
Bta-miR-484 Targets SFRP1 and Affects Preadipocytes Proliferation, Differentiation, and Apoptosis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
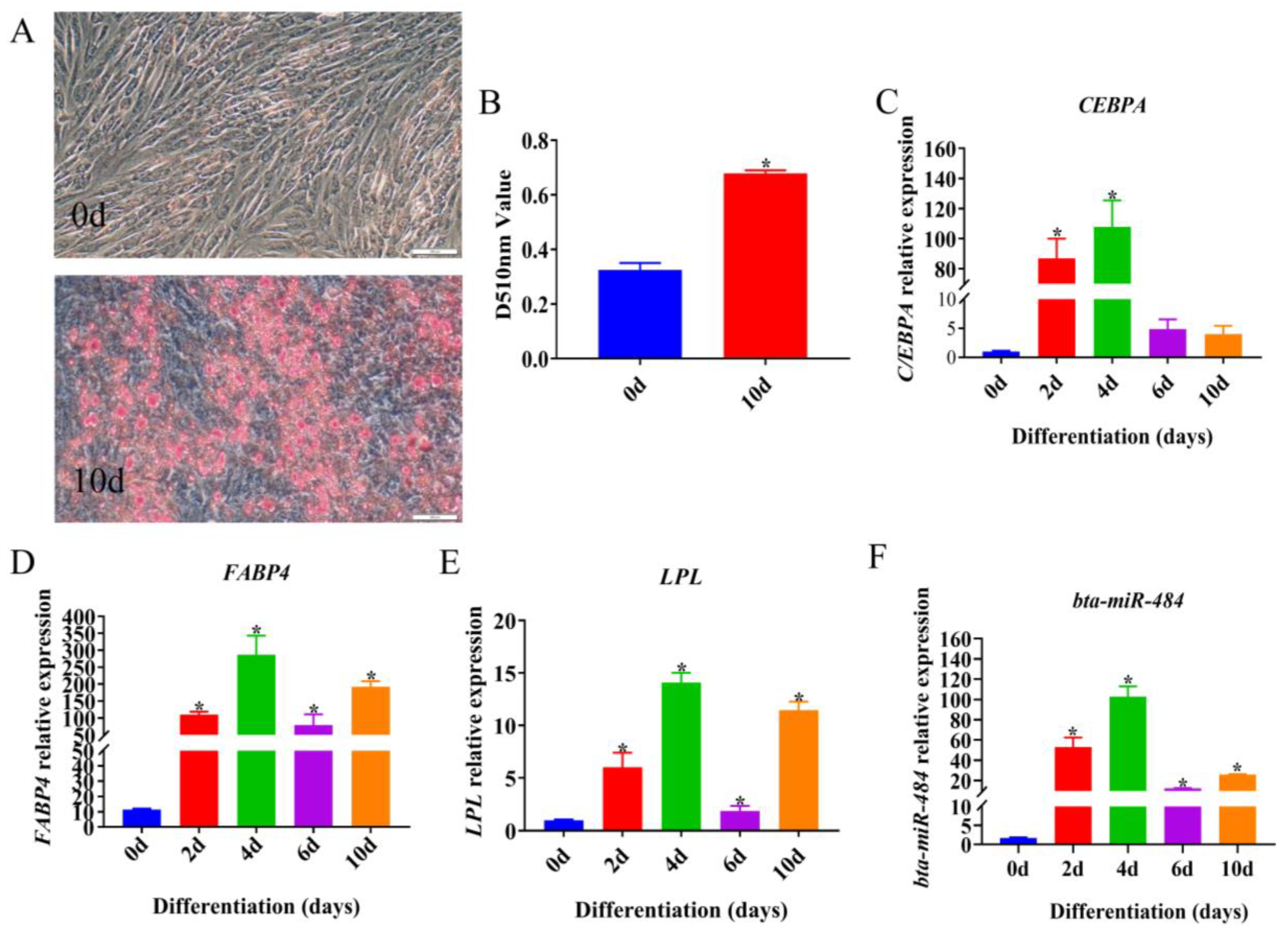
2.1. Temporal Expression Profile of bta-miR-484 during Adipose Differentiation
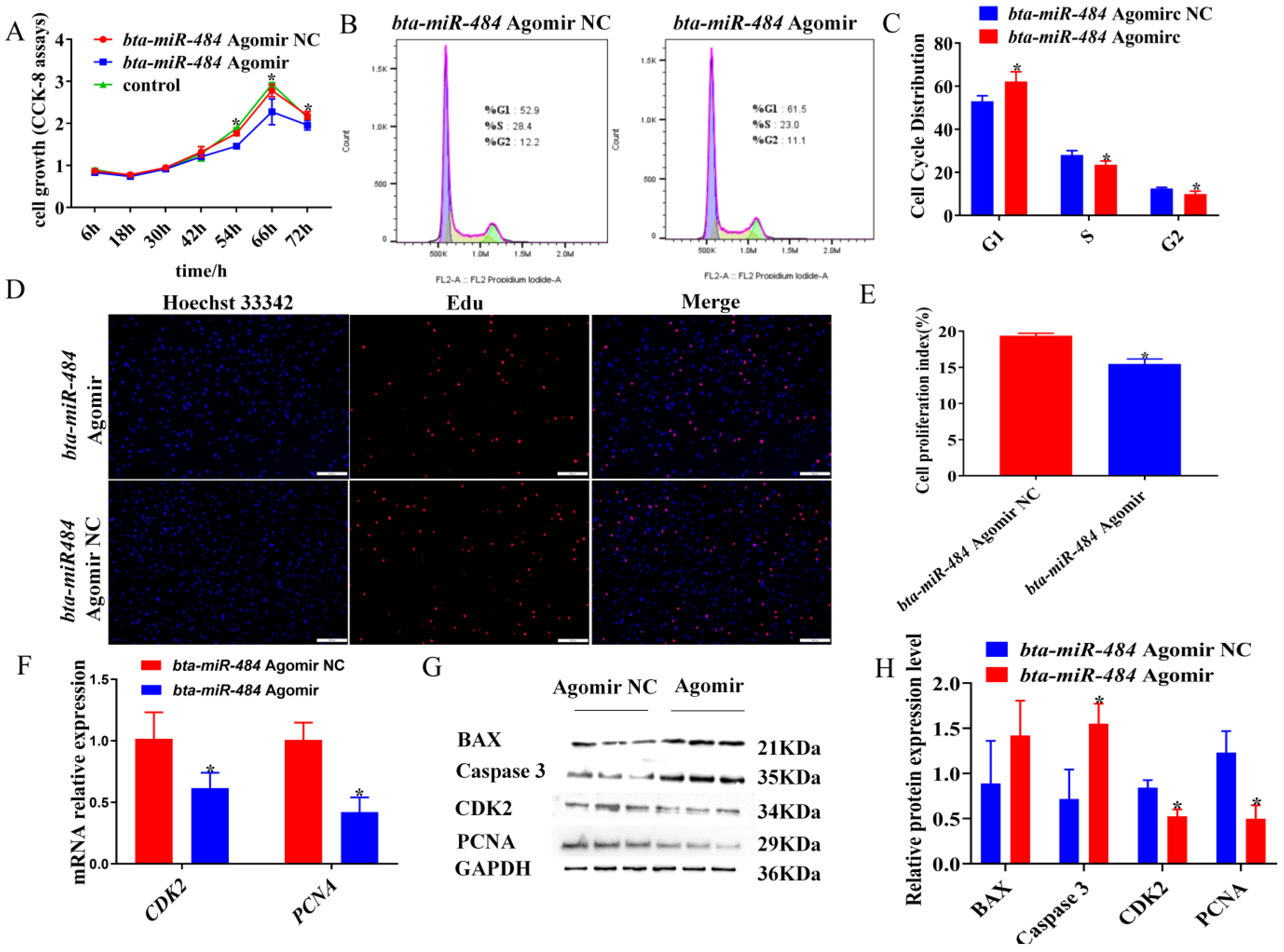
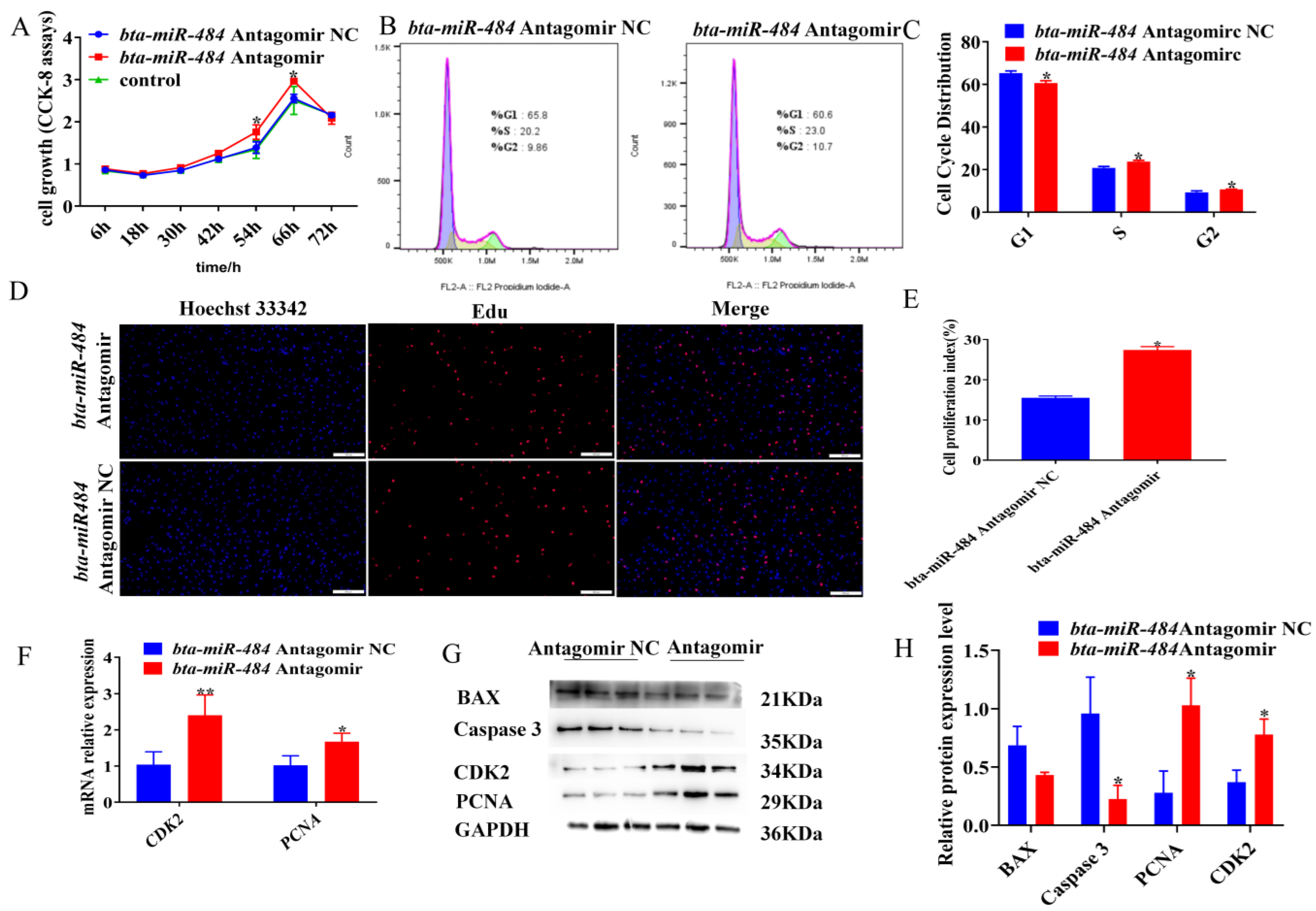
2.2. Bta-miR-484 Inhibits Adipocyte Proliferation
2.3. Bta-miR-484 Promotes Adipocyte Apoptosis
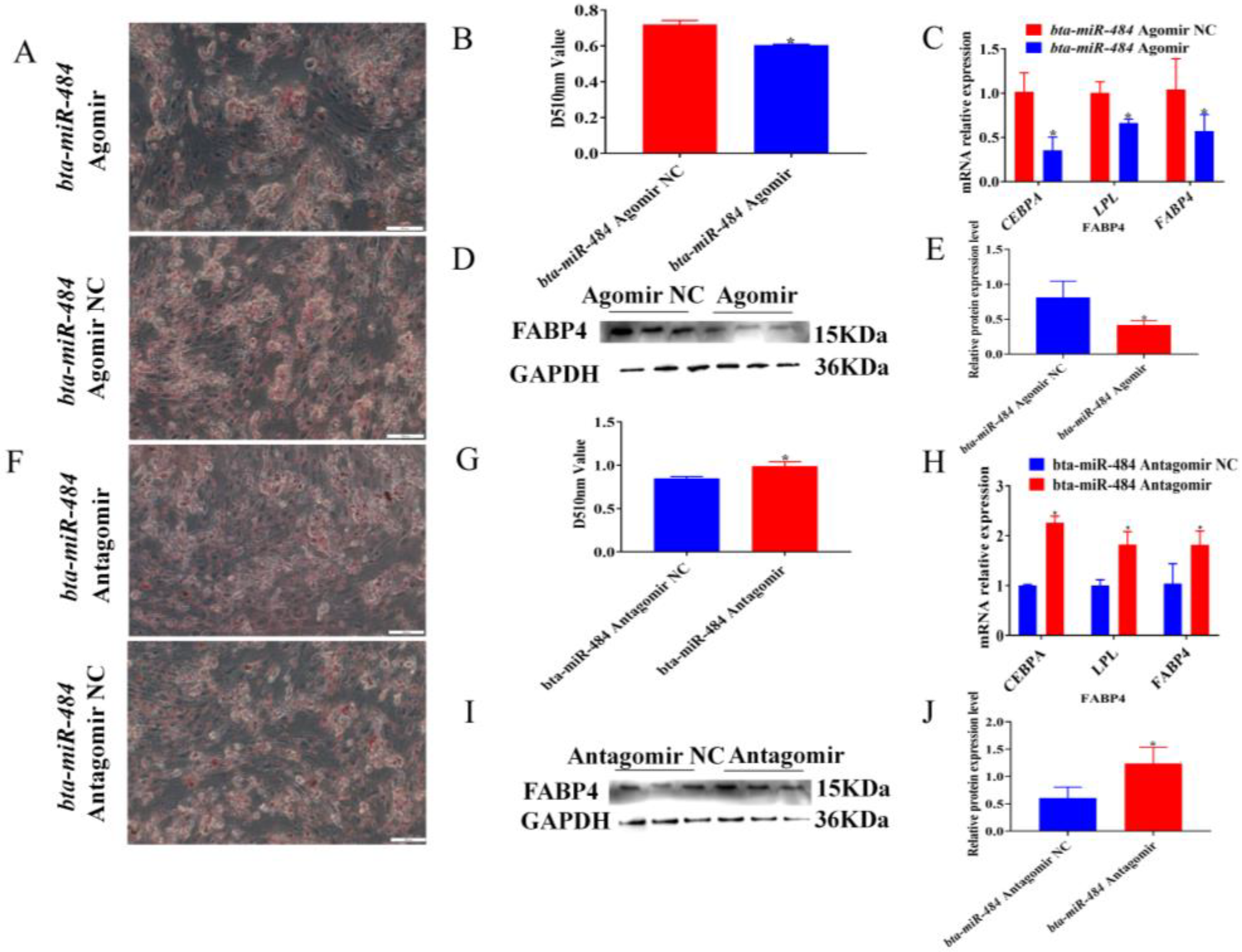
2.4. Bta-miR-484 Inhibits Adipocyte Differentiation
2.5. RNA Sequencing Analysis
2.6. Bta-miR-484 Targeted Binding of SFRP1
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Adipocyte Isolation, Culture and Differentiation
4.3. Oil Red O Staining
4.4. CCK-8 Assay and EdU Staining
4.5. Flow Cytometery
4.6. RNA Extraction and Quantitative Real-Time PCR (qPCR)
4.7. Western Blotting
4.8. RNA-Seq
4.9. Target Gene Prediction
4.10. Dual Luciferase Reporter Gene Assay
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kang, Z.; Zhang, S.; Jiang, E.; Wan, F.; Lan, X.; Liu, M. Mir-193b Regulates the Differentiation, Proliferation, and Apoptosis of Bovine Adipose Cells by Targeting the ACSS2/AKT Axis. Animals 2020, 10, 1265. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zhang, P.; Gan, M.; Zhao, X.; Xu, Y.; Li, Q.; Jiang, Y.; Tang, G.; Li, M.; Wang, J.; et al. MicroRNA-204-5p regulates 3T3-L1 preadipocyte proliferation, apoptosis and differentiation. Gene 2018, 668, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Roos, J.; Dahlhaus, M.; Funcke, J.B.; Kustermann, M.; Strauss, G.; Halbgebauer, D.; Boldrin, E.; Holzmann, K.; Möller, P.; Trojanowski, B.M.; et al. miR-146a regulates insulin sensitivity via NPR3. Cell Mol. Life Sci. 2021, 78, 2987–3003. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Zhang, D.; Katayama, A.; Kurooka, N.; Sugawara, R.; Albuayjan, H.H.H.; Nakatsuka, A.; Eguchi, J.; Wada, J. Adipocyte-Specific Inhibition of Mir221/222 Ameliorates Diet-Induced Obesity Through Targeting Ddit4. Front. Endocrinol. 2022, 12, 750261. [Google Scholar] [CrossRef]
- Murata, Y.; Yamashiro, T.; Kessoku, T.; Jahan, I.; Usuda, H.; Tanaka, T.; Okamoto, T.; Nakajima, A.; Wada, K. Up-Regulated MicroRNA-27b Promotes Adipocyte Differentiation via Induction of Acyl-CoA Thioesterase 2 Expression. Biomed. Res. Int. 2019, 2019, 2916243. [Google Scholar] [CrossRef] [Green Version]
- Acharya, A.; Berry, D.C.; Zhang, H.; Jiang, Y.; Jones, B.T.; Hammer, R.E.; Graff, J.M.; Mendell, J.T. miR-26 suppresses adipocyte progenitor differentiation and fat production by targeting Fbxl19. Genes Dev. 2019, 33, 1367–1380. [Google Scholar] [CrossRef] [Green Version]
- Ahonen, M.A.; Haridas, P.A.N.; Mysore, R.; Wabitsch, M.; Fischer-Posovszky, P.; Olkkonen, V.M. miR-107 inhibits CDK6 expression, differentiation, and lipid storage in human adipocytes. Mol. Cell Endocrinol. 2019, 479, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Raza, S.H.A.; Ma, X.; Wang, J.; Wang, X.; Liang, C.; Yang, X.; Mei, C.; Suhail, S.M.; Zan, L. Bovine Pre-adipocyte Adipogenesis Is Regulated by bta-miR-150 Through mTOR Signaling. Front. Genet. 2021, 12, 636550. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, L.; Gu, Y.; Chang, Y.; Liang, C.; Guo, X.; Bao, P.; Chu, M.; Ding, X.; Yan, P. Bta-miR-2400 Targets SUMO1 to Affect Yak Preadipocytes Proliferation and Differentiation. Biology 2021, 10, 949. [Google Scholar] [CrossRef]
- Khan, R.; Raza, S.H.A.; Junjvlieke, Z.; Wang, X.; Wang, H.; Cheng, G.; Mei, C.; Elsaeid Elnour, I.; Zan, L. Bta-miR-149-5p inhibits proliferation and differentiation of bovine adipocytes through targeting CRTCs at both transcriptional and posttranscriptional levels. J. Cell Physiol. 2020, 235, 5796–5810. [Google Scholar] [CrossRef]
- Chen, X.; Raza, S.H.A.; Cheng, G.; Ma, X.; Wang, J.; Zan, L. Bta-miR-376a Targeting KLF15 Interferes with Adipogenesis Signaling Pathway to Promote Differentiation of Qinchuan Beef Cattle Preadipocytes. Animals 2020, 10, 2362. [Google Scholar] [CrossRef]
- Castrillón-Betancur, J.C.; Urcuqui-Inchima, S. Overexpression of miR-484 and miR-744 in Vero cells alters Dengue virus replication. Mem. Inst. Oswaldo Cruz 2017, 112, 281–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masoodi Khabar, P.; Ghydari, M.E.; Vazifeh Shiran, N.; Shirazy, M.; Hamidpour, M. Platelet MicroRNA-484 as a Novel Diagnostic Biomarker for Acute Coronary Syndrome. Lab. Med. 2023, 54, 256–261. [Google Scholar] [CrossRef]
- Mohamed, D.A.W.; Selim, H.M.; Elmazny, A.; Genena, A.; Nabil, M.M. Apoptotic protease activating factor-1 gene and MicroRNA-484: A possible interplay in relapsing remitting multiple sclerosis. Mult. Scler. Relat. Disord. 2022, 58, 103502. [Google Scholar] [CrossRef] [PubMed]
- El-Maraghy, S.A.; Adel, O.; Zayedm, N.; Yosry, A.; El-Nahaas, S.M.; Gibriel, A.A. Circulatory miRNA-484, 524, 615 and 628 expression profiling in HCV mediated HCC among Egyptian patients; implications for diagnosis and staging of hepatic cirrhosis and fibrosis. J. Adv. Res. 2019, 22, 57–66. [Google Scholar] [CrossRef]
- Konaka, T.; Kawami, M.; Yamamoto, A.; Yumoto, R.; Takano, M. miR-484: A Possible Indicator of Drug-Induced Pulmonary Fibrosis. J. Pharm. Pharm. Sci. 2020, 23, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Tang, W.; Dorsey, T.H.; Ambs, S. miR-484 is associated with disease recurrence and promotes migration in prostate cancer. Biosci. Rep. 2020, 40, BSR20191028. [Google Scholar] [CrossRef] [PubMed]
- Vecchione, A.; Belletti, B.; Lovat, F.; Volinia, S.; Chiappetta, G.; Giglio, S.; Sonego, M.; Cirombella, R.; Onesti, E.C.; Pellegrini, P.; et al. A microRNA signature defines chemoresistance in ovarian cancer through modulation of angiogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, 9845–9850. [Google Scholar] [CrossRef]
- Merhautova, J.; Hezova, R.; Poprach, A.; Kovarikova, A.; Radova, L.; Svoboda, M.; Vyzula, R.; Demlova, R.; Slaby, O. miR-155 and miR-484 Are Associated with Time to Progression in Metastatic Renal Cell Carcinoma Treated with Sunitinib. Biomed. Res. Int. 2015, 2015, 941980. [Google Scholar] [CrossRef] [Green Version]
- Zare, A.; Ahadi, A.; Larki, P.; Omrani, M.D.; Zali, M.R.; Alamdari, N.M.; Ghaedi, H. The clinical significance of miR-335, miR-124, miR-218 and miR-484 downregulation in gastric cancer. Mol. Biol. Rep. 2018, 45, 1587–1595. [Google Scholar] [CrossRef]
- Allach, E.l.; Khattabi, L.; Heide, S.; Caberg, J.H.; Andrieux, J.; Doco Fenzy, M.; Vincent-Delorme, C.; Callier, P.; Chantot-Bastaraud, S.; Afenjar, A.; et al. 16p13.11 microduplication in 45 new patients: Refined clinical significance and genotype-phenotype correlations. J. Med. Genet. 2020, 57, 301–307. [Google Scholar] [CrossRef]
- Bradshaw, N.J.; Ukkola-Vuoti, L.; Pankakoski, M.; Zheutlin, A.B.; Ortega-Alonso, A.; Torniainen-Holm, M.; Sinha, V.; Therman, S.; Paunio, T.; Suvisaari, J.; et al. The NDE1 genomic locus can affect treatment of psychiatric illness through gene expression changes related to microRNA-484. Open Biol. 2017, 7, 170153. [Google Scholar] [CrossRef] [PubMed]
- Du, X. Molecular Characteristics of H-FABP Gene and Identification of miRNAs Associated with Intramuscular Fat Deposition in Beef Cattle; Shenyang Agricultural University: Shenyang, China, 2016. [Google Scholar]
- Rosen, E.D. The transcriptional basis of adipocyte development. Prostaglandins Leukot. Essent. Fat. Acids 2005, 73, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Tontonoz, P.; Spiegelman, B.M. Fat and beyond: The diverse biology of PPARgamma. Annu. Rev. Biochem. 2008, 77, 289–312. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, X.; Liang, C.; Bao, P.; Ding, X.; Chu, M.; Jia, C.; Guo, X.; Yan, P. MicroRNA-200a regulates adipocyte differentiation in the domestic yak Bos grunniens. Gene 2018, 650, 41–48. [Google Scholar] [CrossRef]
- Fagundes, R.; Teixeira, L.K. Cyclin E/CDK2: DNA Replication, Replication Stress and Genomic Instability. Front. Cell Dev. Biol. 2021, 9, 774845. [Google Scholar] [CrossRef]
- González-Magaña, A.; Blanco, F.J. Human PCNA Structure, Function and Interactions. Biomolecules 2020, 10, 570. [Google Scholar] [CrossRef] [Green Version]
- Eskandari, E.; Eaves, C.J. Paradoxical roles of caspase-3 in regulating cell survival, proliferation, and tumorigenesis. J. Cell Biol. 2022, 221, e202201159. [Google Scholar] [CrossRef]
- Araya, L.E.; Soni, I.V.; Hardy, J.A.; Julien, O. Deorphanizing Caspase-3 and Caspase-9 Substrates in and out of Apoptosis with Deep Substrate Profiling. ACS Chem. Biol. 2021, 16, 2280–2296. [Google Scholar] [CrossRef]
- Jensen, K.; WuWong, D.J.; Wong, S.; Matsuyama, M.; Matsuyama, S. Pharmacological inhibition of Bax-induced cell death: Bax-inhibiting peptides and small compounds inhibiting Bax. Exp. Biol. Med. 2019, 244, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, X.; Mu, H.; Mei, Q.; Liu, Y.; Min, Z.; Zhang, L.; Su, P.; Xiang, W. Mir-484 contributes to diminished ovarian reserve by regulating granulosa cell function via YAP1-mediated mitochondrial function and apoptosis. Int. J. Biol. Sci. 2022, 18, 1008–1021. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, S.M. MiR-484 suppressed proliferation, migration, invasion and induced apoptosis of gastric cancer via targeting CCL-18. Int. J. Exp. Pathol. 2020, 101, 203–214. [Google Scholar] [CrossRef]
- Rodríguez-Barrueco, R.; Latorre, J.; Devis-Jáuregui, L.; Lluch, A.; Bonifaci, N.; Llobet, F.J.; Olivan, M.; Coll-Iglesias, L.; Gassner, K.; Davis, M.L.; et al. A microRNA Cluster Controls Fat Cell Differentiation and Adipose Tissue Expansion by Regulating SNCG. Adv. Sci. 2022, 9, e2104759. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Khan, R.; Abbas Raza, S.H.; Suhail, S.M.; Khan, H.; Khan, S.B.; Abd El-Aziz, A.H.; Zan, L. RNA-Seq Reveals Function of Bta-miR-149-5p in the Regulation of Bovine Adipocyte Differentiation. Animals 2021, 11, 1207. [Google Scholar] [CrossRef]
- Juiz-Valiña, P.; Varela-Rodríguez, B.M.; Outeiriño-Blanco, E.; García-Brao, M.J.; Mena, E.; Cordido, F.; Sangiao-Alvarellos, S. MiR-19 Family Impairs Adipogenesis by the Downregulation of the PPARγ Transcriptional Network. Int. J. Mol. Sci. 2022, 13, 15792. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Lee, H.; Jung, C.H.; Jeon, T.I.; Ha, T.Y. MicroRNA-146b promotes adipogenesis by suppressing the SIRT1-FOXO1 cascade. EMBO Mol. Med. 2013, 5, 1602–1612. [Google Scholar] [CrossRef] [PubMed]
- Lagathu, C.; Christodoulides, C.; Tan, C.Y.; Virtue, S.; Laudes, M.; Campbell, M.; Ishikawa, K.; Ortega, F.; Tinahones, F.J.; Fernández-Real, J.M.; et al. Secreted frizzled-related protein 1 regulates adipose tissue expansion and is dysregulated in severe obesity. Int. J. Obes. 2010, 34, 1695–1705. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Uña, M.; Varela-Rey, M.; Cano, A.; Fernández-Ares, L.; Beraza, N.; Aurrekoetxea, I.; Martínez-Arranz, I.; García-Rodríguez, J.L.; Buqué, X.; Mestre, D.; et al. Excess S-adenosylmethionine reroutes phosphatidylethanolamine towards phosphatidylcholine and triglyceride synthesis. Hepatology 2013, 58, 1296–1305. [Google Scholar] [CrossRef] [Green Version]
- Ehrlund, A.; Mejhert, N.; Lorente-Cebrián, S.; Aström, G.; Dahlman, I.; Laurencikiene, J.; Rydén, M. Characterization of the Wnt Inhibitors Secreted Frizzled-Related Proteins (SFRPs) in Human Adipose Tissue. J. Clin. Endocrinol. Metab. 2013, 98, E503–E508. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Liu, L.; Su, Y.W.; Xian, C.J. miR-542-3p Attenuates Bone Loss and Marrow Adiposity Following Methotrexate Treatment by Targeting sFRP-1 and Smurf2. Int. J. Mol. Sci. 2021, 22, 10988. [Google Scholar] [CrossRef]
- Yang, M.; Gao, X.; Ma, Y.; Wang, X.; Lei, Z.; Wang, S.; Hu, H.; Tang, L.; Ma, Y. Bta-miR-6517 promotes proliferation and inhibits differentiation of pre-adipocytes by targeting PFKL. J. Anim. Physiol. Anim. Nutr. 2022, 106, 1197–1207. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Riffo-Campos, Á.L.; Riquelme, I.; Brebi-Mieville, P. Tools for Sequence-Based miRNA Target Prediction: What to Choose? Int. J. Mol. Sci. 2016, 9, 1987. [Google Scholar] [CrossRef] [Green Version]
- Pan, C.; Lei, Z.; Wang, S.; Wang, X.; Wei, D.; Cai, X.; Luoreng, Z.; Wang, L.; Ma, Y. Genome-wide identification of cyclin-dependent kinase (CDK) genes affecting adipocyte differentiation in cattle. BMC Genom. 2021, 22, 532. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.; Gao, X.; Hu, C.; Wang, S.; Sheng, H.; Ma, Y. Bta-miR-484 Targets SFRP1 and Affects Preadipocytes Proliferation, Differentiation, and Apoptosis. Int. J. Mol. Sci. 2023, 24, 12710. https://doi.org/10.3390/ijms241612710
Yang M, Gao X, Hu C, Wang S, Sheng H, Ma Y. Bta-miR-484 Targets SFRP1 and Affects Preadipocytes Proliferation, Differentiation, and Apoptosis. International Journal of Molecular Sciences. 2023; 24(16):12710. https://doi.org/10.3390/ijms241612710
Chicago/Turabian StyleYang, Mengli, Xiaoqian Gao, Chunli Hu, Shuzhe Wang, Hui Sheng, and Yun Ma. 2023. "Bta-miR-484 Targets SFRP1 and Affects Preadipocytes Proliferation, Differentiation, and Apoptosis" International Journal of Molecular Sciences 24, no. 16: 12710. https://doi.org/10.3390/ijms241612710