
The Role of Biological Rhythms in New Drug Formulations to Cross the Brain Barriers
 , and
, and
Abstract
:1. Introduction
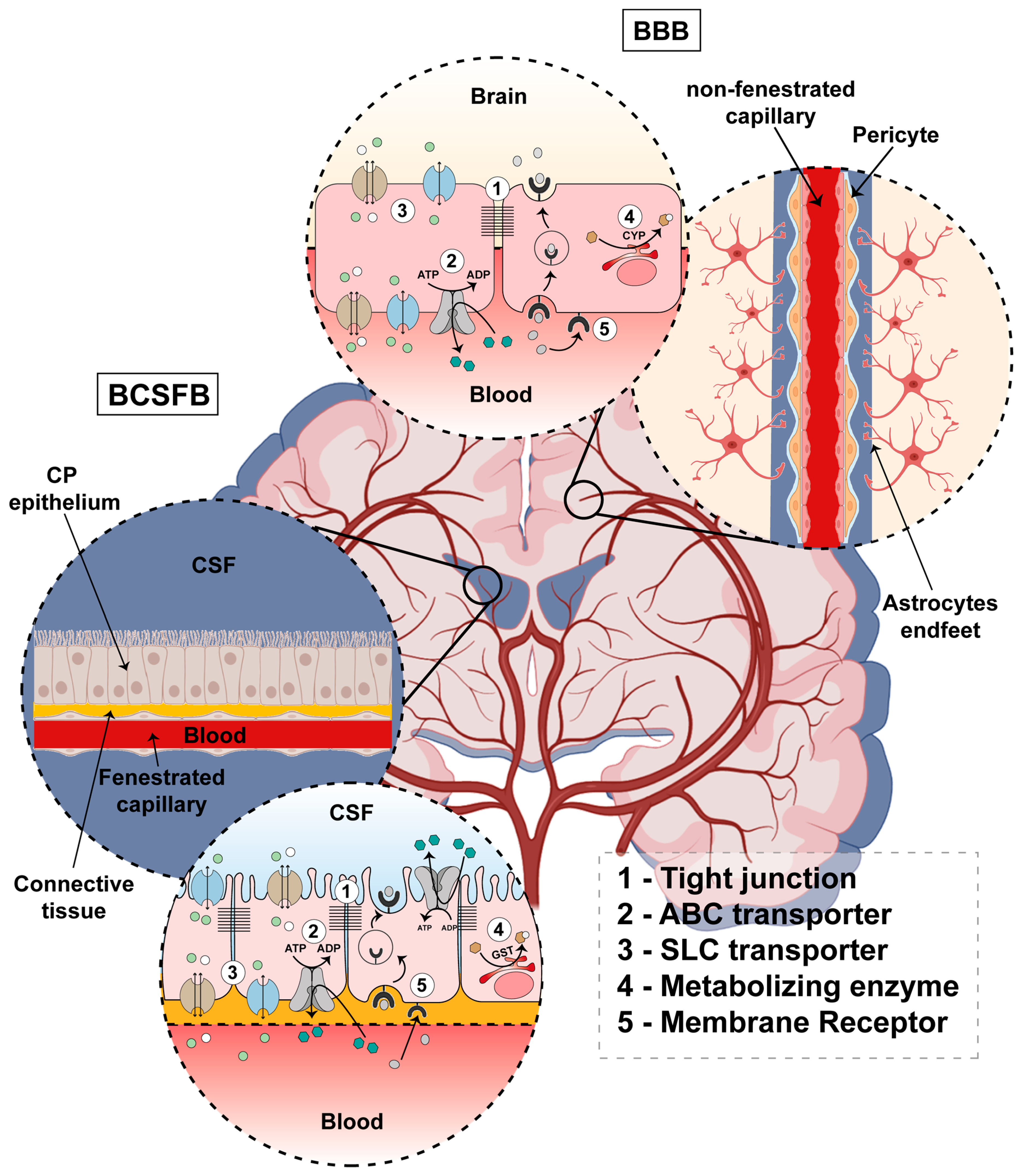
2. General Characterization of Brain Barriers
2.1. Blood–Brain Barrier
2.2. Blood–Cerebrospinal Fluid Barrier
3. Mechanisms of Transport in Brain Barriers
4. Circadian Rhythms in Brain Barriers
4.1. Circadian Rhythms in the Blood–Brain Barrier
4.2. Circadian Rhythms in the Blood–Cerebrospinal Fluid Barrier

{kind=link}
{kind=link}
| Topic | Animals/Cell Model | Results | References |
|---|---|---|---|
| Clock genes | Proestrus adult female and male Wistar rats | The choroid plexus molecular clock is dependent on sex. Cryptochrome (Cry) 2 and period (Per) 2 showed rhythmic expression in the choroid plexus of female and male rats. The expression of brain and muscle ARNT-like 1 (Bmal1) was only rhythmic in the choroid plexus of female rats. | [10] |
| Primary culture of rat choroid plexus epithelial cells | Bmal1, circadian locomotor output cycles kaput (Clock), and Per2 were shown to be rhythmically expressed in choroid plexus epithelial cells. For Per1 and Cry2, no significat rhythmic expression was found. | [10] | |
| Male Wistar rats | Bmal1 gene showed rhythmic expression in the rat choroid plexus. | [73] | |
| Per2::dLuc transgenic male rats on a Wistar rat background | At the lateral ventricle choroid plexus, Bmal1 expression was higher at zeitgeber time (ZT) 22 than ZT10, and the expression of Per1 and Per2 was higher at ZT10 than ZT4. At the fourth-ventricle choroid plexus, Bmal1 expression was higher at ZT 22 than ZT10, the expression of Per1 was higher at ZT10 than ZT4, and the expression of Per2 was higher at ZT16 than ZT4. | [85] | |
| Female Wistar rats | The abolishment of the rhythmic expression of Per2 and a phase advance of 6 h in Bmal1 expression were reported in the choroid plexus of ovariectomized rats. | [9] | |
| Primary culture of Wistar rat choroid plexus epithelial cells | The treatment of primary rat choroid plexus epithelial cells with 17β-estradiol (E2) led to the upregulation of the Bmal1, Per1, and Per2 genes. The E2 effect observed for Per1 and Per2 was mediated by the nuclear estrogen receptors. | [9] | |
| Rats | Bmal1 showed a higher expression in the choroid plexus of female rats when compared to male rats. Circadian-associated repressor of transcription (Ciart), albumin D-box binding protein (Dbp), Per2, and Per3 were shown to be more highly expressed in the choroid plexus of male rats. | [4,86] | |
| Wistar rats | Bmal1 was downregulated, and Ciart, Dbp, Per2, and Per3 were upregulated, in the choroid plexus of ovariectomized rats. Bmal1 was upregulated, and Ciart, Dbp, Per2, and Per3 were downregulated, in the choroid plexus of orchidectomized rats. | [4,87] | |
| Molecular clock robustness | Per2::Luc transgenic mice under C57BL/6J background | The molecular clock in the choroid plexus showed more robust rhythms than the suprachiasmatic nucleus molecular clock. The rhythms were evaluated by the Per2-driven luciferase rhythms of explants removed from Per2::Luc transgenic mice. | [88] |
| Cultured choroid plexuses of Per2::Luc transgenic mice | The choroid plexus showed higher single-cell synchrony in Per2-driven luciferase rhythms than in the suprachiasmatic nucleus. The treatment of the cultured choroid plexuses with a gap junction blocker dose dependently decreases the amplitude, increases the period, and decreases the cell synchrony in Per2-driven luciferase rhythms. | [88] | |
| Per2::Luc transgenic male rats | The intensity of rhythms in the CP, contrary to what happens in SCN, is not recovered after exposure to sevoflurane. | [89] | |
| Entrainment by glucocorticoids | Male Wistar rats | Per1 rhythmic expression was abolished and a decrease in the robustness of Per2, nuclear receptor subfamily 1 group D member 1 (Nd1r1), and Bmal1 rhythms in the choroid plexus was observed in adrenalectomized adult male Wistar rats. The treatment with dexamethasone upregulated Per1 expression and increased the amplitudes of rhythms in the expression of Bmal, Per2, and Nr1d1 in the male Wistar rat choroid plexus in adrenalectomized rats. | [84] |
| Per2::Luc mice choroid plexus explants | Dexamethasone influenced PER2 turnover in mouse choroid plexus explants. DEX treatment increased the amplitude and caused a phase shift in PER2-driven bioluminescence rhythms in mouse choroid plexus explants. The glucocorticoid receptor blocker mifepristone blocked the dexamethasone-induced phase shifts in PER2-driven bioluminescence rhythms in mouse choroid plexus explants. The induction of phase shifts in the choroid plexus clock is partially mediated by the activation of protein kinase A and the activation of the MAP kinase pathway. | [84] | |
| Amyloid β uptake, scavengers, and the influence on molecular clock | Human epithelial choroid plexus papilloma (HIBCPP) cell line | Amyloid β uptake was rhythmic in the human choroid plexus papilloma HIBCPP cell line. | [91] |
| Wistar Han Rats | Transthyretin showed rhythmic expression in the rat choroid plexus, with a peak during ZT16. Apolipoprotein J presented rhythmic expression in the choroid plexus of female rats, with a peak during the dark phase. The rhythmic expression of Apolipoprotein J was independent from a female sex-hormone background. | [94] | |
| APP/PS1 mice (Alzheimer’s disease mice model) | They compared the daily expression of amyloid β scavengers in 6- and 12-month APP/PS1 (Alzheimer’s disease mice model) and wild-type mice. Transthyretin showed a rhythmic expression in wild-type 12-month-old female mice. The angiotensin-converting enzyme was rhythmic in the choroid plexus of 6-month-old wild-type female mice. | [91] | |
| APP/PS1 mice (Alzheimer’s disease mice model) | Bmal1, Per2 and Cry2 were rhythmically expressed in the choroid plexus of 6- and 12-month-old APP/PS1 and wild-type mice. In female and male mice 12 months of age, Bmal1 was shown to be rhythmically expressed only in wild-type mice, but not in APP/PS1 mice. Cry2 only showed rhythmic expression in wild-type 6-month-old mice when compared with correspondent-aged APP/PS1 mice. In male mice 6 months of age, Per2 expression was rhythmic only in APP/PS1 mice, and not in wild-type mice. | [98] | |
| ATP-binding cassette (ABC) transporters | Wistar rats | Abcc4 was reported to be rhythmically expressed in the rat choroid plexus. Abcc1 and Abcg2 circadian expression in the choroid was found to be dependent on sex. Abcc1 showed rhythmic expression in the choroid plexus of male rats. Abcg2 showed rhythmic expression in the choroid plexus of female rats. | [12] |
| HIBCPP cell line | Abcc4 was shown to be rhythmically expressed in the HIBCPP cell line. ABCC4 may be partially responsible for the circadian transport of methotrexate across the HIBCPP cell line. | [12] | |
| Solute carrier (SLC) transporters | Wistar rats | The organic anion transporter 3 encoding gene, Slc22a8, was shown to be rhythmically expressed in the rat choroid plexus. | [12] |
| Melatonin production | Wistar rats | The aralkylamine N-acetyltransferase showed circadian expression in the rat choroid plexus. | [99] |
| Porcine CP explants | The melatonin secretion by porcine CP explant did not show a circadian pattern. | [99] |
5. Nanoformulations as Therapy for Central Nervous System Diseases
5.1. Polymeric NPs
5.2. Metallic NPS
5.3. Liposomes
5.4. Carbon Nanotubes
5.5. Exosomes
6. Circadian Rhythms in Nanoformulations for Central Nervous System Penetration
7. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Dou, X.; Meints, G.A.; Sedaghat-Herati, R. New Insights into the Interactions of a DNA Oligonucleotide with mPEGylated-PAMAM by Circular Dichroism and Solution NMR. J. Phys. Chem. B 2019, 123, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Howes, O.D.; Mehta, M.A. Challenges in CNS drug development and the role of imaging. Psychopharmacology 2021, 238, 1229–1230. [Google Scholar] [CrossRef] [PubMed]
- Daneman, R.; Prat, A. The blood-brain barrier. Cold Spring Harb. Perspect. Biol. 2015, 7, a020412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, C.R.; Duarte, A.C.; Quintela, T.; Tomás, J.; Albuquerque, T.; Marques, F.; Palha, J.A.; Gonçalves, I. The choroid plexus as a sex hormone target: Functional implications. Front. Neuroendocrinol. 2017, 44, 103–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhadra, U.; Thakkar, N.; Das, P.; Pal Bhadra, M. Evolution of circadian rhythms: From bacteria to human. Sleep Med. 2017, 35, 49–61. [Google Scholar] [CrossRef]
- Panda, S.; Hogenesch, J.B.; Kay, S.A. Circadian rhythms from flies to human. Nature 2002, 417, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2017, 18, 164–179. [Google Scholar] [CrossRef] [Green Version]
- Hastings, M.H.; Maywood, E.S.; Brancaccio, M. Generation of circadian rhythms in the suprachiasmatic nucleus. Nat. Rev. Neurosci. 2018, 19, 453–469. [Google Scholar] [CrossRef]
- Quintela, T.; Albuquerque, T.; Lundkvist, G.; Carmine Belin, A.; Talhada, D.; Gonçalves, I.; Carro, E.; Santos, C.R.A. The choroid plexus harbors a circadian oscillator modulated by estrogens. Chronobiol. Int. 2018, 35, 270–279. [Google Scholar] [CrossRef]
- Quintela, T.; Sousa, C.; Patriarca, F.M.; Gonçalves, I.; Santos, C.R. Gender associated circadian oscillations of the clock genes in rat choroid plexus. Brain Struct. Funct. 2015, 220, 1251–1562. [Google Scholar] [CrossRef]
- Zhang, S.L.; Lahens, N.F.; Yue, Z.; Arnold, D.M.; Pakstis, P.P.; Schwarz, J.E.; Sehgal, A. A circadian clock regulates efflux by the blood-brain barrier in mice and human cells. Nat. Commun. 2021, 12, 617. [Google Scholar] [CrossRef]
- Furtado, A.; Mineiro, R.; Duarte, A.C.; Gonçalves, I.; Santos, C.R.; Quintela, T. The Daily Expression of ABCC4 at the BCSFB Affects the Transport of Its Substrate Methotrexate. Int. J. Mol. Sci. 2022, 23, 2443. [Google Scholar] [CrossRef] [PubMed]
- Mudshinge, S.R.; Deore, A.B.; Patil, S.; Bhalgat, C.M. Nanoparticles: Emerging carriers for drug delivery. Saudi Pharm. J. 2011, 19, 129–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulgar, V.M. Transcytosis to Cross the Blood Brain Barrier, New Advancements and Challenges. Front. Neurosci. 2018, 12, 1019. [Google Scholar] [CrossRef]
- Fang, Z.; Chen, S.; Qin, J.; Chen, B.; Ni, G.; Chen, Z.; Zhou, J.; Li, Z.; Ning, Y.; Wu, C.; et al. Pluronic P85-coated poly(butylcyanoacrylate) nanoparticles overcome phenytoin resistance in P-glycoprotein overexpressing rats with lithium-pilocarpine-induced chronic temporal lobe epilepsy. Biomaterials 2016, 97, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Kadry, H.; Noorani, B.; Cucullo, L. A blood-brain barrier overview on structure, function, impairment, and biomarkers of integrity. Fluids Barriers CNS 2020, 17, 69. [Google Scholar] [CrossRef] [PubMed]
- Obermeier, B.; Verma, A.; Ransohoff, R.M. The blood-brain barrier. Handb. Clin. Neurol. 2016, 133, 39–59. [Google Scholar]
- Serlin, Y.; Shelef, I.; Knyazer, B.; Friedman, A. Anatomy and physiology of the blood-brain barrier. Semin. Cell Dev. Biol. 2015, 38, 2–6. [Google Scholar] [CrossRef] [Green Version]
- Abbott, N.J.; Rönnbäck, L.; Hansson, E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar] [CrossRef]
- Armulik, A.; Genové, G.; Mäe, M.; Nisancioglu, M.H.; Wallgard, E.; Niaudet, C.; He, L.; Norlin, J.; Lindblom, P.; Strittmatter, K.; et al. Pericytes regulate the blood-brain barrier. Nature 2010, 468, 557–561. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.S.; Foster, C.G.; Courtney, J.M.; King, N.E.; Howells, D.W.; Sutherland, B.A. Pericytes and Neurovascular Function in the Healthy and Diseased Brain. Front. Cell. Neurosci. 2019, 13, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krause, G.; Winkler, L.; Mueller, S.L.; Haseloff, R.F.; Piontek, J.; Blasig, I.E. Structure and function of claudins. Biochim. Biophys. Acta 2008, 1778, 631–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vermette, D.; Hu, P.; Canarie, M.F.; Funaro, M.; Glover, J.; Pierce, R.W. Tight junction structure, function, and assessment in the critically ill: A systematic review. Intensive Care Med. Exp. 2018, 6, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Günzel, D.; Yu, A.S. Claudins and the modulation of tight junction permeability. Physiol. Rev. 2013, 93, 525–569. [Google Scholar] [CrossRef] [Green Version]
- Berndt, P.; Winkler, L.; Cording, J.; Breitkreuz-Korff, O.; Rex, A.; Dithmer, S.; Rausch, V.; Blasig, R.; Richter, M.; Sporbert, A.; et al. Tight junction proteins at the blood-brain barrier: Far more than claudin-5. Cell. Mol. Life Sci. CMLS 2019, 76, 1987–2002. [Google Scholar] [CrossRef]
- Greene, C.; Hanley, N.; Campbell, M. Claudin-5: Gatekeeper of neurological function. Fluids Barriers CNS 2019, 16, 3. [Google Scholar] [CrossRef] [Green Version]
- Nitta, T.; Hata, M.; Gotoh, S.; Seo, Y.; Sasaki, H.; Hashimoto, N.; Furuse, M.; Tsukita, S. Size-selective loosening of the blood-brain barrier in claudin-5-deficient mice. J. Cell Biol. 2003, 161, 653–660. [Google Scholar] [CrossRef]
- Cooray, H.C.; Blackmore, C.G.; Maskell, L.; Barrand, M.A. Localisation of breast cancer resistance protein in microvessel endothelium of human brain. Neuroreport 2002, 13, 2059–2063. [Google Scholar] [CrossRef]
- Daood, M.; Tsai, C.; Ahdab-Barmada, M.; Watchko, J.F. ABC transporter (P-gp/ABCB1, MRP1/ABCC1, BCRP/ABCG2) expression in the developing human CNS. Neuropediatrics 2008, 39, 211–218. [Google Scholar] [CrossRef]
- Nies, A.T.; Jedlitschky, G.; König, J.; Herold-Mende, C.; Steiner, H.H.; Schmitt, H.P.; Keppler, D. Expression and immunolocalization of the multidrug resistance proteins, MRP1-MRP6 (ABCC1-ABCC6), in human brain. Neuroscience 2004, 129, 349–360. [Google Scholar] [CrossRef]
- Virgintino, D.; Robertson, D.; Errede, M.; Benagiano, V.; Girolamo, F.; Maiorano, E.; Roncali, L.; Bertossi, M. Expression of P-glycoprotein in human cerebral cortex microvessels. J. Histochem. Cytochem. 2002, 50, 1671–1676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dauchy, S.; Dutheil, F.; Weaver, R.J.; Chassoux, F.; Daumas-Duport, C.; Couraud, P.O.; Scherrmann, J.M.; De Waziers, I.; Declèves, X. ABC transporters, cytochromes P450 and their main transcription factors: Expression at the human blood-brain barrier. J. Neurochem. 2008, 107, 1518–1528. [Google Scholar] [CrossRef] [PubMed]
- Spoletini, I.; Vitale, C.; Malorni, W.; Rosano, G.M. Sex differences in drug effects: Interaction with sex hormones in adult life. Handb. Exp. Pharmacol. 2012, 214, 91–105. [Google Scholar]
- Sato, R.; Ohmori, K.; Umetsu, M.; Takao, M.; Tano, M.; Grant, G.; Porter, B.; Bet, A.; Terasaki, T.; Uchida, Y. An Atlas of the Quantitative Protein Expression of Anti-Epileptic-Drug Transporters, Metabolizing Enzymes and Tight Junctions at the Blood-Brain Barrier in Epileptic Patients. Pharmaceutics 2021, 13, 2122. [Google Scholar] [CrossRef]
- Zhao, Z.; Nelson, A.R.; Betsholtz, C.; Zlokovic, B.V. Establishment and Dysfunction of the Blood-Brain Barrier. Cell 2015, 163, 1064–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, B.W.; Gu, C. The molecular constituents of the blood-brain barrier. Trends Neurosci. 2015, 38, 598–608. [Google Scholar] [CrossRef] [Green Version]
- Ghersi-Egea, J.F.; Strazielle, N.; Catala, M.; Silva-Vargas, V.; Doetsch, F.; Engelhardt, B. Molecular anatomy and functions of the choroidal blood-cerebrospinal fluid barrier in health and disease. Acta Neuropathol. 2018, 135, 337–361. [Google Scholar] [CrossRef] [Green Version]
- Liddelow, S.A. Development of the choroid plexus and blood-CSF barrier. Front. Neurosci. 2015, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- Wolburg, H.; Paulus, W. Choroid plexus: Biology and pathology. Acta Neuropathol. 2010, 119, 75–88. [Google Scholar] [CrossRef]
- Praetorius, J.; Damkier, H.H. Transport across the choroid plexus epithelium. Am. J. Physiol. Cell Physiol. 2017, 312, C673–C686. [Google Scholar] [CrossRef]
- Wichmann, T.O.; Damkier, H.H.; Pedersen, M. A Brief Overview of the Cerebrospinal Fluid System and Its Implications for Brain and Spinal Cord Diseases. Front. Hum. Neurosci. 2021, 15, 737217. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, R.; Milatz, S.; Krug, S.M.; Oelrich, B.; Schulzke, J.D.; Amasheh, S.; Günzel, D.; Fromm, M. Claudin-2, a component of the tight junction, forms a paracellular water channel. J. Cell Sci. 2010, 123, 1913–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernd, A.; Ott, M.; Ishikawa, H.; Schroten, H.; Schwerk, C.; Fricker, G. Characterization of efflux transport proteins of the human choroid plexus papilloma cell line HIBCPP, a functional in vitro model of the blood-cerebrospinal fluid barrier. Pharm. Res. 2015, 32, 2973–2982. [Google Scholar] [CrossRef] [PubMed]
- Leggas, M.; Adachi, M.; Scheffer, G.L.; Sun, D.; Wielinga, P.; Du, G.; Mercer, K.E.; Zhuang, Y.; Panetta, J.C.; Johnston, B.; et al. Mrp4 confers resistance to topotecan and protects the brain from chemotherapy. Mol. Cell. Biol. 2004, 24, 7612–7621. [Google Scholar] [CrossRef] [Green Version]
- Rao, V.V.; Dahlheimer, J.L.; Bardgett, M.E.; Snyder, A.Z.; Finch, R.A.; Sartorelli, A.C.; Piwnica-Worms, D. Choroid plexus epithelial expression of MDR1 P glycoprotein and multidrug resistance-associated protein contribute to the blood-cerebrospinal-fluid drug-permeability barrier. Proc. Natl. Acad. Sci. USA 1999, 96, 3900–3905. [Google Scholar] [CrossRef]
- Tachikawa, M.; Watanabe, M.; Hori, S.; Fukaya, M.; Ohtsuki, S.; Asashima, T.; Terasaki, T. Distinct spatio-temporal expression of ABCA and ABCG transporters in the developing and adult mouse brain. J. Neurochem. 2005, 95, 294–304. [Google Scholar] [CrossRef]
- Ho, H.T.; Dahlin, A.; Wang, J. Expression Profiling of Solute Carrier Gene Families at the Blood-CSF Barrier. Front. Pharmacol. 2012, 3, 154. [Google Scholar] [CrossRef] [Green Version]
- Castañeyra-Ruiz, L.; González-Marrero, I.; Hernández-Abad, L.G.; Carmona-Calero, E.M.; Meyer, G.; Castañeyra-Perdomo, A. A Distal to Proximal Gradient of Human Choroid Plexus Development, with Antagonistic Expression of Glut1 and AQP1 in Mature Cells vs. Calbindin and PCNA in Proliferative Cells. Front. Neuroanat. 2016, 10, 87. [Google Scholar] [CrossRef] [Green Version]
- Damkier, H.H.; Nielsen, S.; Praetorius, J. Molecular expression of SLC4-derived Na+-dependent anion transporters in selected human tissues. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R2136–R2146. [Google Scholar] [CrossRef] [Green Version]
- Hinken, M.; Halwachs, S.; Kneuer, C.; Honscha, W. Subcellular localization and distribution of the reduced folate carrier in normal rat tissues. Eur. J. Histochem. EJH 2011, 55, e3. [Google Scholar] [CrossRef] [Green Version]
- Roberts, L.M.; Black, D.S.; Raman, C.; Woodford, K.; Zhou, M.; Haggerty, J.E.; Yan, A.T.; Cwirla, S.E.; Grindstaff, K.K. Subcellular localization of transporters along the rat blood-brain barrier and blood-cerebral-spinal fluid barrier by in vivo biotinylation. Neuroscience 2008, 155, 423–438. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.M.; Woodford, K.; Zhou, M.; Black, D.S.; Haggerty, J.E.; Tate, E.H.; Grindstaff, K.K.; Mengesha, W.; Raman, C.; Zerangue, N. Expression of the thyroid hormone transporters monocarboxylate transporter-8 (SLC16A2) and organic ion transporter-14 (SLCO1C1) at the blood-brain barrier. Endocrinology 2008, 149, 6251–6261. [Google Scholar] [CrossRef] [PubMed]
- Strazielle, N.; Ghersi-Egea, J.F. Potential Pathways for CNS Drug Delivery Across the Blood-Cerebrospinal Fluid Barrier. Curr. Pharm. Des. 2016, 22, 5463–5476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikitsh, J.L.; Chacko, A.M. Pathways for small molecule delivery to the central nervous system across the blood-brain barrier. Perspect. Med. Chem. 2014, 6, 11–24. [Google Scholar] [CrossRef] [Green Version]
- ter Beek, J.; Guskov, A.; Slotboom, D.J. Structural diversity of ABC transporters. J. Gen. Physiol. 2014, 143, 419–435. [Google Scholar] [CrossRef] [Green Version]
- Gil-Martins, E.; Barbosa, D.J.; Silva, V.; Remião, F.; Silva, R. Dysfunction of ABC transporters at the blood-brain barrier: Role in neurological disorders. Pharm. Ther. 2020, 213, 107554. [Google Scholar] [CrossRef]
- Lin, L.; Yee, S.W.; Kim, R.B.; Giacomini, K.M. SLC transporters as therapeutic targets: Emerging opportunities. Nat. Rev. Drug Discov. 2015, 14, 543–560. [Google Scholar] [CrossRef] [Green Version]
- Colas, C.; Ung, P.M.; Schlessinger, A. SLC Transporters: Structure, Function, and Drug Discovery. Medchemcomm 2016, 7, 1069–1081. [Google Scholar] [CrossRef] [Green Version]
- Dickens, D.; Webb, S.D.; Antonyuk, S.; Giannoudis, A.; Owen, A.; Rädisch, S.; Hasnain, S.S.; Pirmohamed, M. Transport of gabapentin by LAT1 (SLC7A5). Biochem. Pharmacol. 2013, 85, 1672–1683. [Google Scholar] [CrossRef]
- Kageyama, T.; Nakamura, M.; Matsuo, A.; Yamasaki, Y.; Takakura, Y.; Hashida, M.; Kanai, Y.; Naito, M.; Tsuruo, T.; Minato, N.; et al. The 4F2hc/LAT1 complex transports L-DOPA across the blood-brain barrier. Brain Res. 2000, 879, 115–121. [Google Scholar] [CrossRef]
- Duelli, R.; Enerson, B.E.; Gerhart, D.Z.; Drewes, L.R. Expression of large amino acid transporter LAT1 in rat brain endothelium. J. Cereb. Blood Flow Metab. 2000, 20, 1557–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.; Ecker, G.F. Insights into the Structure, Function, and Ligand Discovery of the Large Neutral Amino Acid Transporter 1, LAT1. Int. J. Mol. Sci. 2018, 19, 1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, M.E.; Rodriguez-Cruz, V.; Felmlee, M.A. SLC and ABC Transporters: Expression, Localization, and Species Differences at the Blood-Brain and the Blood-Cerebrospinal Fluid Barriers. AAPS J. 2017, 19, 1317–1331. [Google Scholar] [CrossRef] [PubMed]
- Stieger, B.; Gao, B. Drug transporters in the central nervous system. Clin. Pharmacokinet. 2015, 54, 225–242. [Google Scholar] [CrossRef] [Green Version]
- Puris, E.; Fricker, G.; Gynther, M. Targeting Transporters for Drug Delivery to the Brain: Can We Do Better? Pharm. Res. 2022, 39, 1415–1455. [Google Scholar] [CrossRef]
- Azarmi, M.; Maleki, H.; Nikkam, N.; Malekinejad, H. Transcellular brain drug delivery: A review on recent advancements. Int. J. Pharm. 2020, 586, 119582. [Google Scholar] [CrossRef]
- Grapp, M.; Wrede, A.; Schweizer, M.; Hüwel, S.; Galla, H.J.; Snaidero, N.; Simons, M.; Bückers, J.; Low, P.S.; Urlaub, H.; et al. Choroid plexus transcytosis and exosome shuttling deliver folate into brain parenchyma. Nat. Commun. 2013, 4, 2123. [Google Scholar] [CrossRef] [Green Version]
- Hervé, F.; Ghinea, N.; Scherrmann, J.M. CNS delivery via adsorptive transcytosis. AAPS J. 2008, 10, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Vorbrodt, A.W. Ultracytochemical characterization of anionic sites in the wall of brain capillaries. J. Neurocytol. 1989, 18, 359–368. [Google Scholar] [CrossRef]
- Zhu, X.; Jin, K.; Huang, Y.; Pang, Z. 7-Brain drug delivery by adsorption-mediated transcytosis. In Brain Targeted Drug Delivery System; Gao, H., Gao, X., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 159–183. [Google Scholar]
- Nakazato, R.; Kawabe, K.; Yamada, D.; Ikeno, S.; Mieda, M.; Shimba, S.; Hinoi, E.; Yoneda, Y.; Takarada, T. Disruption of Bmal1 Impairs Blood-Brain Barrier Integrity via Pericyte Dysfunction. J. Neurosci. 2017, 37, 10052–10062. [Google Scholar] [CrossRef] [Green Version]
- Hemmeryckx, B.; Frederix, L.; Lijnen, H.R. Deficiency of Bmal1 disrupts the diurnal rhythm of haemostasis. Exp. Gerontol. 2019, 118, 1–8. [Google Scholar] [CrossRef]
- Szczepkowska, A.; Harazin, A.; Barna, L.; Deli, M.A.; Skipor, J. Identification of Reference Genes for Circadian Studies on Brain Microvessels and Choroid Plexus Samples Isolated from Rats. Biomolecules 2021, 11, 1227. [Google Scholar] [CrossRef] [PubMed]
- Savolainen, H.; Meerlo, P.; Elsinga, P.H.; Windhorst, A.D.; Dierckx, R.A.; Colabufo, N.A.; van Waarde, A.; Luurtsema, G. P-glycoprotein Function in the Rodent Brain Displays a Daily Rhythm, a Quantitative In Vivo PET Study. AAPS J. 2016, 18, 1524–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulido, R.S.; Munji, R.N.; Chan, T.C.; Quirk, C.R.; Weiner, G.A.; Weger, B.D.; Rossi, M.J.; Elmsaouri, S.; Malfavon, M.; Deng, A.; et al. Neuronal Activity Regulates Blood-Brain Barrier Efflux Transport through Endothelial Circadian Genes. Neuron 2020, 108, 937–952.e7. [Google Scholar] [CrossRef]
- Ogata, S.; Ito, S.; Masuda, T.; Ohtsuki, S. Diurnal Changes in Protein Expression at the Blood-Brain Barrier in Mice. Biol. Pharm. Bull. 2022, 45, 751–756. [Google Scholar] [CrossRef]
- Zhang, S.L.; Yue, Z.; Arnold, D.M.; Artiushin, G.; Sehgal, A. A Circadian Clock in the Blood-Brain Barrier Regulates Xenobiotic Efflux. Cell 2018, 173, 130–139.e10. [Google Scholar] [CrossRef] [Green Version]
- Voog, L.; Eriksson, T. Relationship between plasma and brain large neutral amino acids in rats fed diets with different compositions at different times of the day. J. Neurochem. 1992, 59, 1868–1874. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, Y.; Zhang, X.Y.; Liu, X.; Liu, X.; Wu, X.; Jose, P.A.; Duan, S.; Xu, F.J.; Yang, Z. Intestinal Gastrin/CCKBR (Cholecystokinin B Receptor) Ameliorates Salt-Sensitive Hypertension by Inhibiting Intestinal Na(+)/H(+) Exchanger 3 Activity Through a PKC (Protein Kinase C)-Mediated NHERF1 and NHERF2 Pathway. Hypertension 2022, 79, 1668–1679. [Google Scholar] [CrossRef]
- Gómez-González, B.; Hurtado-Alvarado, G.; Esqueda-León, E.; Santana-Miranda, R.; Rojas-Zamorano, J.; Velázquez-Moctezuma, J. REM sleep loss and recovery regulates blood-brain barrier function. Curr. Neurovasc. Res. 2013, 10, 197–207. [Google Scholar] [CrossRef]
- Banks, W.A.; Kastin, A.J.; Ehrensing, C.A. Diurnal uptake of circulating interleukin-1alpha by brain, spinal cord, testis and muscle. Neuroimmunomodulation 1998, 5, 36–41. [Google Scholar] [CrossRef]
- Skinner, R.A.; Gibson, R.M.; Rothwell, N.J.; Pinteaux, E.; Penny, J.I. Transport of interleukin-1 across cerebromicrovascular endothelial cells. Br. J. Pharmacol. 2009, 156, 1115–1123. [Google Scholar] [CrossRef] [Green Version]
- Kress, G.J.; Liao, F.; Dimitry, J.; Cedeno, M.R.; FitzGerald, G.A.; Holtzman, D.M.; Musiek, E.S. Regulation of amyloid-β dynamics and pathology by the circadian clock. J. Exp. Med. 2018, 215, 1059–1068. [Google Scholar] [CrossRef] [Green Version]
- Liška, K.; Sládek, M.; Čečmanová, V.; Sumová, A. Glucocorticoids reset circadian clock in choroid plexus via period genes. J. Endocrinol. 2021, 248, 155–166. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Hamada, T.; Matsuzaki, T.; Iijima, N. Characterization of the circadian oscillator in the choroid plexus of rats. Biochem. Biophys. Res. Commun. 2020, 524, 497–501. [Google Scholar] [CrossRef]
- Quintela, T.; Marcelino, H.; Deery, M.J.; Feret, R.; Howard, J.; Lilley, K.S.; Albuquerque, T.; Gonçalves, I.; Duarte, A.C.; Santos, C.R. Sex-Related Differences in Rat Choroid Plexus and Cerebrospinal Fluid: A cDNA Microarray and Proteomic Analysis. J. Neuroendocrinol. 2016, 28. [Google Scholar] [CrossRef]
- Quintela, T.; Gonçalves, I.; Carreto, L.C.; Santos, M.A.; Marcelino, H.; Patriarca, F.M.; Santos, C.R. Analysis of the effects of sex hormone background on the rat choroid plexus transcriptome by cDNA microarrays. PLoS ONE 2013, 8, e60199. [Google Scholar] [CrossRef] [Green Version]
- Myung, J.; Schmal, C.; Hong, S.; Tsukizawa, Y.; Rose, P.; Zhang, Y.; Holtzman, M.J.; De Schutter, E.; Herzel, H.; Bordyugov, G.; et al. The choroid plexus is an important circadian clock component. Nat. Commun. 2018, 9, 1062. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, T.; Hamada, T.; Iijima, N. Differences in recovery processes of circadian oscillators in various tissues after sevoflurane treatment in vivo. Biochem. Biophys. Rep. 2022, 30, 101258. [Google Scholar] [CrossRef]
- Quintela, T.; Furtado, A.; Duarte, A.C.; Gonçalves, I.; Myung, J.; Santos, C.R.A. The role of circadian rhythm in choroid plexus functions. Prog. Neurobiol. 2021, 205, 102129. [Google Scholar] [CrossRef]
- Furtado, A.; Esgalhado, A.J.; Duarte, A.C.; Costa, A.R.; Costa-Brito, A.R.; Carro, E.; Ishikawa, H.; Schroten, H.; Schwerk, C.; Gonçalves, I.; et al. Circadian rhythmicity of amyloid-beta-related molecules is disrupted in the choroid plexus of a female Alzheimer’s disease mouse model. J. Neurosci. Res. 2023, 101, 524–540. [Google Scholar] [CrossRef]
- Oda, T.; Pasinetti, G.M.; Osterburg, H.H.; Anderson, C.; Johnson, S.A.; Finch, C.E. Purification and characterization of brain clusterin. Biochem. Biophys. Res. Commun. 1994, 204, 1131–1136. [Google Scholar] [CrossRef] [PubMed]
- Oda, T.; Wals, P.; Osterburg, H.H.; Johnson, S.A.; Pasinetti, G.M.; Morgan, T.E.; Rozovsky, I.; Stine, W.B.; Snyder, S.W.; Holzman, T.F.; et al. Clusterin (apoJ) alters the aggregation of amyloid beta-peptide (A beta 1–42) and forms slowly sedimenting A beta complexes that cause oxidative stress. Exp. Neurol. 1995, 136, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.C.; Furtado, A.; Hrynchak, M.V.; Costa, A.R.; Talhada, D.; Gonçalves, I.; Lemos, M.C.; Quintela, T.; Santos, C.R.A. Age, Sex Hormones, and Circadian Rhythm Regulate the Expression of Amyloid-Beta Scavengers at the Choroid Plexus. Int. J. Mol. Sci. 2020, 21, 6813. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.; Gonçalves, A.; Saraiva, M.J.; Cardoso, I. Transthyretin binding to A-Beta peptide--impact on A-Beta fibrillogenesis and toxicity. FEBS Lett. 2008, 582, 936–942. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, X.; Ladiwala, A.R.; Du, D.; Yadav, J.K.; Tessier, P.M.; Wright, P.E.; Kelly, J.W.; Buxbaum, J.N. Mechanisms of transthyretin inhibition of β-amyloid aggregation in vitro. J. Neurosci. 2013, 33, 19423–19433. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Kang, H.; Xu, Q.; Chen, M.J.; Liao, Y.; Thiyagarajan, M.; O’Donnell, J.; Christensen, D.J.; Nicholson, C.; Iliff, J.J.; et al. Sleep drives metabolite clearance from the adult brain. Science 2013, 342, 373–377. [Google Scholar] [CrossRef] [Green Version]
- Furtado, A.; Astaburuaga, R.; Costa, A.; Duarte, A.C.; Gonçalves, I.; Cipolla-Neto, J.; Lemos, M.C.; Carro, E.; Relógio, A.; Santos, C.R.A.; et al. The Rhythmicity of Clock Genes is Disrupted in the Choroid Plexus of the APP/PS1 Mouse Model of Alzheimer’s Disease. J. Alzheimer’s Dis. 2020, 77, 795–806. [Google Scholar] [CrossRef]
- Quintela, T.; Gonçalves, I.; Silva, M.; Duarte, A.C.; Guedes, P.; Andrade, K.; Freitas, F.; Talhada, D.; Albuquerque, T.; Tavares, S.; et al. Choroid plexus is an additional source of melatonin in the brain. J. Pineal Res. 2018, 65, e12528. [Google Scholar] [CrossRef]
- Lagaraine, C.; Skipor, J.; Szczepkowska, A.; Dufourny, L.; Thiery, J.C. Tight junction proteins vary in the choroid plexus of ewes according to photoperiod. Brain Res. 2011, 1393, 44–51. [Google Scholar] [CrossRef]
- Domínguez, A.; Suárez-Merino, B.; Goñi-de-Cerio, F. Nanoparticles and blood-brain barrier: The key to central nervous system diseases. J. Nanosci. Nanotechnol. 2014, 14, 766–779. [Google Scholar] [CrossRef]
- Borsook, D. Neurological diseases and pain. Brain 2012, 135 Pt 2, 320–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urquhart, B.L.; Kim, R.B. Blood-brain barrier transporters and response to CNS-active drugs. Eur. J. Clin. Pharmacol. 2009, 65, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Gribkoff, V.K.; Kaczmarek, L.K. The need for new approaches in CNS drug discovery: Why drugs have failed, and what can be done to improve outcomes. Neuropharmacology 2017, 120, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Jeevanandam, J.; Barhoum, A.; Chan, Y.S.; Dufresne, A.; Danquah, M.K. Review on nanoparticles and nanostructured materials: History, sources, toxicity and regulations. Beilstein J. Nanotechnol. 2018, 9, 1050–1074. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.; Nieh, M.P.; Li, Y. Decorating Nanoparticle Surface for Targeted Drug Delivery: Opportunities and Challenges. Polymers 2016, 8, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.X.; Wong, H.L.; Xue, H.Y.; Eoh, J.Y.; Wu, X.Y. Nanomedicine of synergistic drug combinations for cancer therapy-Strategies and perspectives. J. Control. Release 2016, 240, 489–503. [Google Scholar] [CrossRef] [Green Version]
- Ayub, A.; Wettig, S. An Overview of Nanotechnologies for Drug Delivery to the Brain. Pharmaceutics 2022, 14, 224. [Google Scholar] [CrossRef]
- Zhang, W.; Mehta, A.; Tong, Z.; Esser, L.; Voelcker, N.H. Development of Polymeric Nanoparticles for Blood–Brain Barrier Transfer—Strategies and Challenges. Adv. Sci. 2021, 8, 2003937. [Google Scholar] [CrossRef]
- Juhairiyah, F.; de Lange, E.C.M. Understanding Drug Delivery to the Brain Using Liposome-Based Strategies: Studies that Provide Mechanistic Insights Are Essential. AAPS J. 2021, 23, 114. [Google Scholar] [CrossRef]
- Azam, N.; Najabat Ali, M.; Javaid Khan, T. Carbon Quantum Dots for Biomedical Applications: Review and Analysis. Front. Mater. 2021, 8, 700403. [Google Scholar] [CrossRef]
- Ferreira Soares, D.C.; Domingues, S.C.; Viana, D.B.; Tebaldi, M.L. Polymer-hybrid nanoparticles: Current advances in biomedical applications. Biomed. Pharmacother. 2020, 131, 110695. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, A.; Carreiró, F.; Oliveira, A.M.; Neves, A.; Pires, B.; Venkatesh, D.N.; Durazzo, A.; Lucarini, M.; Eder, P.; Silva, A.M.; et al. Polymeric Nanoparticles: Production, Characterization, Toxicology and Ecotoxicology. Molecules 2020, 25, 3731. [Google Scholar] [CrossRef] [PubMed]
- La Barbera, L.; Mauri, E.; D’Amelio, M.; Gori, M. Functionalization strategies of polymeric nanoparticles for drug delivery in Alzheimer’s disease: Current trends and future perspectives. Front. Neurosci. 2022, 16, 939855. [Google Scholar] [CrossRef]
- Caraway, C.A.; Gaitsch, H.; Wicks, E.E.; Kalluri, A.; Kunadi, N.; Tyler, B.M. Polymeric Nanoparticles in Brain Cancer Therapy: A Review of Current Approaches. Polymers 2022, 14, 2963. [Google Scholar] [CrossRef] [PubMed]
- Sartaj, A.; Qamar, Z.; Md, S.; Alhakamy, N.A.; Baboota, S.; Ali, J. An Insight to Brain Targeting Utilizing Polymeric Nanoparticles: Effective Treatment Modalities for Neurological Disorders and Brain Tumor. Front. Bioeng. Biotechnol. 2022, 10, 788128. [Google Scholar]
- Islam, Y.; Leach, A.G.; Smith, J.; Pluchino, S.; Coxonl, C.R.; Sivakumaran, M.; Downing, J.; Fatokun, A.A.; Teixidò, M.; Ehtezazi, T. Peptide based drug delivery systems to the brain. Nano Express 2020, 1, 012002. [Google Scholar] [CrossRef]
- Hartl, N.; Adams, F.; Merkel, O.M. From Adsorption to Covalent Bonding: Apolipoprotein E Functionalization of Polymeric Nanoparticles for Drug Delivery Across the Blood–Brain Barrier. Adv. Ther. 2021, 4, 2000092. [Google Scholar] [CrossRef]
- Nabi, B.; Rehman, S.; Fazil, M.; Khan, S.; Baboota, S.; Ali, J. Riluzole-loaded nanoparticles to alleviate the symptoms of neurological disorders by attenuating oxidative stress. Drug Dev. Ind. Pharm. 2020, 46, 471–483. [Google Scholar] [CrossRef]
- Neves, A.R.; Queiroz, J.F.; Lima, S.A.C.; Reis, S. Apo E-Functionalization of Solid Lipid Nanoparticles Enhances Brain Drug Delivery: Uptake Mechanism and Transport Pathways. Bioconjug. Chem. 2017, 28, 995–1004. [Google Scholar] [CrossRef]
- Suk, J.S.; Xu, Q.; Kim, N.; Hanes, J.; Ensign, L.M. PEGylation as a strategy for improving nanoparticle-based drug and gene delivery. Adv. Drug Deliv. Rev. 2016, 99 Pt A, 28–51. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Xue, A.; Zhang, B.; Lou, H.; Shi, H.; Zhang, X. Polysorbate 80-coated PLGA nanoparticles improve the permeability of acetylpuerarin and enhance its brain-protective effects in rats. J. Pharm. Pharmacol. 2015, 67, 1650–1662. [Google Scholar] [CrossRef] [PubMed]
- Di Mauro, P.P.; Cascante, A.; Brugada Vilà, P.; Gómez-Vallejo, V.; Llop, J.; Borrós, S. Peptide-functionalized and high drug loaded novel nanoparticles as dual-targeting drug delivery system for modulated and controlled release of paclitaxel to brain glioma. Int. J. Pharm. 2018, 553, 169–185. [Google Scholar] [CrossRef] [PubMed]
- Gothwal, A.; Kumar, H.; Nakhate, K.T.; Ajazuddin; Dutta, A.; Borah, A.; Gupta, U. Lactoferrin Coupled Lower Generation PAMAM Dendrimers for Brain Targeted Delivery of Memantine in Aluminum-Chloride-Induced Alzheimer’s Disease in Mice. Bioconjug. Chem. 2019, 30, 2573–2583. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A. Chapter 1-Metal nanoparticles toxicity: Role of physicochemical aspects. In Metal Nanoparticles for Drug Delivery and Diagnostic Applications; Shah, M.R., Imran, M., Ullah, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–11. [Google Scholar]
- Piñón-Segundo, E.; Mendoza-Muñoz, N.; Quintanar-Guerrero, D. Chapter 23-Nanoparticles as Dental Drug-Delivery Systems. In Nanobiomaterials in Clinical Dentistry; Subramani, K., Ahmed, W., Hartsfield, J.K., Eds.; William Andrew Publishing: Norwich, NY, USA, 2013; pp. 475–495. [Google Scholar]
- Chandrakala, V.; Aruna, V.; Angajala, G. Review on metal nanoparticles as nanocarriers: Current challenges and perspectives in drug delivery systems. Emergent Mater. 2022, 5, 1593–1615. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liu, Z.W.; Allaker, R.P.; Reip, P.; Oxford, J.; Ahmad, Z.; Ren, G. A review of nanoparticle functionality and toxicity on the central nervous system. J. R. Soc. Interface 2010, 7 (Suppl. 4), S411–S422. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, B.; Su, L.; Nie, W.; Han, D.; Han, G.; Zhang, H.; Chong, C.; Tan, J. Subcellular distributions of iron oxide nanoparticles in rat brains affected by different surface modifications. J. Biomed. Mater. Res. Part A 2019, 107, 1988–1998. [Google Scholar] [CrossRef]
- Hersh, A.M.; Alomari, S.; Tyler, B.M. Crossing the Blood-Brain Barrier: Advances in Nanoparticle Technology for Drug Delivery in Neuro-Oncology. Int. J. Mol. Sci. 2022, 23, 4153. [Google Scholar] [CrossRef]
- Afzalipour, R.; Khoei, S.; Khoee, S.; Shirvalilou, S.; Raoufi, N.J.; Motevalian, M.; Karimi, M.Y. Thermosensitive magnetic nanoparticles exposed to alternating magnetic field and heat-mediated chemotherapy for an effective dual therapy in rat glioma model. Nanomed. Nanotechnol. Biol. Med. 2021, 31, 102319. [Google Scholar] [CrossRef]
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the clinic: An update. Bioeng. Transl. Med. 2019, 4, e10143. [Google Scholar] [CrossRef] [Green Version]
- Micheli, M.R.; Bova, R.; Magini, A.; Polidoro, M.; Emiliani, C. Lipid-based nanocarriers for CNS-targeted drug delivery. Recent Pat. CNS Drug Discov. 2012, 7, 71–86. [Google Scholar] [CrossRef]
- Amin, M.; Seynhaeve, A.L.B.; Sharifi, M.; Falahati, M.; ten Hagen, T.L.M. Liposomal Drug Delivery Systems for Cancer Therapy: The Rotterdam Experience. Pharmaceutics 2022, 14, 2165. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Chen, G.; Zhang, J. A Review of Liposomes as a Drug Delivery System: Current Status of Approved Products, Regulatory Environments, and Future Perspectives. Molecules 2022, 27, 1372. [Google Scholar] [CrossRef] [PubMed]
- Schnyder, A.; Huwyler, J. Drug transport to brain with targeted liposomes. NeuroRx 2005, 2, 99–107. [Google Scholar] [CrossRef]
- Johnsen, K.B.; Burkhart, A.; Melander, F.; Kempen, P.J.; Vejlebo, J.B.; Siupka, P.; Nielsen, M.S.; Andresen, T.L.; Moos, T. Targeting transferrin receptors at the blood-brain barrier improves the uptake of immunoliposomes and subsequent cargo transport into the brain parenchyma. Sci. Rep. 2017, 7, 10396. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, K.B.; Bak, M.; Melander, F.; Thomsen, M.S.; Burkhart, A.; Kempen, P.J.; Andresen, T.L.; Moos, T. Modulating the antibody density changes the uptake and transport at the blood-brain barrier of both transferrin receptor-targeted gold nanoparticles and liposomal cargo. J. Control. Release 2019, 295, 237–249. [Google Scholar] [CrossRef]
- Zhang, Z.; Guan, J.; Jiang, Z.; Yang, Y.; Liu, J.; Hua, W.; Mao, Y.; Li, C.; Lu, W.; Qian, J.; et al. Brain-targeted drug delivery by manipulating protein corona functions. Nat. Commun. 2019, 10, 3561. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Gaillard, P.J.; de Lange, E.C.M.; Hammarlund-Udenaes, M. Targeted brain delivery of methotrexate by glutathione PEGylated liposomes: How can the formulation make a difference? Eur. J. Pharm. Biopharm. 2019, 139, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.; Nayak, P.; Singh, G.; Khandai, M.; Sarangi, R.R.; Kar, M.K. Carbon Nanostructures as Therapeutic Cargoes: Recent Developments and Challenges. C 2023, 9, 3. [Google Scholar] [CrossRef]
- Dubey, R.; Dutta, D.; Sarkar, A.; Chattopadhyay, P. Functionalized carbon nanotubes: Synthesis, properties and applications in water purification, drug delivery, and material and biomedical sciences. Nanoscale Adv. 2021, 3, 5722–5744. [Google Scholar] [CrossRef]
- Zare, H.; Ahmadi, S.; Ghasemi, A.; Ghanbari, M.; Rabiee, N.; Bagherzadeh, M.; Karimi, M.; Webster, T.J.; Hamblin, M.R.; Mostafavi, E. Carbon Nanotubes: Smart Drug/Gene Delivery Carriers. Int. J. Nanomed. 2021, 16, 1681–1706. [Google Scholar] [CrossRef]
- Mohanta, D.; Patnaik, S.; Sood, S.; Das, N. Carbon nanotubes:Evaluation of toxicity at biointerfaces. J. Pharm. Anal. 2019, 9, 293. [Google Scholar] [CrossRef] [PubMed]
- Kafa, H.; Wang, J.T.; Rubio, N.; Venner, K.; Anderson, G.; Pach, E.; Ballesteros, B.; Preston, J.E.; Abbott, N.J.; Al-Jamal, K.T. The interaction of carbon nanotubes with an in vitro blood-brain barrier model and mouse brain in vivo. Biomaterials 2015, 53, 437–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kafa, H.; Wang, J.T.; Rubio, N.; Klippstein, R.; Costa, P.M.; Hassan, H.A.; Sosabowski, J.K.; Bansal, S.S.; Preston, J.E.; Abbott, N.J.; et al. Translocation of LRP1 targeted carbon nanotubes of different diameters across the blood-brain barrier in vitro and in vivo. J. Control. Release 2016, 225, 217–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, Y.; Wang, N.; He, L.; Shi, C.; Zhang, D.; Liu, Y.; Luo, L.; Chen, T. Designing dual-functionalized carbon nanotubes with high blood–brain-barrier permeability for precise orthotopic glioma therapy. Dalton Trans. 2019, 48, 1569–1573. [Google Scholar] [CrossRef]
- Harsha, P.J.; Thotakura, N.; Kumar, M.; Sharma, S.; Mittal, A.; Khurana, R.K.; Singh, B.; Negi, P.; Raza, K. A novel PEGylated carbon nanotube conjugated mangiferin: An explorative nanomedicine for brain cancer cells. J. Drug Deliv. Sci. Technol. 2019, 53, 101186. [Google Scholar] [CrossRef]
- Guo, Q.; You, H.; Yang, X.; Lin, B.; Zhu, Z.; Lu, Z.; Li, X.; Zhao, Y.; Mao, L.; Shen, S.; et al. Functional single-walled carbon nanotubes ‘CAR’ for targeting dopamine delivery into the brain of parkinsonian mice. Nanoscale 2017, 9, 10832–10845. [Google Scholar] [CrossRef]
- Qiao, R.; Jia, Q.; Hüwel, S.; Xia, R.; Liu, T.; Gao, F.; Galla, H.-J.; Gao, M. Receptor-Mediated Delivery of Magnetic Nanoparticles across the Blood–Brain Barrier. ACS Nano 2012, 6, 3304–3310. [Google Scholar] [CrossRef]
- Wang, J.; Chen, D.; Ho, E.A. Challenges in the development and establishment of exosome-based drug delivery systems. J. Control. Release 2021, 329, 894–906. [Google Scholar] [CrossRef]
- Yakubovich, E.I.; Polischouk, A.G.; Evtushenko, V.I. Principles and Problems of Exosome Isolation from Biological Fluids. Biochem. Suppl. Ser. A Membr. Cell Biol. 2022, 16, 115–126. [Google Scholar] [CrossRef]
- Heidarzadeh, M.; Gürsoy-Özdemir, Y.; Kaya, M.; Eslami Abriz, A.; Zarebkohan, A.; Rahbarghazi, R.; Sokullu, E. Exosomal delivery of therapeutic modulators through the blood–brain barrier; promise and pitfalls. Cell Biosci. 2021, 11, 142. [Google Scholar] [CrossRef]
- Matsumoto, J.; Stewart, T.; Banks, W.A.; Zhang, J. The Transport Mechanism of Extracellular Vesicles at the Blood-Brain Barrier. Curr. Pharm. Des. 2017, 23, 6206–6214. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xia, T.; Fang, Y.; Zuo, H.; Dong, X.; Xu, P.; Ouyang, J. Overcoming the blood–brain barrier by using a multistage exosome delivery system to inhibit central nervous system lymphoma. Nanomed. Nanotechnol. Biol. Med. 2022, 41, 102523. [Google Scholar] [CrossRef] [PubMed]
- Karami Fath, M.; Azami, J.; Masoudi, A.; Mosaddeghi Heris, R.; Rahmani, E.; Alavi, F.; Alagheband Bahrami, A.; Payandeh, Z.; Khalesi, B.; Dadkhah, M.; et al. Exosome-based strategies for diagnosis and therapy of glioma cancer. Cancer Cell Int. 2022, 22, 262. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Han, Y.; An, Y.; Ding, Y.; He, C.; Wang, X.; Tang, Q. NRP-1 targeted and cargo-loaded exosomes facilitate simultaneous imaging and therapy of glioma in vitro and in vivo. Biomaterials 2018, 178, 302–316. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Li, Y.; Ji, W.; Zhao, R.; Lu, Z.; Shen, J.; Wu, Y.; Wang, J.; Hao, Q.; Wang, J.; et al. Intranasal Administration of Self-Oriented Nanocarriers Based on Therapeutic Exosomes for Synergistic Treatment of Parkinson’s Disease. ACS Nano 2022, 16, 869–884. [Google Scholar] [CrossRef]
- Patel, A.; Olang, C.A.; Lewis, G.; Mandalaneni, K.; Anand, N.; Gorantla, V.R. An Overview of Parkinson’s Disease: Curcumin as a Possible Alternative Treatment. Cureus 2022, 14, e25032. [Google Scholar] [CrossRef]
- Copeland, C.; Stabenfeldt, S.E. Leveraging the Dynamic Blood-Brain Barrier for Central Nervous System Nanoparticle-based Drug Delivery Applications. Curr. Opin. Biomed. Eng. 2020, 14, 1–8. [Google Scholar] [CrossRef]
- Kreuter, J. Influence of chronobiology on the nanoparticle-mediated drug uptake into the brain. Pharmaceutics 2015, 7, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Carter, D.; Liu, X.; Tockary, T.A.; Dirisala, A.; Toh, K.; Anraku, Y.; Kataoka, K. Targeting nanoparticles to the brain by exploiting the blood-brain barrier impermeability to selectively label the brain endothelium. Proc. Natl. Acad. Sci. USA 2020, 117, 19141–19150. [Google Scholar] [CrossRef]
- Kudo, T.; Kawashima, M.; Tamagawa, T.; Shibata, S. Clock mutation facilitates accumulation of cholesterol in the liver of mice fed a cholesterol and/or cholic acid diet. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E120–E130. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.J.; Han, D.H.; Pak, Y.K.; Cho, S.H. Circadian regulation of low density lipoprotein receptor promoter activity by CLOCK/BMAL1, Hes1 and Hes6. Exp. Mol. Med. 2012, 44, 642–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okazaki, F.; Matsunaga, N.; Okazaki, H.; Azuma, H.; Hamamura, K.; Tsuruta, A.; Tsurudome, Y.; Ogino, T.; Hara, Y.; Suzuki, T.; et al. Circadian Clock in a Mouse Colon Tumor Regulates Intracellular Iron Levels to Promote Tumor Progression. J. Biol. Chem. 2016, 291, 7017–7028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Cortés, B.; Hurtado-Alvarado, G.; Martínez-Gómez, R.; León-Mercado, L.A.; Prager-Khoutorsky, M.; Buijs, R.M. Suprachiasmatic nucleus-mediated glucose entry into the arcuate nucleus determines the daily rhythm in blood glycemia. Curr. Biol. 2022, 32, 796–805.e4. [Google Scholar] [CrossRef] [PubMed]
- Milićević, N.; Ten Brink, J.B.; Ten Asbroek, A.; Bergen, A.A.; Felder-Schmittbuhl, M.P. The circadian clock regulates RPE-mediated lactate transport via SLC16A1 (MCT1). Exp. Eye Res. 2020, 190, 107861. [Google Scholar] [CrossRef]
- Soltésová, D.; Veselá, A.; Mravec, B.; Herichová, I. Daily profile of glut1 and glut4 expression in tissues inside and outside the blood-brain barrier in control and streptozotocin-treated rats. Physiol. Res. 2013, 62 (Suppl. 1), S115. [Google Scholar] [CrossRef]
- Vagnerová, K.; Ergang, P.; Soták, M.; Balounová, K.; Kvapilová, P.; Vodička, M.; Pácha, J. Diurnal expression of ABC and SLC transporters in jejunum is modulated by adrenalectomy. Comp. Biochem. Physiol. Toxicol. Pharmacol. 2019, 226, 108607. [Google Scholar] [CrossRef]
- Yuan, P.; Yang, T.; Mu, J.; Zhao, J.; Yang, Y.; Yan, Z.; Hou, Y.; Chen, C.; Xing, J.; Zhang, H.; et al. Circadian clock gene NPAS2 promotes reprogramming of glucose metabolism in hepatocellular carcinoma cells. Cancer Lett. 2020, 469, 498–509. [Google Scholar] [CrossRef]
- Min, H.S.; Kim, H.J.; Naito, M.; Ogura, S.; Toh, K.; Hayashi, K.; Kim, B.S.; Fukushima, S.; Anraku, Y.; Miyata, K.; et al. Systemic Brain Delivery of Antisense Oligonucleotides across the Blood–Brain Barrier with a Glucose-Coated Polymeric Nanocarrier. Angew. Chem. Int. Ed. 2020, 59, 8173–8180. [Google Scholar] [CrossRef] [Green Version]
- Anraku, Y.; Kuwahara, H.; Fukusato, Y.; Mizoguchi, A.; Ishii, T.; Nitta, K.; Matsumoto, Y.; Toh, K.; Miyata, K.; Uchida, S.; et al. Glycaemic control boosts glucosylated nanocarrier crossing the BBB into the brain. Nat. Commun. 2017, 8, 1001. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhao, Y.; Yue, Q.; Fu, Q.; Hai, L.; Guo, L.; Wang, Q.; Wu, Y. Preparation and Characterization of GLUT1-mediated Novel Brain Targeting Magnetic Nanoparticles. Lett. Drug Des. Discov. 2018, 15, 1308–1313. [Google Scholar] [CrossRef]
- Ak, G.; Ünal, A.; Karakayalı, T.; Özel, B.; Selvi Günel, N.; Hamarat Şanlıer, Ş. Brain-targeted, drug-loaded solid lipid nanoparticles against glioblastoma cells in culture. Colloids Surf. B Biointerfaces 2021, 206, 111946. [Google Scholar] [CrossRef]
- Ruan, S.; Qin, L.; Xiao, W.; Hu, C.; Zhou, Y.; Wang, R.; Sun, X.; Yu, W.; He, Q.; Gao, H. Acid-Responsive Transferrin Dissociation and GLUT Mediated Exocytosis for Increased Blood–Brain Barrier Transcytosis and Programmed Glioma Targeting Delivery. Adv. Funct. Mater. 2018, 28, 1802227. [Google Scholar] [CrossRef]
- Lam, F.C.; Morton, S.W.; Wyckoff, J.; Vu Han, T.-L.; Hwang, M.K.; Maffa, A.; Balkanska-Sinclair, E.; Yaffe, M.B.; Floyd, S.R.; Hammond, P.T. Enhanced efficacy of combined temozolomide and bromodomain inhibitor therapy for gliomas using targeted nanoparticles. Nat. Commun. 2018, 9, 1991. [Google Scholar] [CrossRef] [Green Version]
- Serna, N.; Céspedes, M.V.; Saccardo, P.; Xu, Z.; Unzueta, U.; Álamo, P.; Pesarrodona, M.; Sánchez-Chardi, A.; Roldán, M.; Mangues, R.; et al. Rational engineering of single-chain polypeptides into protein-only, BBB-targeted nanoparticles. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 1241–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, I.; Swami, R.; Jeengar, M.K.; Khan, W.; Sistla, R. p-Aminophenyl-α-d-mannopyranoside engineered lipidic nanoparticles for effective delivery of docetaxel to brain. Chem. Phys. Lipids 2015, 188, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, Y.; Lu, R.; Peng, Y.; Guo, L.; Hai, L.; Guan, M.; Wu, Y. Preparation and Characterization of Novel Brain Targeting Magnetic Nanoparticles Modified with Ascorbic Acid. Nano 2018, 13, 1850008. [Google Scholar] [CrossRef]
- Sun, R.; Liu, M.; Xu, Z.; Song, B.; He, Y.; Wang, H. Silicon-based nanoprobes cross the blood—Brain barrier for photothermal therapy of glioblastoma. Nano Res. 2022, 15, 7392–7401. [Google Scholar] [CrossRef]
- Venishetty, V.K.; Samala, R.; Komuravelli, R.; Kuncha, M.; Sistla, R.; Diwan, P.V. β-Hydroxybutyric acid grafted solid lipid nanoparticles: A novel strategy to improve drug delivery to brain. Nanomed. Nanotechnol. Biol. Med. 2013, 9, 388–397. [Google Scholar] [CrossRef]
- Neves, A.R.; Albuquerque, T.; Faria, R.; Gonçalves, A.M.; Santos, C.; Vivès, E.; Boisguérin, P.; Passarinha, L.A.; Sousa, Â.; Costa, D. Development of WRAP5 Peptide Complexes for Targeted Drug/Gene Co-Delivery toward Glioblastoma Therapy. Pharmaceutics 2022, 14, 2213. [Google Scholar] [CrossRef]
- de Castro, R.R.; do Carmo, F.A.; Martins, C.; Simon, A.; de Sousa, V.P.; Rodrigues, C.R.; Cabral, L.M.; Sarmento, B. Clofazimine functionalized polymeric nanoparticles for brain delivery in the tuberculosis treatment. Int. J. Pharm. 2021, 602, 120655. [Google Scholar] [CrossRef]
- Shen, Y.; Cao, B.; Snyder, N.R.; Woeppel, K.M.; Eles, J.R.; Cui, X.T. ROS responsive resveratrol delivery from LDLR peptide conjugated PLA-coated mesoporous silica nanoparticles across the blood–brain barrier. J. Nanobiotechnol. 2018, 16, 13. [Google Scholar] [CrossRef] [Green Version]
- Seo, S.; Kim, E.H.; Chang, W.S.; Lee, W.S.; Kim, K.H.; Kim, J.K. Enhanced proton treatment with a LDLR-ligand peptide-conjugated gold nanoparticles targeting the tumor microenvironment in an infiltrative brain tumor model. Am. J. Cancer Res. 2022, 12, 198–209. [Google Scholar] [PubMed]
- Portioli, C.; Bovi, M.; Benati, D.; Donini, M.; Perduca, M.; Romeo, A.; Dusi, S.; Monaco, H.L.; Bentivoglio, M. Novel functionalization strategies of polymeric nanoparticles as carriers for brain medications. J. Biomed. Mater. Res. Part A 2017, 105, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, T.; Neves, A.R.; Quintela, T.; Costa, D. The Influence of Circadian Rhythm on Cancer Cells Targeting and Transfection Efficiency of a Polycation-Drug/Gene Delivery Vector. Polymers 2022, 14, 681. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, L.S.; Álvaro, A.R.; Carmo-Silva, S.; Mendes, A.F.; Relógio, A.; Cavadas, C. The importance of determining circadian parameters in pharmacological studies. Br. J. Pharmacol. 2019, 176, 2827–2847. [Google Scholar] [CrossRef] [PubMed]


| Topic | Species/Cell Line | Results | References |
|---|---|---|---|
| Clock genes | C57BL/6J mice background (Bmal1fl/fl (control) and Bmal1fl/fl; Tie2: Cre) | The following genes were rhythmically expressed in mice brain endothelial cells: brain and muscle ARNT-like 1 (Bmal1), nuclear receptor subfamily 1 group D member 1 (Nr1d1), period (Per) 2, albumin D-box binding protein (Dbp), hepatic leukemia factor (Hlf), and thyrotroph embryonic factor (Tef). The rhythmicity of these genes was lost in mice with an endothelial-specific Bmal1 deletion. | [11] |
| Male Wistar rats | Bmal was showed to be rhythmically expressed in the brain microvessels of Wistar rats. | [73] | |
| hCMED/D3 cell line | Bmal1 gene transcript showed a circadian rhythm in the human brain microvascular endothelial cell line hCMED/D3. | [11] | |
| Blood–brain barrier (BBB) permeability | C57BL/6J mice background (Bmal1fl/fl (control) and Bmal1fl/fl; Nestin: Cre) | BBB permeability to Evans blue and biotin was increased in brain Bmal1 knock-out mice when compared to their littermate controls. | [71] |
| C57BL/6J mice | BBB permeability to Evans blue was not significantly different from Bmal1 knock-out mice or their littermate controls. | [72] | |
| ATP-binding cassette (ABC) transporters | Male Sprague–Dawley rats | Wistar rats showed higher brain permeability to the ABCB1 substrate MC225 at Zeitgeber time (ZT) 15. | [74] |
| C57BL/6J mice background (Bmal1fl/fl (control) and Bmal1fl/fl; VECadherinERT2: CRE) | The deletion of Bmal1 in mice endothelial cells resulted in the abrogation of the circadian oscillations of ABCB1-mediated Rhodamine123 brain efflux. | [75] | |
| C57BL/6J mice background (Bmal1fl/fl (control) and Bmal1fl/fl; Tie2: Cre) | The deletion of endothelial Bmal1 resulted in the loss of the rhythmic oscillation in Rhodamine B (substrate of ABCB1) brain efflux. However, the expression of the two rodent abcb1 genes (abcb1a and abcb1b) was not rhythmic either in the control mice or the mice with a specific deletion for endothelial Bmal1. | [11] | |
| C57BL/6J mice background (Bmal1fl/fl (control) and Bmal1fl/fl; Tie2: Cre) | Among various tested members of the ABC family, a microarray study reported that the Abcg2 gene was the only one presenting circadian oscillations in brain endothelial cells. The circadian oscillation of Abcg2 was not dependent on Bmal1. | [11] | |
| C57BL/6N male mice | A quantitative proteomic analysis showed that the protein expression of ABCB1 (a and b), ABCC4, ABCC9, and ABCG2 in mouse brain capillaries was not significantly different between ZT 0 and ZT 6, 12, and 18. | [76] | |
| hCMED/D3 cell line | Abcb1 gene transcript and protein expression were not rhythmic in the human brain microvascular endothelial cell line hCMED/D3. | [11] | |
| Solute carrier (SLC) transporters | C57BL/6J mice background (Bmal1fl/fl (control) and Bmal1fl/fl; Tie2: Cre) | Slc7a5, which encodes for the large neutral amino acid transporter 1 small subunit 1, is rhythmically expressed in mouse brain endothelial cells. The rhythmic expression of Slc7a5 is independent of Bmal1, since the expression was rhythmic either in brain endothelial cells from control mice or mice negative for endothelial Bmal1. | [11] |
| C57BL/6N male mice | Among various SLC proteins tested, a quantitative proteomic analysis only showed differences for SLC9A3R2 expression between ZT0 and ZT6, 12, and 18. | [76] | |
| Membrane receptors | C57BL/6N male mice | A quantitative proteomic analysis showed that the protein expression of insulin and transferrin receptors did not present any significant differences between ZT0 and ZT6, 12, and 18. | [76] |
| Tight junctions | C57BL/6N male mice | A proteomic analysis showed no significant differences in protein expression for claudin 5, occludin, and tight junction protein 1 between ZT0 and ZT6, 12, and 18. | [76] |
| Nanoformulation | Circadian-Expressed Receptor | Results | References |
|---|---|---|---|
| Docetaxel (DTX)-loaded solid lipidic nanoparticle surface modified with mannose | GLUT1 |
| [178] |
| Ibuprofen-loaded brain-targeting magnetic nanoparticles (AA-Ibu-PEG-DA@MNPs) modified with ascorbic acid (AA) |
| [179] | |
| SiNPs-based nanoprobes bearing the ligand of glucosamine (G) and indocyanine green (ICG) |
| [180] | |
| Betreliesoxybutyric acid (HBA) grafted docetaxel-loaded solid lipid nanoparticles (HD-SLNs) | MCT1 |
| [181] |
| Cell-penetrating peptide (WRAP5) bearing Tf ligand and loading p53 plasmid | TfR |
| [182] |
| Clofazimine-loaded PLGA-PEG nanoparticles (NP-CFZ) functionalized with the Tf ligand |
| [183] | |
| Polylactic acid (PLA)-coated mesoporous silica nanoparticles (MSNPs) conjugated with a ligand peptide of LDLR for resveratrol delivery | LDLR |
| [184] |
| LDLR ligand-functionalized gold nanoparticles (ApoB@AuNPs) |
| [185] | |
| PLGA nanoparticles functionalized with an apolipoprotein E-modified peptide (pep-apoE) or with lipocalin-type prostaglandin-d-synthase (L-PGDS) |
| [186] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mineiro, R.; Albuquerque, T.; Neves, A.R.; Santos, C.R.A.; Costa, D.; Quintela, T. The Role of Biological Rhythms in New Drug Formulations to Cross the Brain Barriers. Int. J. Mol. Sci. 2023, 24, 12541. https://doi.org/10.3390/ijms241612541
Mineiro R, Albuquerque T, Neves AR, Santos CRA, Costa D, Quintela T. The Role of Biological Rhythms in New Drug Formulations to Cross the Brain Barriers. International Journal of Molecular Sciences. 2023; 24(16):12541. https://doi.org/10.3390/ijms241612541
Chicago/Turabian StyleMineiro, Rafael, Tânia Albuquerque, Ana Raquel Neves, Cecília R. A. Santos, Diana Costa, and Telma Quintela. 2023. "The Role of Biological Rhythms in New Drug Formulations to Cross the Brain Barriers" International Journal of Molecular Sciences 24, no. 16: 12541. https://doi.org/10.3390/ijms241612541