

Establishment and Characterization of Mild Atopic Dermatitis in the DNCB-Induced Mouse Model

 , ,
, ,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
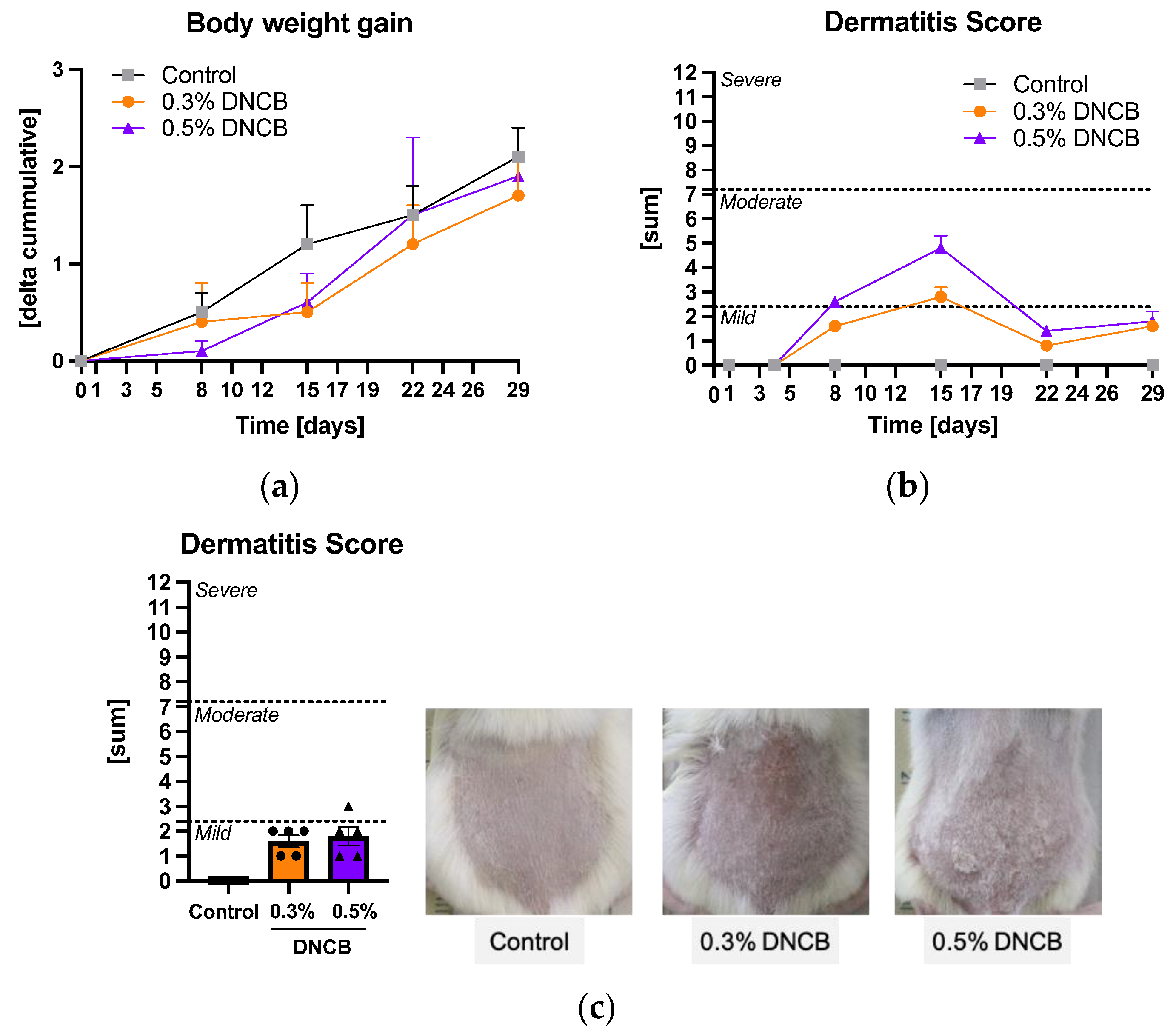
2.1. Effects of DNCB on the Disease Severity
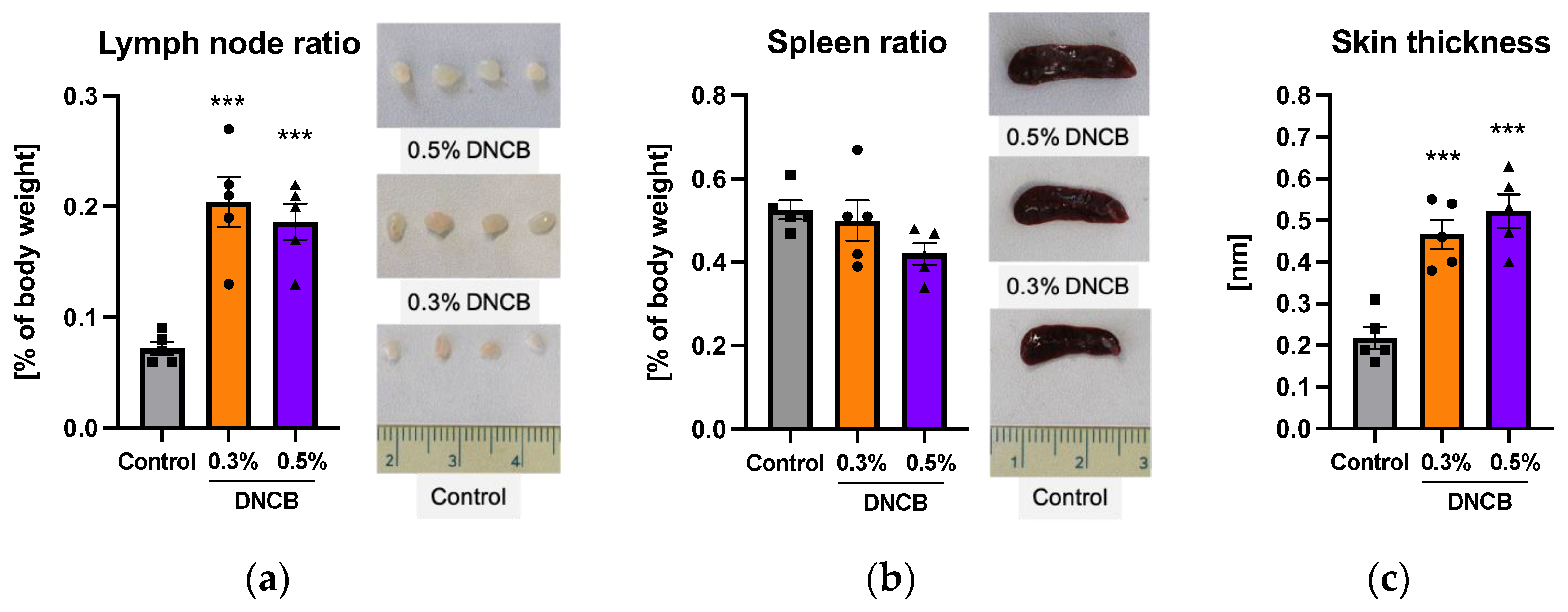
2.2. Effects of DNCB on Secondary Lymphoid Organs
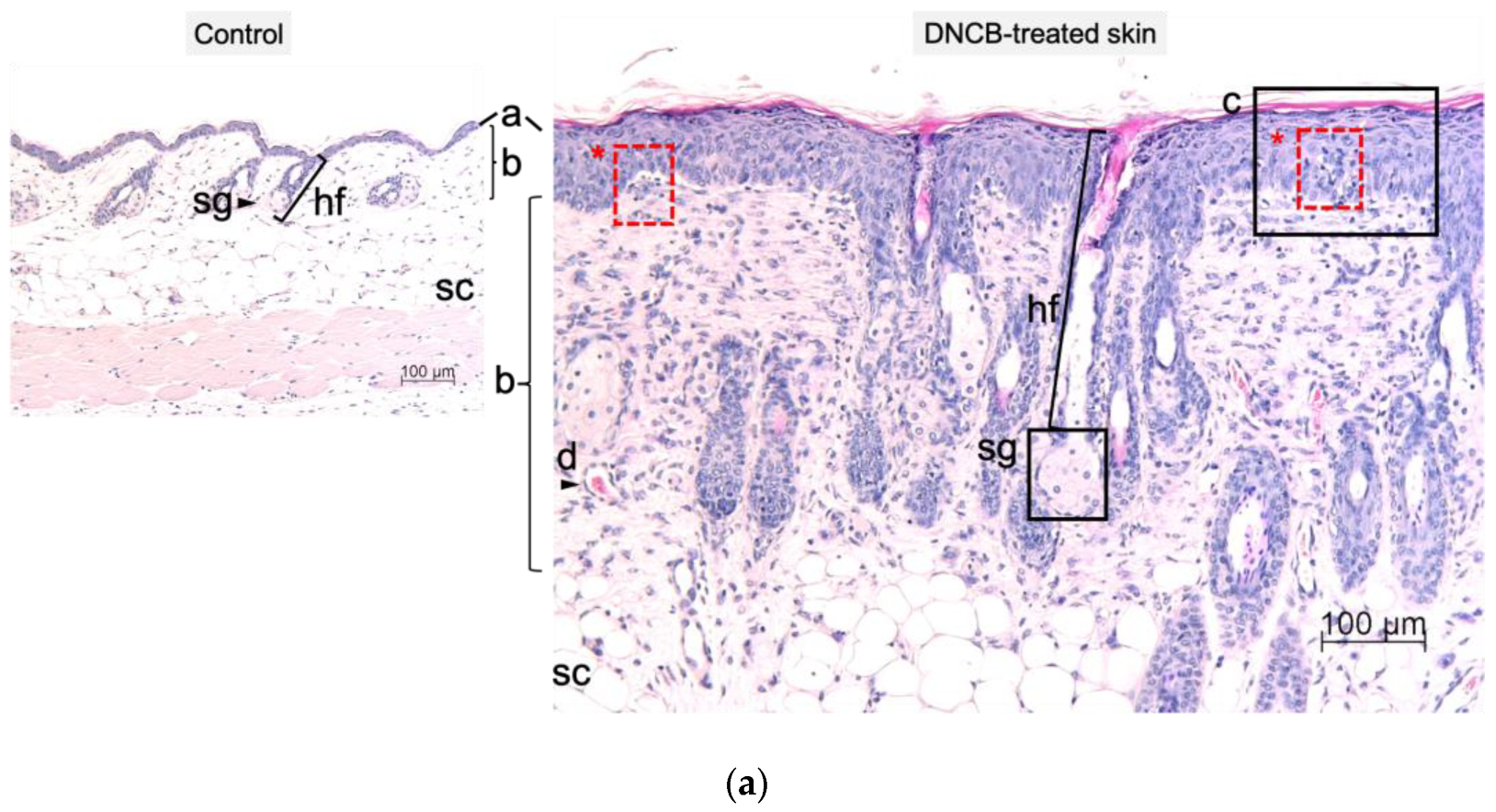
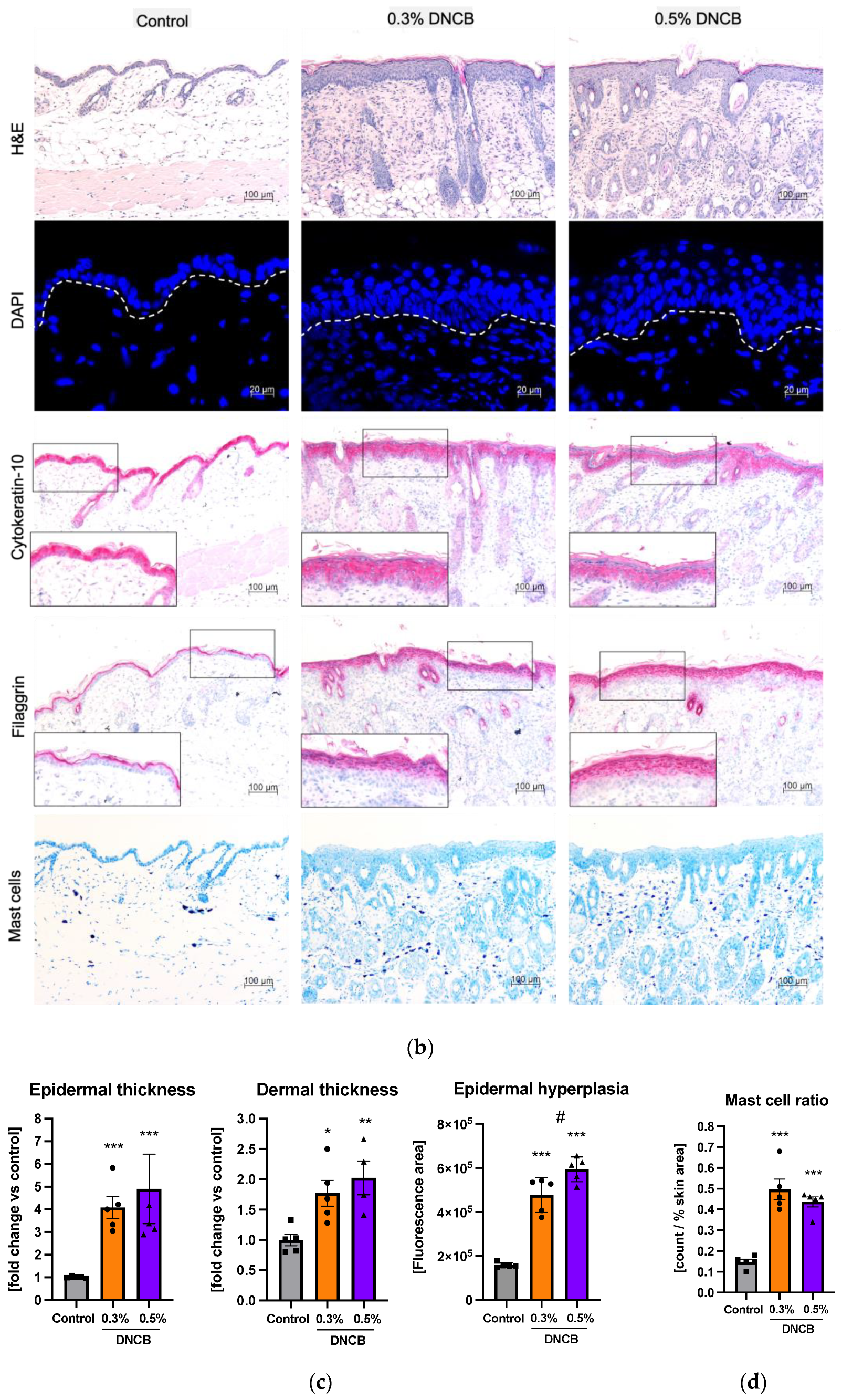
2.3. Histological and Immunohistological Changes of DNCB-Treated Skin
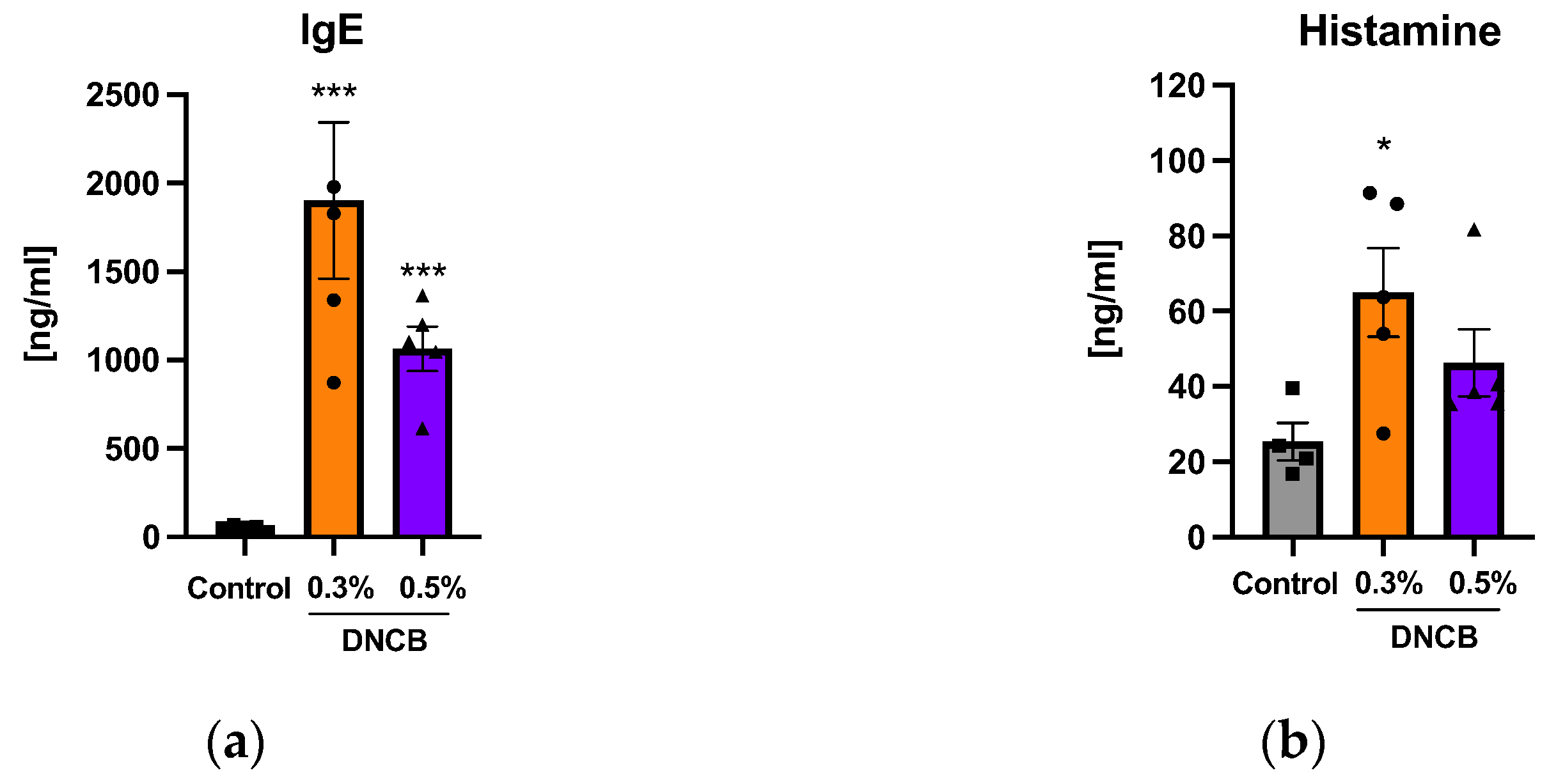
2.4. Effects of DNCB on IgE and Histamine Concentrations
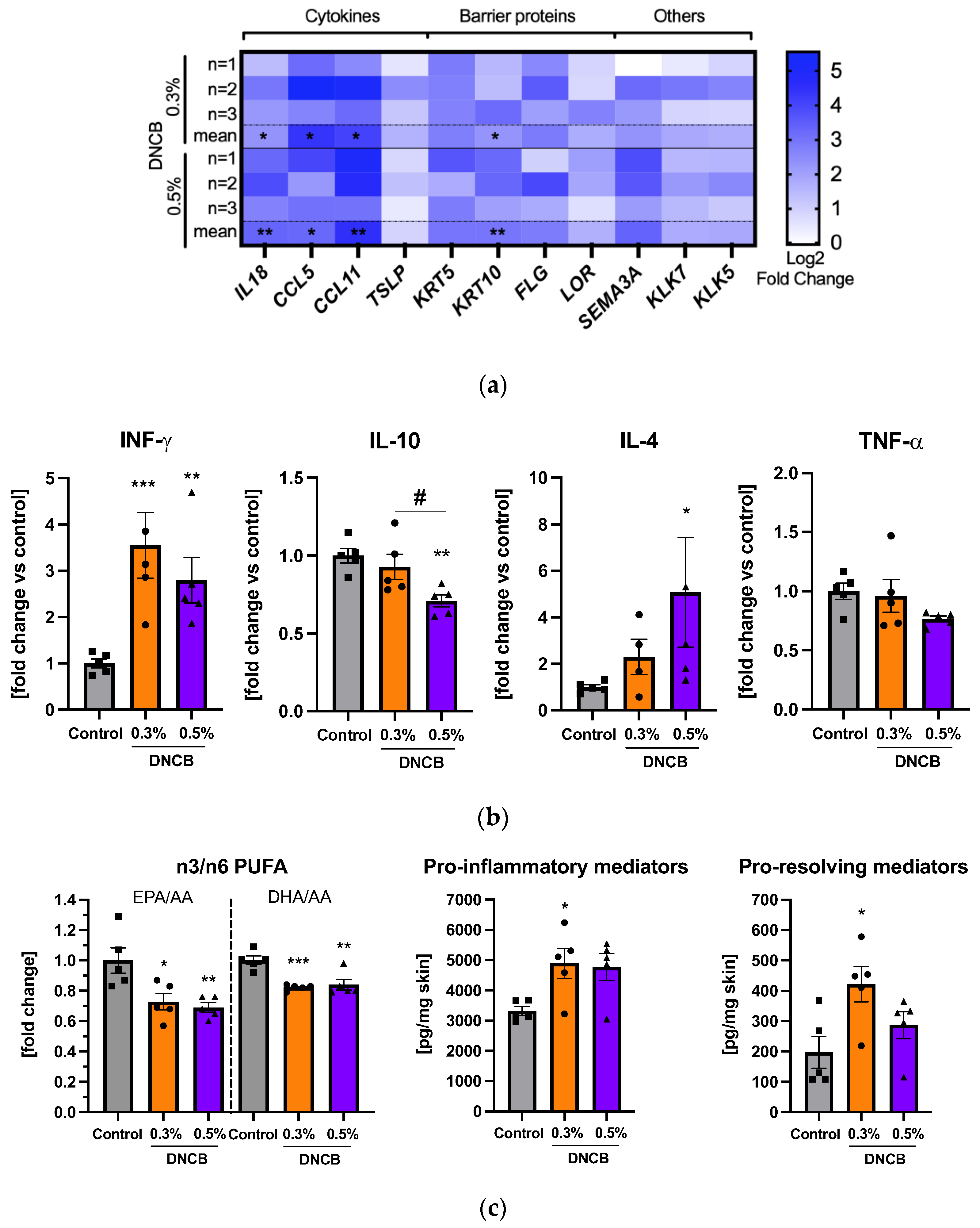
2.5. Effects of DNCB on Gene and Cytokine Expression and Lipid Mediator Formation
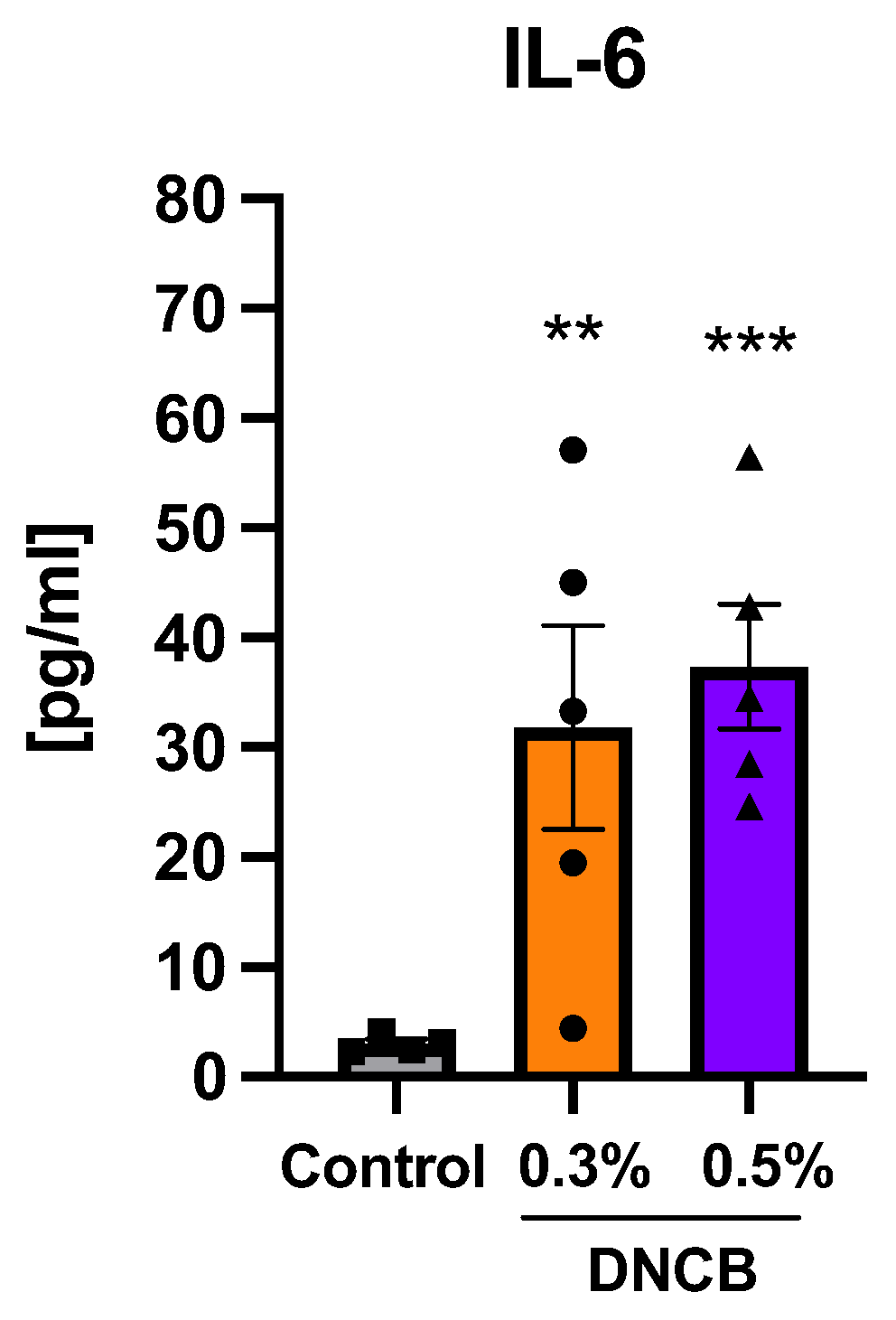
2.6. Effects of DNCB on Cytokine Expression in Splenocytes
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals and Housing Conditions
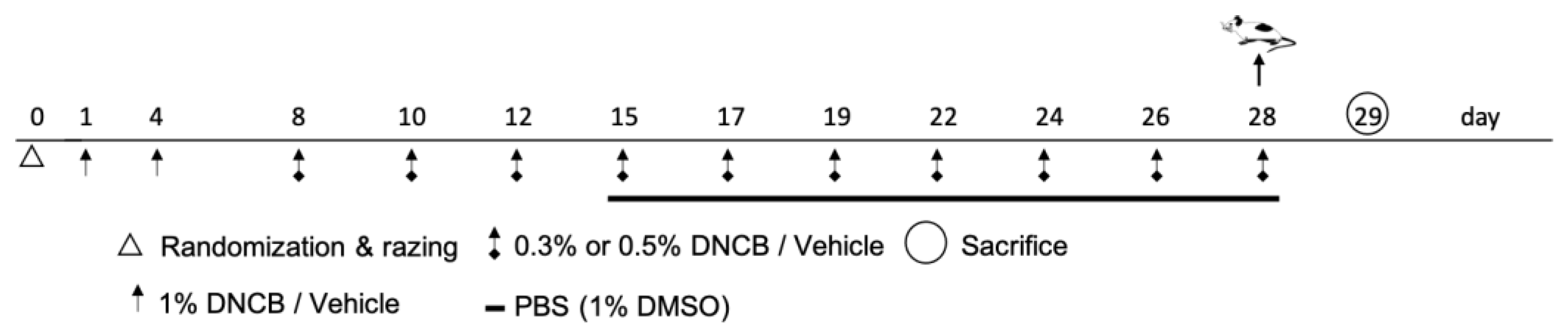
4.3. Induction of Atopic Dermatitis
4.4. Scoring
4.5. Organ Preparation and FACS Measurement
4.6. Measurement of IgE and Histamine Concentration in Plasma
4.7. Cytokine Measurement in the Skin
4.8. Histological and Immunohistological Staining
4.9. RNA Isolation, cDNA Synthesis, and Quantitative Real-Time PCR
4.10. Lipid Mediator Metabololipidomics by UPLC-MS-MS
4.11. Splenocyte Cell Culture
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B
References
- Asher, M.I.; Montefort, S.; Björkstén, B.; Lai, C.K.; Strachan, D.P.; Weiland, S.K.; Williams, H. Worldwide Time Trends in the Prevalence of Symptoms of Asthma, Allergic Rhinoconjunctivitis, and Eczema in Childhood: ISAAC Phases One and Three Repeat Multicountry Cross-Sectional Surveys. Lancet 2006, 368, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Nutten, S. Atopic Dermatitis: Global Epidemiology and Risk Factors. Ann. Nutr. Metab. 2015, 66, 8–16. [Google Scholar] [CrossRef]
- Gittler, J.K.; Shemer, A.; Suárez-Fariñas, M.; Fuentes-Duculan, J.; Gulewicz, K.J.; Wang, C.Q.F.; Mitsui, H.; Cardinale, I.; de Guzman Strong, C.; Krueger, J.G.; et al. Progressive Activation of TH2/TH22 Cytokines and Selective Epidermal Proteins Characterizes Acute and Chronic Atopic Dermatitis. J. Allergy Clin. Immunol. 2012, 130, 1344–1354. [Google Scholar] [CrossRef] [Green Version]
- Guttman-Yassky, E.; Nograles, K.E.; Krueger, J.G. Contrasting Pathogenesis of Atopic Dermatitis and Psoriasis—Part I: Clinical and Pathologic Concepts. J. Allergy Clin. Immunol. 2011, 127, 1110–1118. [Google Scholar] [CrossRef]
- Burgess, J.A.; Dharmage, S.C.; Byrnes, G.B.; Matheson, M.C.; Gurrin, L.C.; Wharton, C.L.; Johns, D.P.; Abramson, M.J.; Hopper, J.L.; Walters, E.H. Childhood Eczema and Asthma Incidence and Persistence: A Cohort Study from Childhood to Middle Age. J. Allergy Clin. Immunol. 2008, 122, 280–285. [Google Scholar] [CrossRef] [Green Version]
- Dharmage, S.C.; Lowe, A.J.; Matheson, M.C.; Burgess, J.A.; Allen, K.J.; Abramson, M.J. Atopic Dermatitis and the Atopic March Revisited. Allergy 2014, 69, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Martel, B.C.; Litman, T.; Hald, A.; Norsgaard, H.; Lovato, P.; Dyring-Andersen, B.; Skov, L.; Thestrup-Pedersen, K.; Skov, S.; Skak, K.; et al. Distinct Molecular Signatures of Mild Extrinsic and Intrinsic Atopic Dermatitis. Exp. Dermatol. 2016, 25, 453–459. [Google Scholar] [CrossRef]
- Tokura, Y. Extrinsic and Intrinsic Types of Atopic Dermatitis. J. Dermatol. Sci. 2010, 58, 1–7. [Google Scholar] [CrossRef]
- Novak, N.; Bieber, T. Allergic and Nonallergic Forms of Atopic Diseases. J. Allergy Clin. Immunol. 2003, 112, 252–262. [Google Scholar] [CrossRef]
- Bawany, F.; Northcott, C.A.; Beck, L.A.; Pigeon, W.R. Sleep Disturbances and Atopic Dermatitis: Relationships, Methods for Assessment, and Therapies. J. Allergy Clin. Immunol. Pract. 2021, 9, 1488–1500. [Google Scholar] [CrossRef] [PubMed]
- Schonmann, Y.; Mansfield, K.E.; Hayes, J.F.; Abuabara, K.; Roberts, A.; Smeeth, L.; Langan, S.M. Atopic Eczema in Adulthood and Risk of Depression and Anxiety: A Population-Based Cohort Study. J. Allergy Clin. Immunol. Pract. 2020, 8, 248–257.e16. [Google Scholar] [CrossRef] [PubMed]
- Thyssen, J.P.; Hamann, C.R.; Linneberg, A.; Dantoft, T.M.; Skov, L.; Gislason, G.H.; Wu, J.J.; Egeberg, A. Atopic Dermatitis Is Associated with Anxiety, Depression, and Suicidal Ideation, but not with Psychiatric Hospitalization or Suicide. Allergy 2018, 73, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; He, R.; Oyoshi, M.; Geha, R.S. Animal Models of Atopic Dermatitis. J. Investig. Dermatol. 2009, 129, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilhar, A.; Reich, K.; Keren, A.; Kabashima, K.; Steinhoff, M.; Paus, R. Mouse Models of Atopic Dermatitis: A Critical Reappraisal. Exp. Dermatol. 2021, 30, 319–336. [Google Scholar] [CrossRef]
- Pickard, C.; Smith, A.M.; Cooper, H.; Strickland, I.; Jackson, J.; Healy, E.; Friedmann, P.S. Investigation of Mechanisms Underlying the T-Cell Response to the Hapten 2,4-Dinitrochlorobenzene. J. Investig. Dermatol. 2007, 127, 630–637. [Google Scholar] [CrossRef] [Green Version]
- Jin, W.; Huang, W.; Chen, L.; Jin, M.; Wang, Q.; Gao, Z.; Jin, Z. Topical Application of JAK1/JAK2 Inhibitor Momelotinib Exhibits Significant Anti-Inflammatory Responses in DNCB-Induced Atopic Dermatitis Model Mice. Int. J. Mol. Sci. 2018, 19, 3973. [Google Scholar] [CrossRef] [Green Version]
- Hou, D.-D.; Di, Z.-H.; Qi, R.-Q.; Wang, H.-X.; Zheng, S.; Hong, Y.-X.; Guo, H.; Chen, H.-D.; Gao, X.-H. Sea Buckthorn (Hippophaë rhamnoides L.) Oil Improves Atopic Dermatitis-Like Skin Lesions via Inhibition of NF-κB and STAT1 Activation. Skin Pharmacol. Physiol. 2017, 30, 268–276. [Google Scholar] [CrossRef]
- Kang, B.-K.; Kim, M.-J.; Kim, K.-B.-W.-R.; Ahn, D.-H. In Vivo and In Vitro Inhibitory Activity of an Ethanolic Extract of Sargassum fulvellum and Its Component Grasshopper Ketone on Atopic Dermatitis. Int. Immunopharmacol. 2016, 40, 176–183. [Google Scholar] [CrossRef]
- Kim, S.R.; Choi, H.-S.; Seo, H.S.; Ku, J.M.; Hong, S.H.; Yoo, H.H.; Shin, Y.C.; Ko, S.-G. Oral Administration of Herbal Mixture Extract Inhibits 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis in BALB/c Mice. Mediat. Inflamm. 2014, 2014, 319438. [Google Scholar] [CrossRef]
- Han, S.-C.; Kang, G.-J.; Ko, Y.-J.; Kang, H.-K.; Moon, S.-W.; Ann, Y.-S.; Yoo, E.-S. Fermented Fish Oil Suppresses T helper 1/2 Cell Response in a Mouse Model of Atopic Dermatitis via Generation of CD4+CD25+Foxp3+ T Cells. BMC Immunol. 2012, 13, 44. [Google Scholar] [CrossRef] [Green Version]
- Heo, J.H.; Heo, Y.; Lee, H.J.; Kim, M.; Shin, H.Y. Topical Anti-Inflammatory and Anti-Oxidative Effects of porcine Placenta Extracts on 2,4-dinitrochlorobenzene-Induced Contact Dermatitis. BMC Complement Altern. Med. 2018, 18, 331. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, P.; Zhang, J.; Hong, T. Inhibitory Effect of Bisdemethoxycurcumin on DNCB-Induced Atopic Dermatitis in Mice. Molecules 2022, 28, 293. [Google Scholar] [CrossRef]
- Jang, S.; Ohn, J.; Kim, J.W.; Kang, S.M.; Jeon, D.; Heo, C.Y.; Lee, Y.-S.; Kwon, O.; Kim, K.H. Caffeoyl–Pro–His Amide Relieve DNCB-Induced Atopic Dermatitis-Like Phenotypes in BALB/c mice. Sci. Rep. 2020, 10, 8417. [Google Scholar] [CrossRef] [PubMed]
- Council of Europe. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes; Council of Europe: Strasbourg, France, 2010. [Google Scholar]
- Pabst, R. Plasticity and Heterogeneity of Lymphoid Organs. Immunol. Lett. 2007, 112, 1–8. [Google Scholar] [CrossRef]
- Lewis, S.M.; Williams, A.; Eisenbarth, S.C. Structure and Function of the Immune System in the Spleen. Sci. Immunol. 2019, 4, eaau6085. [Google Scholar] [CrossRef]
- Carr, M.M.; Botham, P.A.; Gawkrodger, D.J.; Mcvittie, E.; Ross, J.A.; Stewart, I.C.; Hunter, J.A.A. Early Cellular Reactions Induced by Dinitrochlorobenzene in Sensitized Human Skin. Br. J. Dermatol. 1984, 110, 637–641. [Google Scholar] [CrossRef]
- Gawkrodger, D.J.; Haftek, M.; Botham, P.A.; Carr, M.M.; Spencer, M.-J.; Ross, J.A.; Hunter, J.A.A.; Thivolet, J. The Hapten in Contact Hypersensitivity to Dinitrochlorobenzene: Immunoelectron Microscopic and Immunofluorescent Studies. Dermatology 1989, 178, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.A.; Song, H.-Y.; Byun, E.-H.; Ahn, N.-G.; Kim, H.-M.; Nam, Y.R.; Lee, G.H.; Jang, B.-S.; Choi, D.S.; Lee, D.-E.; et al. Gamma-Irradiated Black Ginseng Extract Inhibits Mast Cell Degranulation and Suppresses Atopic Dermatitis-Like Skin Lesions in Mice. Food Chem. Toxicol. 2018, 111, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Ju, D.B.; Kye, Y.-C.; Ju, Y.-J.; Kim, C.G.; Lee, I.K.; Park, S.-M.; Choi, I.S.; Cho, K.K.; Lee, S.H.; et al. Galectin-9 Induced by Dietary Probiotic Mixture Regulates Immune Balance to Reduce Atopic Dermatitis Symptoms in Mice. Front. Immunol. 2020, 10, 3063. [Google Scholar] [CrossRef]
- Kim, Y.-E.; Cho, N.; Cheon, S.; Kim, K.K. Bortezomib, A Proteasome Inhibitor, Alleviates Atopic Dermatitis by Increasing Claudin 1 Protein Expression. Biochem. Biophys. Res. Commun. 2017, 493, 744–750. [Google Scholar] [CrossRef]
- Li, Y.; Chen, L.; Du, Y.; Huang, D.; Han, H.; Dong, Z. Fluoxetine Ameliorates Atopic Dermatitis-Like Skin Lesions in BALB/c Mice through Reducing Psychological Stress and Inflammatory Response. Front. Pharmacol. 2016, 7, 318. [Google Scholar] [CrossRef] [Green Version]
- Purushothaman, B.; Arumugam, P.; Kulsi, G.; Song, J.M. Design, Synthesis, and Biological Evaluation of novel Catecholopyrimidine Based PDE4 Inhibitor for the Treatment of Atopic Dermatitis. Eur. J. Med. Chem. 2018, 145, 673–690. [Google Scholar] [CrossRef]
- Peng, G.; Mu, Z.; Cui, L.; Liu, P.; Wang, Y.; Wu, W.; Han, X. Anti-IL-33 Antibody Has a Therapeutic Effect in an Atopic Dermatitis Murine Model Induced by 2, 4-Dinitrochlorobenzene. Inflammation 2018, 41, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-S.; Lin, S.-C.; Li, S.; Chiang, Y.-C.; Bracci, N.; Lehman, C.W.; Tang, K.-T.; Lin, C.-C. Phloretin Alleviates Dinitrochlorobenzene-Induced Dermatitis in BALB/c Mice. Int. J. Immunopathol. Pharmacol. 2020, 34, 205873842092944. [Google Scholar] [CrossRef]
- Chang, Y.-S.; Tsai, C.-C.; Yang, P.-Y.; Tang, C.-Y.; Chiang, B.-L. Topical Melatonin Exerts Immunomodulatory Effect and Improves Dermatitis Severity in a Mouse Model of Atopic Dermatitis. Int. J. Mol. Sci. 2022, 23, 1373. [Google Scholar] [CrossRef] [PubMed]
- Bak, D.-H.; Lee, E.; Lee, B.C.; Choi, M.J.; Kwon, T.-R.; Hong, J.; Mun, S.; Lee, K.; Kim, S.; Na, J.; et al. Therapeutic Potential of Topically Administered γ-AlOOH on 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis-Like Lesions in Balb/c Mice. Exp. Dermatol. 2019, 28, 169–176. [Google Scholar] [CrossRef]
- Happle, R.; Echternacht, K. Induction of Hair Growth in Alopecia Areata with D.N.C.B. Lancet 1977, 310, 1002–1003. [Google Scholar] [CrossRef]
- He, H.; Del Duca, E.; Diaz, A.; Kim, H.J.; Gay-Mimbrera, J.; Zhang, N.; Wu, J.; Beaziz, J.; Estrada, Y.; Krueger, J.G.; et al. Mild Atopic Dermatitis Lacks Systemic Inflammation and Shows Reduced Nonlesional Skin Abnormalities. J. Allergy Clin. Immunol. 2021, 147, 1369–1380. [Google Scholar] [CrossRef]
- Biedermann, T.; Skabytska, Y.; Kaesler, S.; Volz, T. Regulation of T Cell Immunity in Atopic Dermatitis by Microbes: The Yin and Yang of Cutaneous Inflammation. Front. Immunol. 2015, 6, 353. [Google Scholar] [CrossRef] [PubMed]
- Rafei-Shamsabadi, D.A.; Klose, C.S.N.; Halim, T.Y.F.; Tanriver, Y.; Jakob, T. Context Dependent Role of Type 2 Innate Lymphoid Cells in Allergic Skin Inflammation. Front. Immunol. 2019, 10, 2591. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.-Y.; Liu, P.; Chai, Y.-W.; Wang, Y.; Ren, S.-H.; Li, Y.-Y.; Zhou, H. Artesunate Attenuates 2, 4-Dinitrochlorobenzene-Induced Atopic Dermatitis by Down-Regulating Th17 Cell Responses in BALB/c Mice. Eur. J. Pharmacol. 2020, 874, 173020. [Google Scholar] [CrossRef]
- Yang, H.-L.; Yang, T.-Y.; Gowrisankar, Y.V.; Liao, C.-H.; Liao, J.-W.; Huang, P.-J.; Hseu, Y.-C. Suppression of LPS-Induced Inflammation by Chalcone Flavokawain A through Activation of Nrf2/ARE-Mediated Antioxidant Genes and Inhibition of ROS/NF κ B Signaling Pathways in Primary Splenocytes. Oxidative Med. Cell. Longev. 2020, 2020, 3476212. [Google Scholar] [CrossRef]
- Lin, G.; Gao, S.; Cheng, J.; Li, Y.; Shan, L.; Hu, Z. 1 β-Hydroxyalantolactone, A Sesquiterpene Lactone from Inula japonica, Attenuates Atopic Dermatitis-Like Skin Lesions Induced by 2,4-Dinitrochlorobenzene in the Mouse. Pharm. Biol. 2016, 54, 516–522. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Li, H.; Li, Y.; Li, M.; Chen, G. Effect of Isoliquiritigenin for the Treatment of Atopic Dermatitis-Like Skin Lesions in Mice. Arch. Dermatol. Res. 2017, 309, 805–813. [Google Scholar] [CrossRef]
- Komatsu, N.; Saijoh, K.; Kuk, C.; Liu, A.C.; Khan, S.; Shirasaki, F.; Takehara, K.; Diamandis, E.P. Human tissue Kallikrein Expression in the Stratum Corneum and Serum of Atopic Dermatitis Patients. Exp. Dermatol. 2007, 16, 513–519. [Google Scholar] [CrossRef]
- Guo, C.J.; Mack, M.R.; Oetjen, L.K.; Trier, A.M.; Council, M.L.; Pavel, A.B.; Guttman-Yassky, E.; Kim, B.S.; Liu, Q. Kallikrein 7 Promotes Atopic Dermatitis-Associated Itch Independently of Skin Inflammation. J. Investig. Dermatol. 2020, 140, 1244–1252.e4. [Google Scholar] [CrossRef]
- Zhu, Y.; Underwood, J.; Macmillan, D.; Shariff, L.; O’Shaughnessy, R.; Harper, J.I.; Pickard, C.; Friedmann, P.S.; Healy, E.; Di, W.-L. Persistent Kallikrein 5 Activation Induces Atopic Dermatitis-Like Skin Architecture Independent of PAR2 Activity. J. Allergy Clin. Immunol. 2017, 140, 1310–1322.e5. [Google Scholar] [CrossRef] [Green Version]
- Töröcsik, D.; Weise, C.; Gericke, J.; Szegedi, A.; Lucas, R.; Mihaly, J.; Worm, M.; Rühl, R. Transcriptomic and Lipidomic Profiling of Eicosanoid/Docosanoid Signalling in Affected and Non-Affected Skin of Human Atopic Dermatitis Patients. Exp. Dermatol. 2019, 28, 177–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funk, C.D. Prostaglandins and Leukotrienes: Advances in Eicosanoid Biology. Science 2001, 294, 1871–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fogh, K.; Herlin, T.; Kragballe, K. Eicosanoids in Skin of Patients with Atopic Dermatitis: Prostaglandin E and Leukotriene B are Present in Biologically Active Concentrations. J. Allergy Clin. Immunol. 1989, 83, 450–455. [Google Scholar] [CrossRef]
- Hong, S.-H.; Han, J.E.; Ko, J.-S.; Do, S.H.; Lee, E.H.; Cho, M.-H. Quantitative Determination of 12-Hydroxyeicosatetraenoic Acids by Chiral Liquid Chromatography Tandem Mass Spectrometry in a Murine Atopic Dermatitis Model. J. Vet. Sci. 2015, 16, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bieber, T. Atopic Dermatitis. Ann. Dermatol. 2010, 22, 125. [Google Scholar] [CrossRef] [Green Version]
- Spergel, J.M.; Mizoguchi, E.; Oettgen, H.; Bhan, A.K.; Geha, R.S. Roles of TH1 and TH2 Cytokines in a Murine Model of Allergic Dermatitis. J. Clin. Invest. 1999, 103, 1103–1111. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Martinez, O.; Overbergh, L.; Mathieu, C.; Prabhakar, B.S.; Chan, L.S. Early Up-Regulation of Th2 Cytokines and Late Surge of Th1 Cytokines in an Atopic Dermatitis Model. Clin. Exp. Immunol. 2004, 138, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Kitagaki, H.; Ono, N.; Hayakawa, K.; Kitazawa, T.; Watanabe, K.; Shiohara, T. Repeated Elicitation of Contact Hypersensitivity Induces A Shift in Cutaneous Cytokine Milieu from A T Helper Cell Type 1 to A T Helper Cell Type 2 Profile. J. Immunol. 1997, 159, 2484–2491. [Google Scholar] [CrossRef]
- DaSilva-Arnold, S.C.; Thyagarajan, A.; Seymour, L.J.; Yi, Q.; Bradish, J.R.; Al-Hassani, M.; Zhou, H.; Perdue, N.J.; Nemeth, V.; Krbanjevic, A.; et al. Phenotyping Acute and Chronic Atopic Dermatitis-Like Lesions in Stat6VT Mice Identifies a Role for IL-33 in Disease Pathogenesis. Arch. Dermatol. Res. 2018, 310, 197–207. [Google Scholar] [CrossRef]
- Kim, J.; Lee, J.; Shin, S.; Cho, A.; Heo, Y. Molecular Mechanism of Atopic Dermatitis Induction Following Sensitization and Challenge with 2,4-Dinitrochlorobenzene in Mouse Skin Tissue. Toxicol. Res. 2018, 34, 7–12. [Google Scholar] [CrossRef] [Green Version]
- An, H.-J.; Kim, J.-Y.; Kim, W.-H.; Gwon, M.-G.; Gu, H.M.; Jeon, M.J.; Han, S.-M.; Pak, S.C.; Lee, C.-K.; Park, I.S.; et al. Therapeutic Effects of Bee Venom and Its Major Component, Melittin, on Atopic Dermatitis In Vivo And In Vitro: Effects of Bee Venom and Melittin on Atopic Eczema. Br. J. Pharmacol. 2018, 175, 4310–4324. [Google Scholar] [CrossRef] [Green Version]
- Mlitz, V.; Latreille, J.; Gardinier, S.; Jdid, R.; Drouault, Y.; Hufnagl, P.; Eckhart, L.; Guinot, C.; Tschachler, E. Impact of Filaggrin Mutations on Raman Spectra and Biophysical Properties of the Stratum Corneum in Mild to Moderate Atopic Dermatitis: Raman Profiling of Mild to Moderate Atopic Dermatitis. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 983–990. [Google Scholar] [CrossRef]
- FELASA Working Group on Revision of Guidelines for Health Monitoring of Rodents and Rabbits; Mähler Convenor, M.; Berard, M.; Feinstein, R.; Gallagher, A.; Illgen-Wilcke, B.; Pritchett-Corning, K.; Raspa, M. FELASA Recommendations for the Health Monitoring of Mouse, Rat, Hamster, Guinea Pig and Rabbit Colonies in Breeding and Experimental Units. Lab. Anim. 2014, 48, 178–192. [Google Scholar] [CrossRef]
- European Task Force on Atopic Dermatitis. Severity Scoring of Atopic Dermatitis: The SCORAD Index. Dermatology 1993, 186, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Pace, S.; Jordan, P.M.; Peltner, L.K.; Weber, A.; Fischer, D.; Hofstetter, R.K.; Chen, X.; Werz, O. Beneficial Modulation of Lipid Mediator Biosynthesis in Innate Immune Cells by Antirheumatic Tripterygium wilfordii Glycosides. Biomolecules 2021, 11, 746. [Google Scholar] [CrossRef]
- Werner, M.; Jordan, P.M.; Romp, E.; Czapka, A.; Rao, Z.; Kretzer, C.; Koeberle, A.; Garscha, U.; Pace, S.; Claesson, H.; et al. Targeting Biosynthetic Networks of the Proinflammatory and Proresolving Lipid Metabolome. FASEB J. 2019, 33, 6140–6153. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riedl, R.; Kühn, A.; Rietz, D.; Hebecker, B.; Glowalla, K.-G.; Peltner, L.K.; Jordan, P.M.; Werz, O.; Lorkowski, S.; Wiegand, C.; et al. Establishment and Characterization of Mild Atopic Dermatitis in the DNCB-Induced Mouse Model. Int. J. Mol. Sci. 2023, 24, 12325. https://doi.org/10.3390/ijms241512325
Riedl R, Kühn A, Rietz D, Hebecker B, Glowalla K-G, Peltner LK, Jordan PM, Werz O, Lorkowski S, Wiegand C, et al. Establishment and Characterization of Mild Atopic Dermatitis in the DNCB-Induced Mouse Model. International Journal of Molecular Sciences. 2023; 24(15):12325. https://doi.org/10.3390/ijms241512325
Chicago/Turabian StyleRiedl, Rebecca, Annika Kühn, Denise Rietz, Betty Hebecker, Karl-Gunther Glowalla, Lukas K. Peltner, Paul M. Jordan, Oliver Werz, Stefan Lorkowski, Cornelia Wiegand, and et al. 2023. "Establishment and Characterization of Mild Atopic Dermatitis in the DNCB-Induced Mouse Model" International Journal of Molecular Sciences 24, no. 15: 12325. https://doi.org/10.3390/ijms241512325