
The Contribution of TRPA1 to Corneal Thermosensitivity and Blink Regulation in Young and Aged Mice

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
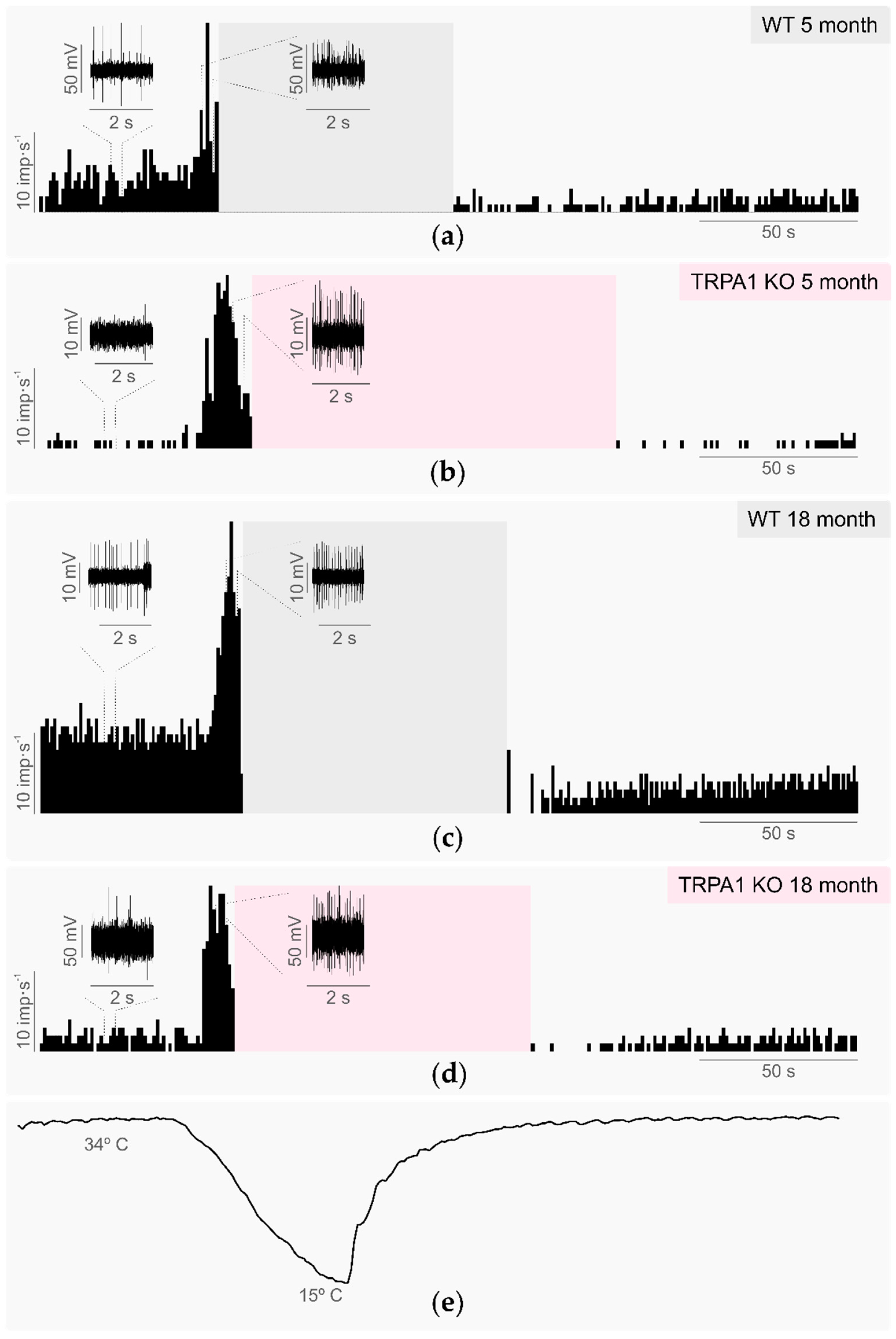
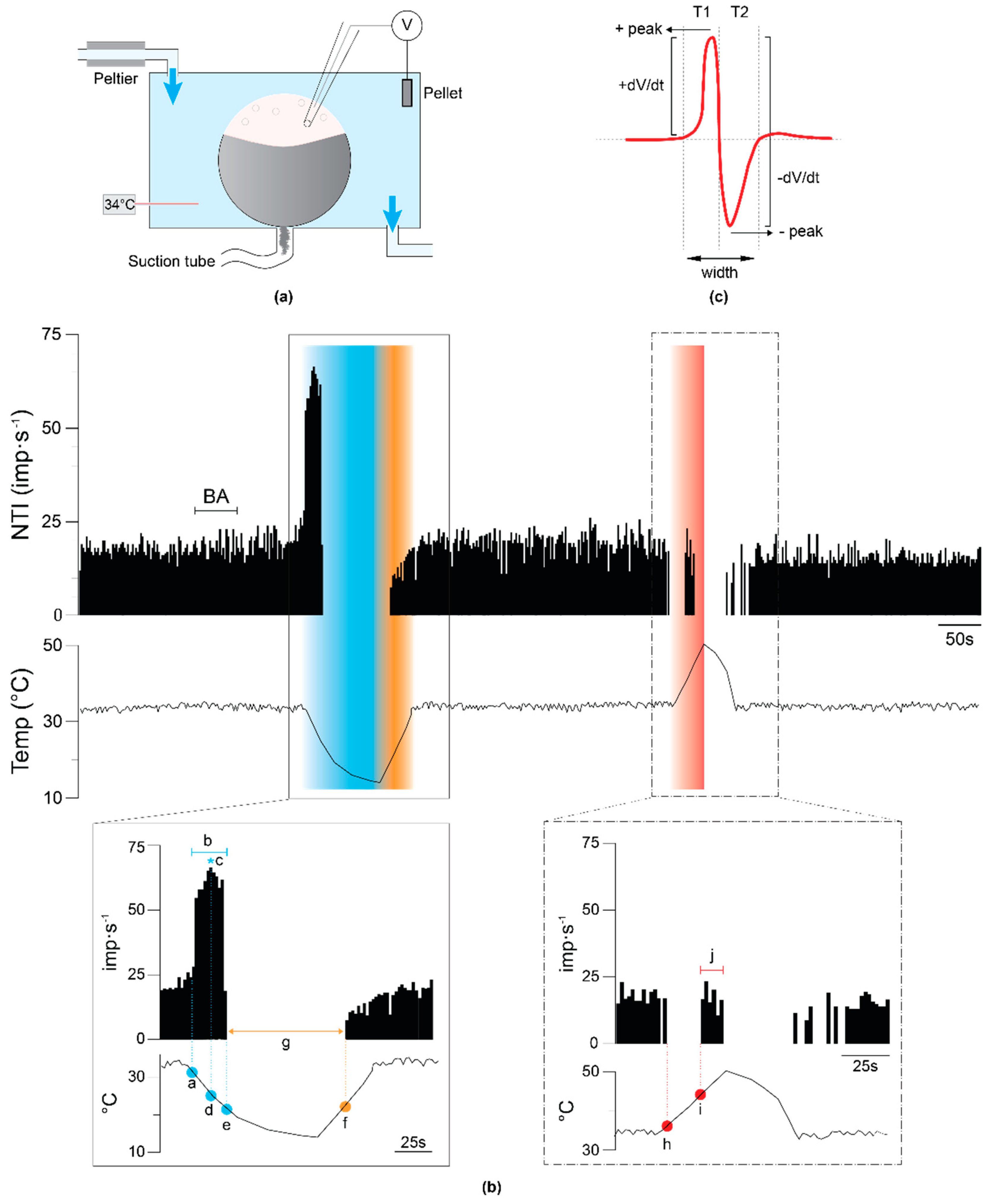
2.1. Corneal Cold Thermoreceptors’ Activity in WT and TRPA1-KO Young Mice
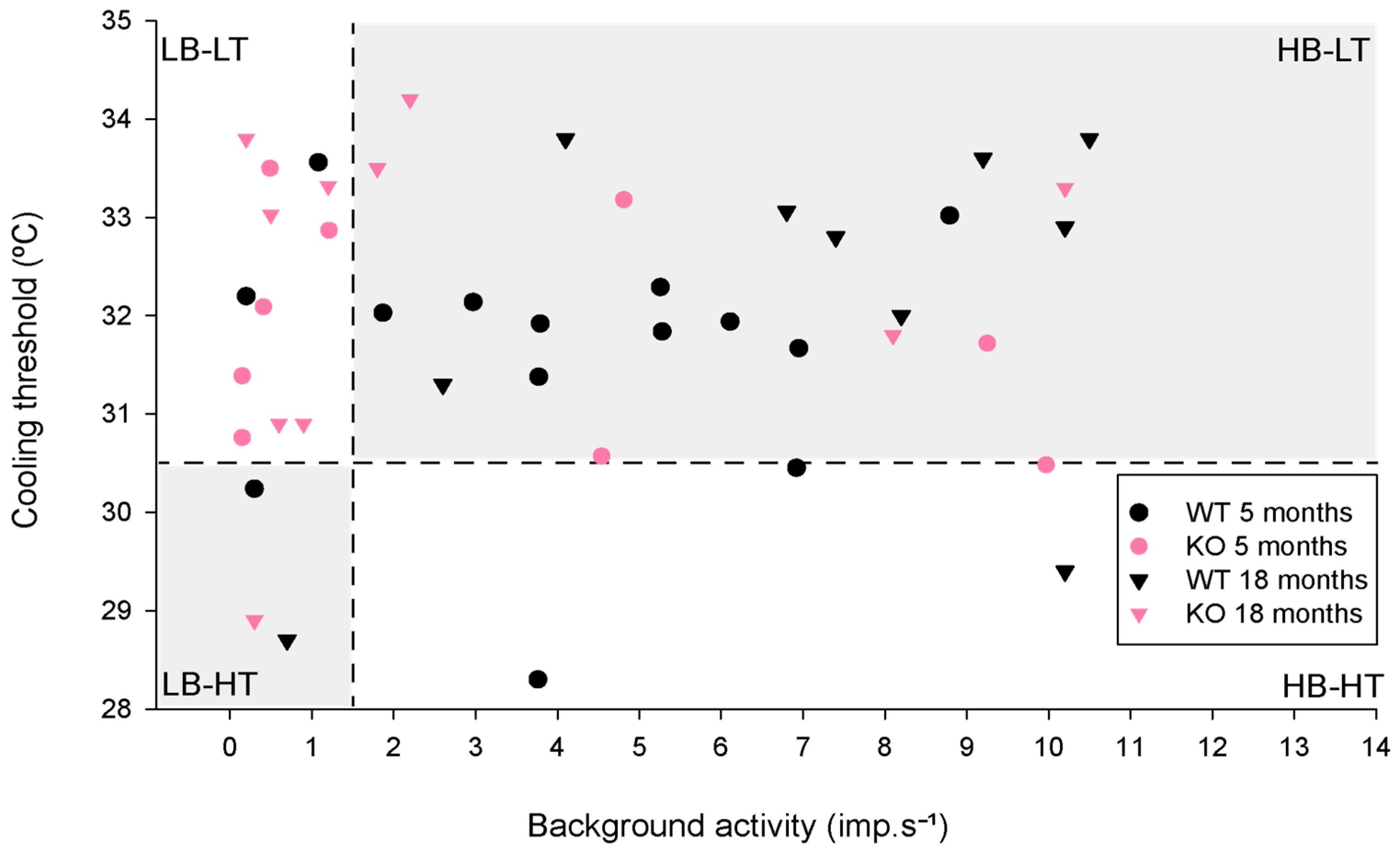
2.1.1. Background Activity and Cold-Evoked Responses
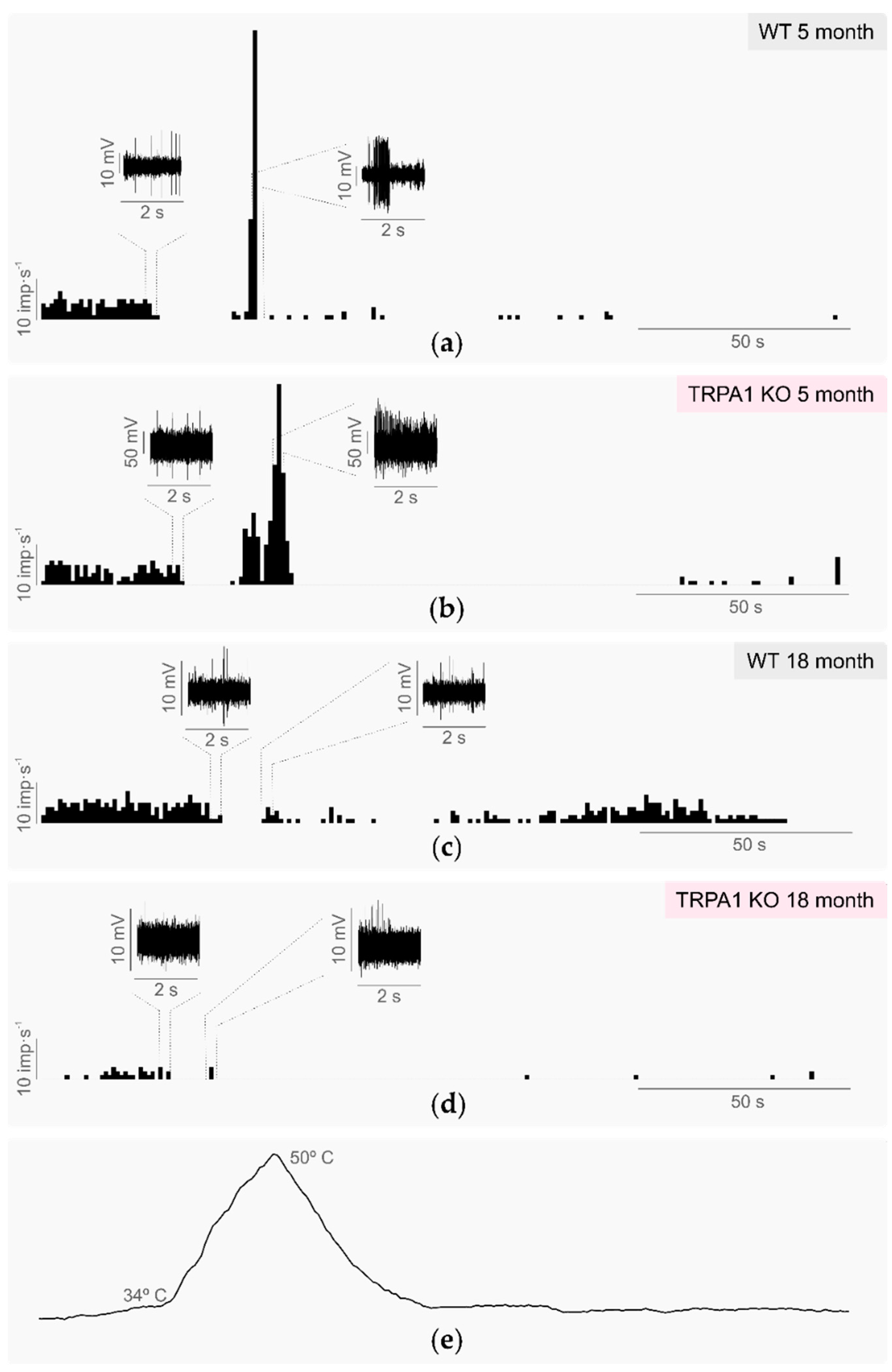
2.1.2. Sensitivity to Warm and Heat Stimuli
2.1.3. Shape of the Nerve Terminal Impulses
2.2. Tearing and Blinking in Young WT and TRPA1-KO Mice
2.2.1. Basal Tearing Volume
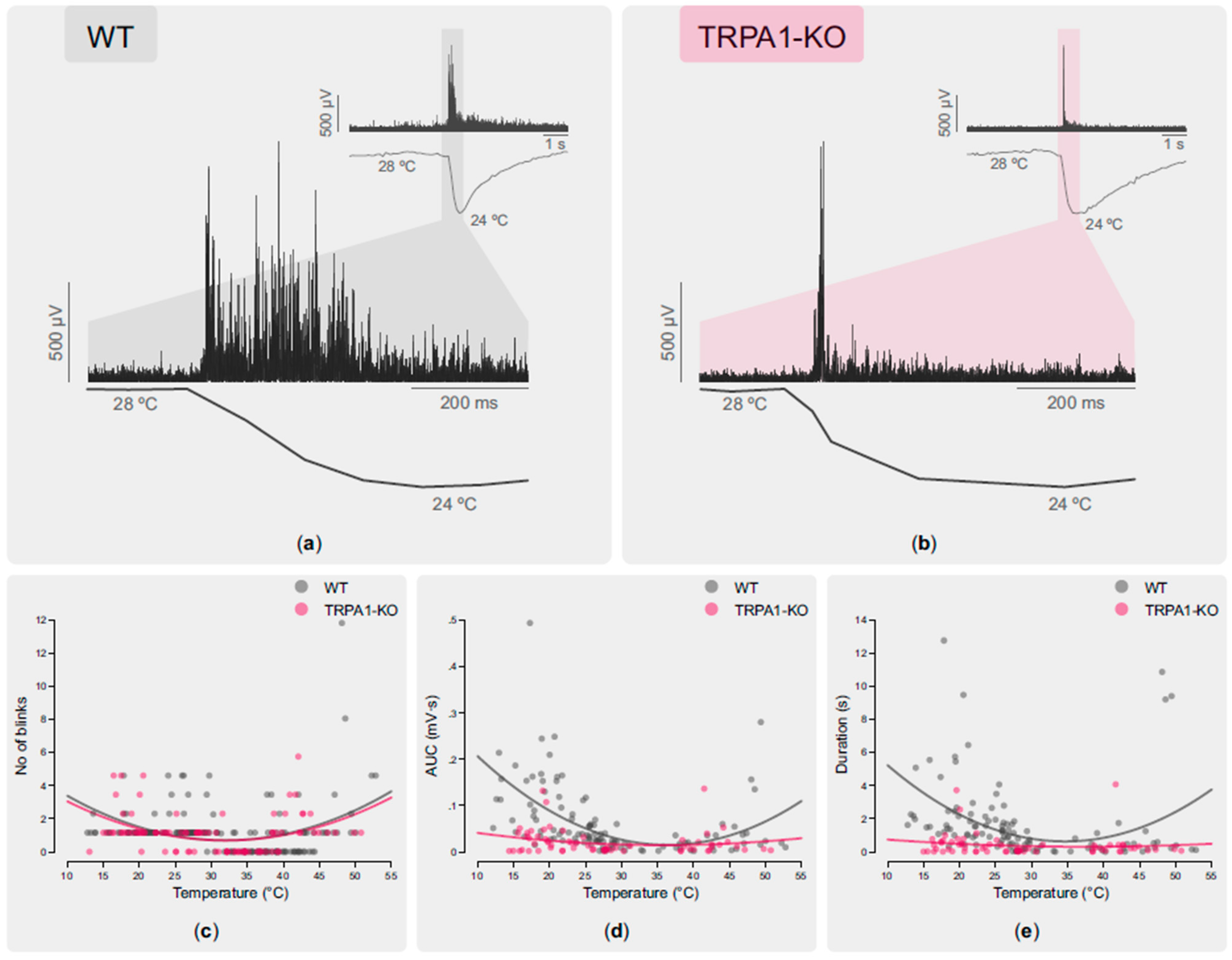
2.2.2. Blinking Evoked by Cooling and Heating Stimulation of the Ocular Surface
2.3. Effects of Aging in TRPA1-Dependent Thermosensitivity
2.3.1. Background and Stimulus-Evoked Activity of Cold Thermoreceptors
2.3.2. Shape of the Nerve Terminal Impulse
2.3.3. Basal Tearing Volume
3. Discussion
4. Methods
4.1. Animals
4.2. Electrophysiological Recording
4.2.1. Experimental Protocol
4.2.2. Analysis of the Electrophysiological Recordings
4.2.2.1. Background and Stimulus-Evoked NTI Activity
4.2.2.2. Shape of the NTIs
4.3. Measurement of the Basal Tearing Volume
4.4. Orbicularis Oculi Muscle Electromyography (OOemg)
4.4.1. Electrode Implantation
4.4.2. OOemg Recording
4.4.3. OOemg Analysis
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nilius, B.; Appendino, G.; Owsianik, G. The transient receptor potential channel TRPA1: From gene to pathophysiology. Pflugers Arch. Eur. J. Physiol. 2012, 464, 425–458. [Google Scholar] [CrossRef] [PubMed]
- Bandell, M.; Story, G.M.; Hwang, S.W.; Viswanath, V.; Eid, S.R.; Petrus, M.J.; Earley, T.J.; Patapoutian, A. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron 2004, 41, 849–857. [Google Scholar] [CrossRef] [Green Version]
- Sawada, Y.; Hosokawa, H.; Hori, A.; Matsumura, K.; Kobayashi, S. Cold sensitivity of recombinant TRPA1 channels. Brain Res. 2007, 1160, 39–46. [Google Scholar] [CrossRef]
- Latorre, R.; Brauchi, S.; Orta, G.; Zaelzer, C.; Vargas, G. ThermoTRP channels as modular proteins with allosteric gating. Cell Calcium 2007, 42, 427–438. [Google Scholar] [CrossRef]
- Karashima, Y.; Talavera, K.; Everaerts, W.; Janssens, A.; Kwan, K.Y.; Vennekens, R.; Nilius, B.; Voets, T. TRPA1 acts as a cold sensor in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2009, 106, 1273–1278. [Google Scholar] [CrossRef] [PubMed]
- Bautista, D.M.; Jordt, S.-E.; Nikai, T.; Tsuruda, P.R.; Read, A.J.; Poblete, J.; Yamoah, E.N.; Basbaum, A.I.; Julius, D. TRPA1 mediates the inflammatory actions of environmental irritants and proalgesic agents. Cell 2006, 124, 1269–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knowlton, W.M.; Bifolck-Fisher, A.; Bautista, D.M.; McKemy, D.D. TRPM8, but not TRPA1, is required for neural and behavioral responses to acute noxious cold temperatures and cold-mimetics in vivo. Pain 2010, 150, 340–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caspani, O.; Heppenstall, P. A TRPA1 and cold transduction: An unresolved issue? J. Gen. Physiol. 2009, 133, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Sinica, V.; Zimova, L.; Barvikova, K.; Macikova, L.; Barvik, I.; Vlachova, V. Human and Mouse TRPA1 Are Heat and Cold Sensors Differentially Tuned by Voltage. Cells 2019, 9, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moparthi, L.; Kichko, T.I.; Eberhardt, M.; Högestätt, E.D.; Kjellbom, P.; Johanson, U.; Reeh, P.W.; Leffler, A.; Filipovic, M.R.; Zygmunt, P.M. Human TRPA1 is a heat sensor displaying intrinsic U-shaped thermosensitivity. Sci. Rep. 2016, 6, 28763. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, T.; Kistner, K.; Miermeister, F.; Winkelmann, R.; Wittmann, J.; Fischer, M.J.M.; Weidner, C.; Reeh, P.W. TRPA1 and TRPV1 are differentially involved in heat nociception of mice. Eur. J. Pain 2013, 17, 1472–1482. [Google Scholar] [CrossRef]
- Vandewauw, I.; De Clercq, K.; Mulier, M.; Held, K.; Pinto, S.; Van Ranst, N.; Segal, A.; Voet, T.; Vennekens, R.; Zimmermann, K.; et al. A TRP channel trio mediates acute noxious heat sensing. Nature 2018, 555, 662–666. [Google Scholar] [CrossRef]
- Talavera, K.; Startek, J.B.; Alvarez-Collazo, J.; Boonen, B.; Alpizar, Y.A.; Sanchez, A.; Naert, R.; Nilius, B. Mammalian Transient Receptor Potential TRPA1 Channels: From Structure to Disease. Physiol. Rev. 2020, 100, 725–803. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Moreno, A.; Baudouin, C.; Melik Parsadaniantz, S.; Réaux-Le Goazigo, A. Morphological and Functional Changes of Corneal Nerves and Their Contribution to Peripheral and Central Sensory Abnormalities. Front. Cell. Neurosci. 2020, 14, 610342. [Google Scholar] [CrossRef]
- MacIver, M.B.; Tanelian, D.L. Structural and functional specialization of A delta and C fiber free nerve endings innervating rabbit corneal epithelium. J. Neurosci. 1993, 13, 4511–4524. [Google Scholar] [CrossRef]
- Gallar, J.; Pozo, M.A.; Tuckett, R.P.; Belmonte, C. Response of sensory units with unmyelinated fibres to mechanical, thermal and chemical stimulation of the cat’s cornea. J. Physiol. 1993, 468, 609–622. [Google Scholar] [CrossRef]
- Marfurt, C.F.; Cox, J.; Deek, S.; Dvorscak, L. Anatomy of the human corneal innervation. Exp. Eye Res. 2010, 90, 478–492. [Google Scholar] [CrossRef] [PubMed]
- Marfurt, C.F. The central projections of trigeminal primary afferent neurons in the cat as determined by the tranganglionic transport of horseradish peroxidase. J. Comp. Neurol. 1981, 203, 785–798. [Google Scholar] [CrossRef] [PubMed]
- Marfurt, C.F.; Del Toro, D.R. Corneal sensory pathway in the rat: A horseradish peroxidase tracing study. J. Comp. Neurol. 1987, 261, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Marfurt, C.F.; Echtenkamp, S.F. Central projections and trigeminal ganglion location of corneal afferent neurons in the monkey, Macaca fascicularis. J. Comp. Neurol. 1988, 272, 370–382. [Google Scholar] [CrossRef]
- Panneton, W.M.; Hsu, H.; Gan, Q. Distinct central representations for sensory fibers innervating either the conjunctiva or cornea of the rat. Exp. Eye Res. 2010, 90, 388–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belmonte, C.; Acosta, M.C.; Gallar, J. Neural basis of sensation in intact and injured corneas. Exp. Eye Res. 2004, 78, 513–525. [Google Scholar] [CrossRef]
- Belmonte, C.; Giraldez, F. Responses of cat corneal sensory receptors to mechanical and thermal stimulation. J. Physiol. 1981, 321, 355–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belmonte, C.; Gallar, J.; Pozo, M.A.; Rebollo, I. Excitation by irritant chemical substances of sensory afferent units in the cat’s cornea. J. Physiol. 1991, 437, 709–725. [Google Scholar] [CrossRef] [PubMed]
- Carr, R.W.; Pianova, S.; Fernandez, J.; Fallon, J.B.; Belmonte, C.; Brock, J.A. Effects of heating and cooling on nerve terminal impulses recorded from cold-sensitive receptors in the guinea-pig cornea. J. Gen. Physiol. 2003, 121, 427–439. [Google Scholar] [CrossRef]
- Parra, A.; Gonzalez-Gonzalez, O.; Gallar, J.; Belmonte, C. Tear fluid hyperosmolality increases nerve impulse activity of cold thermoreceptor endings of the cornea. Pain 2014, 155, 1481–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-González, O.; Bech, F.; Gallar, J.; Merayo-Lloves, J.; Belmonte, C. Functional properties of sensory nerve terminals of the mouse cornea. Investig. Ophthalmol. Vis. Sci. 2017, 58, 404–415. [Google Scholar] [CrossRef] [Green Version]
- Viana, F. Chemosensory properties of the trigeminal system. ACS Chem. Neurosci. 2011, 2, 38–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belmonte, C.; Gallar, J. Cold thermoreceptors, unexpected players in tear production and ocular dryness sensations. Investig. Ophthalmol. Vis. Sci. 2011, 52, 3888–3892. [Google Scholar] [CrossRef] [Green Version]
- Acosta, M.C.; Peral, A.; Luna, C.; Pintor, J.; Belmonte, C.; Gallar, J. Tear secretion induced by selective stimulation of corneal and conjunctival sensory nerve fibers. Investig. Ophthalmol. Vis. Sci. 2004, 45, 2333–2336. [Google Scholar] [CrossRef] [Green Version]
- Parra, A.; Madrid, R.; Echevarria, D.; del Olmo, S.; Morenilla-Palao, C.; Acosta, M.C.; Gallar, J.; Dhaka, A.; Viana, F.; Belmonte, C. Ocular surface wetness is regulated by TRPM8-dependent cold thermoreceptors of the cornea. Nat. Med. 2010, 16, 1396–1399. [Google Scholar] [CrossRef]
- Acosta, M.C.; Luna, C.; Quirce, S.; Belmonte, C.; Gallar, J. Corneal sensory nerve activity in an experimental model of UV keratitis. Investig. Ophthalmol. Vis. Sci. 2014, 55, 3403–3412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quallo, T.; Vastani, N.; Horridge, E.; Gentry, C.; Parra, A.; Moss, S.; Viana, F.; Belmonte, C.; Andersson, D.A.; Bevan, S. TRPM8 is a neuronal osmosensor that regulates eye blinking in mice. Nat. Commun. 2015, 6, 7150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKemy, D.D.; Neuhausser, W.M.; Julius, D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Peier, A.M.; Moqrich, A.; Hergarden, A.C.; Reeve, A.J.; Andersson, D.A.; Story, G.M.; Earley, T.J.; Dragoni, I.; McIntyre, P.; Bevan, S.; et al. A TRP channel that senses cold stimuli and menthol. Cell 2002, 108, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Acosta, M.C.; Luna, C.; Quirce, S.; Belmonte, C.; Gallar, J. Changes in sensory activity of ocular surface sensory nerves during allergic keratoconjunctivitis. Pain 2013, 154, 2353–2362. [Google Scholar] [CrossRef]
- Katagiri, A.; Thompson, R.; Rahman, M.; Okamoto, K.; Bereiter, D.A. Evidence for TRPA1 involvement in central neural mechanisms in a rat model of dry eye. Neuroscience 2015, 290, 204–213. [Google Scholar] [CrossRef] [Green Version]
- Guergova, S.; Dufour, A. Thermal sensitivity in the elderly: A review. Ageing Res. Rev. 2011, 10, 80–92. [Google Scholar] [CrossRef]
- Neira-Zalentein, W.; Holopainen, J.M.; Tervo, T.M.T.; Borrás, F.; Carmen Acosta, M.; Belmonte, C.; Gallar, J. Corneal sensitivity in diabetic patients subjected to retinal laser photocoagulation. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6043–6049. [Google Scholar] [CrossRef] [Green Version]
- Alcalde, I.; Íñigo-Portugués, A.; González-González, O.; Almaraz, L.; Artime, E.; Morenilla-Palao, C.; Gallar, J.; Viana, F.; Merayo-Lloves, J.; Belmonte, C. Morphological and functional changes in TRPM8-expressing corneal cold thermoreceptor neurons during aging and their impact on tearing in mice. J. Comp. Neurol. 2018, 526, 1859–1874. [Google Scholar] [CrossRef]
- Bech, F.; González-González, O.; Artime, E.; Serrano, J.; Alcalde, I.; Gallar, J.; Merayo-Lloves, J.; Belmonte, C. Functional and morphologic alterations in mechanical, polymodal, and cold sensory nerve fibers of the cornea following photorefractive keratectomy. Investig. Ophthalmol. Vis. Sci. 2018, 59, 2281–2292. [Google Scholar] [CrossRef] [Green Version]
- Carr, R.W.; Pianova, S.; Brock, J.A. The effects of polarizing current on nerve terminal impulses recorded from polymodal and cold receptors in the guinea-pig cornea. J. Gen. Physiol. 2002, 120, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Metzner, K.; Engel, P.; Balzulat, A.; Sisignano, M.; Ruth, P.; Lukowski, R.; Schmidtko, A.; Lu, R. Slack Potassium Channels Modulate TRPA1-Mediated Nociception in Sensory Neurons. Cells 2022, 11, 1693. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.O. Determinants of nerve terminal excitability. Neurol. Neurobiol. 1988, 35, 411–438. [Google Scholar]
- Brock, J.; Acosta, M.C.; Al Abed, A.; Pianova, S.; Belmonte, C. Barium ions inhibit the dynamic response of guinea-pig corneal cold receptors to heating but not to cooling. J. Physiol. 2006, 5752, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Bausch, A.E.; Kallenborn-Gerhardt, W.; Stoetzer, C.; Debruin, N.; Ruth, P.; Geisslinger, G.; Leffler, A.; Lukowski, R.; Schmidtko, A. Slack Channels Expressed in Sensory Neurons Control Neuropathic Pain in Mice. J. Neurosci. 2015, 35, 1125–1135. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Espinosa, P.L.; Wu, J.; Yang, C.; Gonzalez-Perez, V.; Zhou, H.; Liang, H.; Xia, X.M.; Lingle, C.J. Knockout of Slo2.2 enhances itch, abolishes KNa current, and increases action potential firing frequency in DRG neurons. eLife 2015, 4, e10013. [Google Scholar] [CrossRef]
- Lu, R.; Metzner, K.; Zhou, F.; Flauaus, C.; Balzulat, A.; Engel, P.; Petersen, J.; Ehinger, R.; Bausch, A.; Ruth, P.; et al. Functional Coupling of Slack Channels and P2X3 Receptors Contributes to Neuropathic Pain Processing. Int. J. Mol. Sci. 2021, 22, 405. [Google Scholar] [CrossRef] [PubMed]
- Kwan, K.Y.; Glazer, J.M.; Corey, D.P.; Rice, F.L.; Stucky, C.L. TRPA1 modulates mechanotransduction in cutaneous sensory neurons. J. Neurosci. 2009, 29, 4808–4819. [Google Scholar] [CrossRef] [Green Version]
- Paricio-Montesinos, R.; Schwaller, F.; Udhayachandran, A.; Rau, F.; Walcher, J.; Evangelista, R.; Vriens, J.; Voets, T.; Poulet, J.F.A.; Lewin, G.R. The Sensory Coding of Warm Perception. Neuron 2020, 106, 830–841.e3. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Wang, D.H. Knockout of Trpa1 accelerates age-related cardiac fibrosis and dysfunction. PLoS ONE 2022, 17, e0274618. [Google Scholar] [CrossRef] [PubMed]
- Delicado-Miralles, M.; Velasco, E.; Díaz-Tahoces, A.; Gallar, J.; Acosta, M.C.; Aracil-Marco, A. Deciphering the Action of Perfluorohexyloctane Eye Drops to Reduce Ocular Discomfort and Pain. Front. Med. 2021, 8, 709712. [Google Scholar] [CrossRef]
- Moparthi, L.; Sinica, V.; Moparthi, V.K.; Kreir, M.; Vignane, T.; Filipovic, M.R.; Vlachova, V.; Zygmunt, P.M. The human TRPA1 intrinsic cold and heat sensitivity involves separate channel structures beyond the N-ARD domain. Nat. Commun. 2022, 13, 6113. [Google Scholar] [CrossRef] [PubMed]
- Comes, N.; Gasull, X.; Callejo, G. Proton Sensing on the Ocular Surface: Implications in Eye Pain. Front. Pharmacol. 2021, 12, 773871. [Google Scholar] [CrossRef] [PubMed]
- Brock, J.A.; McLachlan, E.M.; Belmonte, C. Tetrodotoxin-resistant impulses in single nociceptor nerve terminals in guinea-pig cornea. J. Physiol. 1998, 512 Pt 1, 211–217. [Google Scholar] [CrossRef]
- Brock, J.A.; Pianova, S.; Belmonte, C. Differences between nerve terminal impulses of polymodal nociceptors and cold sensory receptors of the guinea-pig cornea. J. Physiol. 2001, 533, 493–501. [Google Scholar] [CrossRef]
- López-Ramos, J.C.; Houdek, Z.; Cendelín, J.; Vožeh, F.; Delgado-Garciá, J.M. Timing correlations between cerebellar interpositus neuronal firing and classically conditioned eyelid responses in wild-type and Lurcher mice. Sci. Rep. 2018, 8, 10697. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Young Mice | Aged Mice | |||
|---|---|---|---|---|
| WT n = 14 | TRPA1-KO n = 9 | WT n = 11 | TRPA1-KO n = 11 | |
| Basal temperature of 34 °C | 4.1 ± 0.7 | 3.4 ± 1.3 | 7.7 ± 1.2 | 4.5 ± 2.2 |
| Background activity (mean discharge rate; imp.s−1) | ||||
| Cooling from 34 °C to 15 °C | 31.6 ± 0.3 | 31.8 ± 0.4 | 32.2 ± 0.5 | 32.5 ± 0.5 |
| Cooling threshold (°C) | ||||
| Cooling response (mean discharge rate; imp.s−1) | 14.9 ± 2.5 | 19.8 ± 3.2 | 16.9 ± 2 | 13.0 ± 3.8 |
| Peak frequency (PF; imp.s−1) | 28.6 ± 4.9 | 40.3 ± 5.1 | 30.5 ± 3.3 | 25.8 ± 6.6 |
| Temperature at PF (°C) | 29 ± 0.9 | 29.7 ± 0.9 | 31.2 ± 0.8 | 30.5 ± 1.1 |
| Silencing temperature during the cooling ramp (°C) | 26.9 ± 0.9 | 27.7 ± 1.2 | 30 ± 1 | 28.6 ± 1.3 |
| Warming from 15 °C to 34 °C | 28.9 ± 0.7 | 33.2 ± 0.6 * | 32.4 ± 0.6 * | 31.9 ± 0.5 * |
| Return temperature (temperature to recover impulse activity; °C) | ||||
| Return time (time to recover impulse activity; s) | 74.2 ± 8.6 | 164.5 ± 23 * | 167.5 ± 16.5 * | 131.3 ± 15.2 * |
| Heating from 34 °C to 45 °C | 4/9 | 6/9 | 4/11 | 3/11 |
| Terminals fully silenced during the heating ramp (silenced terminals/n) | ||||
| Silencing temperature during the heating ramp (°C) | 35.4 ± 0.8 | 38.3 ± 1.5 | 39.9 ± 1.8 | 38.6 ± 2.4 |
| Terminals responding to heat (responding terminals/n) | 9/12 | 6/9 | 3/11 | 6/11 |
| Heating threshold (°C) | 41.1 ± 1.3 | 43.7 ± 0.8 | 44.7 ± 1.0 | 43.2 ± 1.9 |
| Response to heat (mean discharge rate; imp.s−1) | 11.5 ± 4.6 | 16.2 ± 6.3 | 10.2 ± 5.1 | 12.1 ± 3.4 |
| Young Mice | Aged Mice | |||
|---|---|---|---|---|
| WT n = 13 | TRPA1-KO n = 9 | WT n = 7 | TRPA1-KO n = 9 | |
| +dV/dt max (mV/ms) | 3.02 ± 0.14 | 3.15 ± 0.13 | 2.46 ± 0.25 | 2.79 ± 0.28 |
| −dV/dt max (mV/ms) | −3.25 ± 0.14 | −3.32 ± 0.19 | −2.83 ± 0.31 | −3.10 ± 0.19 |
| Ratio | −0.95 ± 0.06 | −0.97 ± 0.06 | −0.89 ± 0.06 | −0.93 ± 0.09 |
| Width (ms) | 1.5 ± 0.09 | 1.4 ± 0.07 | 1.8 ± 0.17 | 1.5 ± 0.07 |
| T1 (ms) | 0.5 ± 0.02 | 0.5 ± 0.02 | 0.7 ± 0.06 * | 0.6 ± 0.05 |
| T2 (ms) | 1.0 ± 0.09 | 0.9 ± 0.06 | 1.2 ± 0.1 | 0.8 ± 0.05 |
| WT | TRPA1-KO | |
|---|---|---|
| Cooling | ||
| No. of blinks | r =−0.206 p = 0.047 n = 94 | r = −0.484 p = 0.00005 n = 64 |
| OOemg AUC | r = −0.556 p = 0.00000003 n= 85 | r = −0.339 p = 0.023 n = 45 |
| OOemg duration | r = −0.457 p = 0.00001 n = 85 | r = −0.243 p = 0.108 n = 45 |
| Heating | ||
| No. of blinks | r = 0.517 p = 0.000002 n = 75 | r = 0.423 p = 0.014 n = 33 |
| OOemg AUC | r = 0.366 p = 0.061 n = 26 | r = −0.08 p = 0.747 n = 20 |
| OOemg duration | r = 0.324 p = 0.106 n = 27 | r = −0.09 p = 0.706 n = 20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frutos-Rincón, L.; Luna, C.; Aleixandre-Carrera, F.; Velasco, E.; Diaz-Tahoces, A.; Meseguer, V.; Gallar, J.; Acosta, M.C. The Contribution of TRPA1 to Corneal Thermosensitivity and Blink Regulation in Young and Aged Mice. Int. J. Mol. Sci. 2023, 24, 12620. https://doi.org/10.3390/ijms241612620
Frutos-Rincón L, Luna C, Aleixandre-Carrera F, Velasco E, Diaz-Tahoces A, Meseguer V, Gallar J, Acosta MC. The Contribution of TRPA1 to Corneal Thermosensitivity and Blink Regulation in Young and Aged Mice. International Journal of Molecular Sciences. 2023; 24(16):12620. https://doi.org/10.3390/ijms241612620
Chicago/Turabian StyleFrutos-Rincón, Laura, Carolina Luna, Fernando Aleixandre-Carrera, Enrique Velasco, Ariadna Diaz-Tahoces, Víctor Meseguer, Juana Gallar, and M. Carmen Acosta. 2023. "The Contribution of TRPA1 to Corneal Thermosensitivity and Blink Regulation in Young and Aged Mice" International Journal of Molecular Sciences 24, no. 16: 12620. https://doi.org/10.3390/ijms241612620