland, B.A.; Barisano, G.; Abedi, A.; Shiroishi, M.S.; Cen, S.; Emanuel, B.; Bulic, S.; Kim-Tenser, M.; Nguyen, P.; Giannotta, S.L.; et al. Minocycline decreases blood-brain barrier permeability following aneurysmal subarachnoid hemorrhage: A randomized, double-blind, controlled trial. J. Neurosurg. 2022, 136, 1251–1259. [Google Scholar] [CrossRef]
- Plane, J.M.; Shen, Y.; Pleasure, D.E.; Deng, W. Prospects for minocycline neuroprotection. Arch. Neurol. 2010, 67, 1442–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.S.; Shi, Q.; Mendoza, T.; Lin, S.; Chang, J.Y.; Bokhari, R.H.; Lin, H.K.; Garcia-Gonzalez, A.; Kamal, M.; Cleeland, C.S.; et al. Minocycline Reduces Chemoradiation-Related Symptom Burden in Patients with Non-Small Cell Lung Cancer: A Phase 2 Randomized Trial. Int. J. Radiat. Oncol. Biol. Phys. 2020, 106, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Syngle, A.; Verma, I.; Krishan, P.; Garg, N.; Syngle, V. Minocycline improves peripheral and autonomic neuropathy in type 2 diabetes: MIND study. Neurol. Sci. 2014, 35, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Narang, T.; Arshdeep; Dogra, S. Minocycline in leprosy patients with recent onset clinical nerve function impairment. Dermatol. Ther. 2017, 30, e12404. [Google Scholar] [CrossRef] [PubMed]
- Starobova, H.; Mueller, A.; Allavena, R.; Lohman, R.J.; Sweet, M.J.; Vetter, I. Minocycline Prevents the Development of Mechanical Allodynia in Mouse Models of Vincristine-Induced Peripheral Neuropathy. Front. Neurosci. 2019, 13, 653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.C.; Lu, N.; Cui, Y.; Yang, T.; Zhao, Z.Q.; Xin, W.J.; Liu, X.G. Prevention of paclitaxel-induced allodynia by minocycline: Effect on loss of peripheral nerve fibers and infiltration of macrophages in rats. Mol. Pain 2010, 6, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hains, B.C.; Waxman, S.G. Activated microglia contribute to the maintenance of chronic pain after spinal cord injury. J. Neurosci. 2006, 26, 4308–4317. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.J.; Kress, B.; Han, X.; Moll, K.; Peng, W.; Ji, R.R.; Nedergaard, M. Astrocytic CX43 hemichannels and gap junctions play a crucial role in development of chronic neuropathic pain following spinal cord injury. Glia 2012, 60, 1660–1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berta, T.; Qadri, Y.J.; Chen, G.; Ji, R.R. Microglial Signaling in Chronic Pain with a Special Focus on Caspase 6, p38 MAP Kinase, and Sex Dependence. J. Dent. Res. 2016, 95, 1124–1131. [Google Scholar] [CrossRef] [Green Version]
- Beery, A.K. Inclusion of females does not increase variability in rodent research studies. Curr. Opin. Behav. Sci. 2018, 23, 143–149. [Google Scholar] [CrossRef]
- Zeng, P.Y.; Tsai, Y.H.; Lee, C.L.; Ma, Y.K.; Kuo, T.H. Minimal influence of estrous cycle on studies of female mouse behaviors. Front. Mol. Neurosci. 2023, 16, 1146109. [Google Scholar] [CrossRef]
- Hassan, S.; Muere, A.; Einstein, G. Ovarian hormones and chronic pain: A comprehensive review. Pain 2014, 155, 2448–2460. [Google Scholar] [CrossRef]
- Stening, K.; Eriksson, O.; Wahren, L.; Berg, G.; Hammar, M.; Blomqvist, A. Pain sensations to the cold pressor test in normally menstruating women: Comparison with men and relation to menstrual phase and serum sex steroid levels. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R1711–R1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.M.; Dragunow, M. The human side of microglia. Trends Neurosci. 2014, 37, 125–135. [Google Scholar] [CrossRef]
- Sumitani, M.; Ueda, H.; Hozumi, J.; Inoue, R.; Kogure, T.; Yamada, Y.; Kogure, T. Minocycline Does Not Decrease Intensity of Neuropathic Pain Intensity, But Does Improve Its Affective Dimension. J. Pain Palliat. Care Pharmacother. 2016, 30, 31–35. [Google Scholar] [CrossRef]
- Vanelderen, P.; Van Zundert, J.; Kozicz, T.; Puylaert, M.; De Vooght, P.; Mestrum, R.; Heylen, R.; Roubos, E.; Vissers, K. Effect of minocycline on lumbar radicular neuropathic pain: A randomized, placebo-controlled, double-blind clinical trial with amitriptyline as a comparator. Anesthesiology 2015, 122, 399–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvathy, S.S.; Masocha, W. Coadministration of indomethacin and minocycline attenuates established paclitaxel-induced neuropathic thermal hyperalgesia: Involvement of cannabinoid CB1 receptors. Sci. Rep. 2015, 5, 10541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richner, M.; Jager, S.B.; Siupka, P.; Vaegter, C.B. Hydraulic Extrusion of the Spinal Cord and Isolation of Dorsal Root Ganglia in Rodents. J. Vis. Exp. 2017, 2017, 55226. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group | Number of Male Mice | Number of Female Mice |
|---|---|---|
| vehicle only * | 16 | 16 |
| ddC * | 16 | 16 |
| ddC + minocycline | 7 | 6 |
| ddC + adezmapimod | 6 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alhadlaq, M.W.; Masocha, W. Microglia and p38 MAPK Inhibitors Suppress Development of Mechanical Allodynia in Both Sexes in a Mouse Model of Antiretroviral-Induced Neuropathic Pain. Int. J. Mol. Sci. 2023, 24, 12805. https://doi.org/10.3390/ijms241612805
Alhadlaq MW, Masocha W. Microglia and p38 MAPK Inhibitors Suppress Development of Mechanical Allodynia in Both Sexes in a Mouse Model of Antiretroviral-Induced Neuropathic Pain. International Journal of Molecular Sciences. 2023; 24(16):12805. https://doi.org/10.3390/ijms241612805
Chicago/Turabian StyleAlhadlaq, Maryam W., and Willias Masocha. 2023. "Microglia and p38 MAPK Inhibitors Suppress Development of Mechanical Allodynia in Both Sexes in a Mouse Model of Antiretroviral-Induced Neuropathic Pain" International Journal of Molecular Sciences 24, no. 16: 12805. https://doi.org/10.3390/ijms241612805