
BFNB Enhances Hair Growth in C57BL/6 Mice through the Induction of EGF and FGF7 Factors and the PI3K-AKT-β-Catenin Pathway
 and
and
Abstract
:
1. Introduction
2. Results
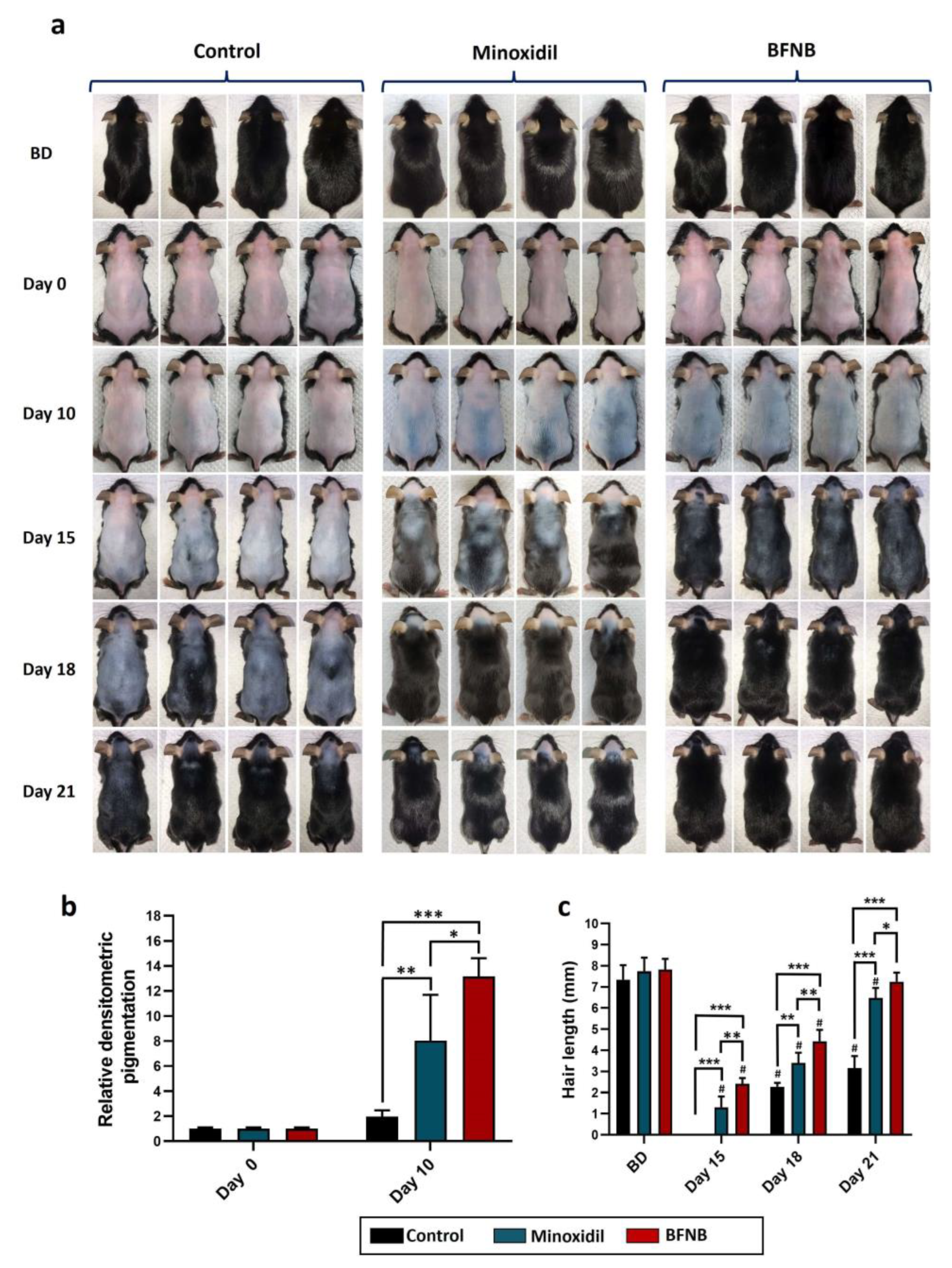
2.1. Effect of a Nanostructured Formulation of B. procumbens on Hair Growth Regulation and Follicular Phase Dynamics in the Dorsa of C57BL/6 Mice
2.2. Effect of BFNB Formulation on the Modulation of Hair Growth and Follicular Phase Dynamics on the Heads of C57BL/6 Mice
2.3. Histological Analysis of the BFNB Effect on the Dorsa of C57BL/6 Mice
2.4. In Silico Prediction of Signaling Pathways Related to the Modulation of Hair Growth by EGF and FGF7
2.5. Immunodetection of Hair-Growth-Inducing Proteins
3. Discussion
4. Materials and Methods
4.1. Lipophilic Serum Containing the Nanostructured BFNB Formulation
4.2. Animals
4.3. Hair Growth in C57BL/6 Mice
4.4. Follicular Analysis
4.5. Histological Study
4.6. Interactomics of EGF and FGF7 Growth Factors
4.7. Western Blotting
4.8. Immunohistochemistry
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Park, A.M.; Khan, S.; Rawnsley, J. Hair Biology: Growth and Pigmentation. Facial Plast. Surg. Clin. N. Am. 2018, 26, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.W.; Kloepper, J.; Langan, E.A.; Kim, Y.; Yeo, J.; Kim, M.J.; Hsi, T.C.; Rose, C.; Yoon, G.S.; Lee, S.J.; et al. A guide to studying human Hair follicle cycling in vivo. J. Investig. Dermatol. 2016, 136, 33–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grymowicz, M.; Rudnicka, E.; Podfigurna, A.; Napierala, P.; Smolarczyk, R.; Smolarczyk, K.; Meczekalski, B. Hormonal effects on hair follicles. Int. J. Mol. Sci. 2020, 21, 5342. [Google Scholar] [CrossRef] [PubMed]
- Piérard-Franchimont, C.; Piérard, G.E. Alterations in hair follicle dynamics in women. BioMed Res. Int. 2013, 2013, 957432. [Google Scholar] [CrossRef] [Green Version]
- Al-Aradi, I.; Al-Ghareeb, M. Review Article: Hair Fall: Common Causes and Treatment Modalities Review Article: Hair Fall: Common Causes and Treatment Modalities. Pan Arab Leag. Dermatol. 2005, 16, 9–15. [Google Scholar]
- American Academy of Dermatology. Hair Loss Resource Center. 2022. Available online: https://www.aad.org/public/diseases/hair-loss (accessed on 24 February 2023).
- Asfour, L.; Cranwell, W.; Sinclair, R. Male Androgenetic Alopecia. In Endotext; MDText.com, Inc.: South Dartmouth, MA, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK278957/?report=reader (accessed on 20 May 2023).
- Oiwoh, S.; Akinboro, A.; Olasode, O.; Onayemi, E. Androgenetic alopecia: Prevalence and clinical characteristics in a South-West Nigerian population. Niger. J. Med. 2021, 30, 507. [Google Scholar] [CrossRef]
- Wang, H.; Pan, L.; Wu, Y. Epidemiological Trends in Alopecia Areata at the Global, Regional, and National Levels. Front. Immunol. 2022, 13, 874677. [Google Scholar] [CrossRef] [PubMed]
- Ateeq Ahmad, S.S. A New Topical Formulation of Minoxidil and Finasteride Improves Hair Growth in Men with Androgenetic Alopecia. J. Clin. Exp. Dermatol. Res. 2015, 6, 253. [Google Scholar] [CrossRef]
- Rathi, V.; Rathi, J.C.; Tamizharasi, S.; Kumar, A. Plants used for hair growth promotion: A review. Rev. Lit. Arts Am. 2008, 2, 185. [Google Scholar]
- Jain, P.K.; Das, D.; Singhai, A.K. Alternative herbal drugs used for treating hair disease. Asian J. Pharm. Clin. Res. 2016, 9, 75–77. [Google Scholar]
- Kim, J.A.; Eun, H.C. Effect of several growth factors on hair follicle growth in hair follicle organ culture. Ann. Dermatol. 1994, 6, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Peus, D.; Pittelkow, M.R. Growth factors in hair organ development and the hair growth cycle. Dermatol. Clin. 1996, 14, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Hamada, K.; Ozawa, K.; Itami, S.; Yoshikawa, K. 196 Fibroblast growth factor 10 expression in human hair follicle. J. Dermatol. Sci. 1997, 15, 135. [Google Scholar] [CrossRef]
- Hwang, K.A.; Hwang, Y.L.; Lee, M.H.; Kim, N.R.; Roh, S.S.; Lee, Y.; Kim, C.D.; Lee, J.H.; Choi, K.C. Adenosine stimulates growth of dermal papilla and lengthens the anagen phase by increasing the cysteine level via fibroblast growth factors 2 and 7 in an organ culture of mouse vibrissae hair follicles. Int. J. Mol. Med. 2012, 29, 195–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trüeb, R.M. Further clinical evidence for the effect of IGF-1 on hair growth and alopecia. Ski. Appendage Disord. 2018, 4, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Du Cros, D.L. Fibroblast Growth Factor and Epidermal Growth Factor in Hair Development. J. Investig. Dermatol. 1993, 101, 106S–113S. [Google Scholar] [CrossRef]
- Jindo, T.; Tsuboi, R.; Imai, R.; Takamori, K.; Rubin, J.S.; Ogawa, H. Hepatocyte growth factor stimulates hair growth of mouse vibrissae in organ culture. J. Dermatol. Sci. 1993, 6, 104. [Google Scholar] [CrossRef]
- Lachgar, S.; Moukadiri, H.; Jonca, F.; Charveron, M.; Bouhaddioui, N.; Gall, Y.; Bonafe, J.L.; Plouët, J. Vascular endothelial growth factor is an autocrine growth factor for hair dermal papilla cells. J. Investig. Dermatol. 1996, 106, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Danilenko, D.M.; Ring, B.D.; Yanagihara, D.; Benson, W.; Wiemann, B.; Starnes, C.O.; Pierce, G.F. Keratinocyte growth factor is an important endogenous mediator of hair follicle growth, development, and differentiation: Normalization of the nu/nu follicular differentiation defect amelioration of chemotherapy-induced alopecia. Am. J. Pathol. 1995, 147, 145. [Google Scholar]
- Du Cros, D.L. Fibroblast growth factor and the hair cycle of the hairless mouse. J. Investig. Dermatol. 1995, 104, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.Y.; Su, C.H.; Chiang, C.Y.; Wu, C.N.; Kuan, Y.H. Observation of the Expression of Vascular Endothelial Growth Factor and the Potential Effect of Promoting Hair Growth Treated with Chinese Herbal BeauTop. Evid. Based Complement. Altern. Med. 2021, 2021, 6667011. [Google Scholar] [CrossRef] [PubMed]
- Mak KK, L.; Chan, S.Y. Epidermal growth factor as a biologic switch in hair growth cycle. J. Biol. Chem. 2003, 278, 26120–26126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenquist, T.A.; Martin, G.R. Fibroblast growth factor signalling in the hair growth cycle: Expression of the fibroblast growth factor receptor and ligand genes in the murine hair follicle. Dev. Dyn. 1996, 205, 379–386. [Google Scholar] [CrossRef]
- Martínez-Cuazitl, A.; Gómez-García, M.d.C.; Hidalgo-Alegria, O.; Flores, O.M.; Núñez-Gastélum, J.A.; Martínez ES, M.; Ríos-Cortés, A.M.; Garcia-Solis, M.; Pérez-Ishiwara, D.G. Characterization of Polyphenolic Compounds from Bacopa procumbens and Their Effects on Wound-Healing Process. Molecules 2022, 27, 6521. [Google Scholar] [CrossRef] [PubMed]
- Müller-Röver, S.; Handjiski, B.; Van Der Veen, C.; Eichmüller, S.; Foitzik, K.; McKay, I.A.; Stenn, K.S.; Paus, R. A comprehensive guide for the accurate classification of murine hair follicles in distinct hair cycle stages. J. Investig. Dermatol. 2001, 117, 3–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenn, K.S.; Paus, R. Controls of hair follicle cycling. Physiol. Rev. 2001, 81, 449–494. [Google Scholar] [CrossRef] [Green Version]
- Park, H.J.; Zhang, N.; Park, D.K. Topical application of Polygonum multiflorum extract induces hair growth of resting hair follicles through upregulating Shh and β-catenin expression in C57BL/6 mice. J. Ethnopharmacol. 2011, 135, 369–375. [Google Scholar] [CrossRef]
- Park, H.J.; Jin, G.R.; Jung, J.H.; Hwang, S.B.; Lee, S.H.; Lee, B.H. Hair Growth Promotion Effect of Nelumbinis Semen Extract with High Antioxidant Activity. Evid. Based Complement. Altern. Med. 2021, 2021, 6631042. [Google Scholar] [CrossRef]
- Park, S.; Shin, W.S.; Ho, J. Fructus panax ginseng extract promotes hair regeneration in C57BL/6 mice. J. Ethnopharmacol. 2011, 138, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Begum, A.; Sandhya, S.; Kumar, A.N.; Ali, S.S. Evaluation of Herbal Hair Lotion loaded with Rosemary for Possible Hair Growth in C57BL/6 Mice. Adv. Biomed. Res. 2023, 12, 60. [Google Scholar] [PubMed]
- Park, S. Hair Follicle Morphogenesis During Embryogenesis, Neogenesis, and Organogenesis. Front. Cell Dev. Biol. 2022, 10, 933370. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Zhu, L.; He, J. Morphogenesis, Growth Cycle and Molecular Regulation of Hair Follicles. Front. Cell Dev. Biol. 2022, 10, 899095. [Google Scholar] [CrossRef]
- Hori, H.; Moretti, G.; Rebora, A.; Crovato, F. The thickness of human scalp: Normal and bald. J. Investig. Dermatol. 1972, 58, 396–399. [Google Scholar] [CrossRef] [Green Version]
- Lintzeri, D.A.; Karimian, N.; Blume-Peytavi, U.; Kottner, J. Epidermal thickness in healthy humans: A systematic review and meta-analysis. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 1191–1200. [Google Scholar] [CrossRef] [PubMed]
- Nepal, S.; Venkataram, A.; Mysore, V. The role of adipose tissue in hair regeneration: A potential tool for management? J. Cutan. Aesthetic Surg. 2021, 14, 295. [Google Scholar]
- Walocko, F.M.; Khouri, R.K.; Urbanchek, M.G.; Levi, B.; Cederna, P.S. The potential roles for adipose tissue in peripheral nerve regeneration. Microsurgery 2016, 36, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouhassira, E.E. Adipose: Development and Regeneration Potential. In The SAGE Encyclopedia of Stem Cell Research, 2nd ed.; SAGE Publications, Inc.: Thousand Oaks, CA, USA, 2015. [Google Scholar] [CrossRef]
- Gimble, J.; Rad, M.R.; Yao, S. Adipose Tissue-Derived Stem Cells and Their Regeneration Potential. Stem Cells Craniofacial Dev. Regen. 2013, 46, 241–258. [Google Scholar] [CrossRef]
- Mohseni, M.; Dang, H.P.; Tran, P.A.; Castro, N.J.; Nguyen, T.D.; Ho, H.M.; Tran, M.P.N.; Nguyen, T.H. Adipose tissue regeneration. In Biomaterials in Translational Medicine; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, C.; Zhan, X.; Wang, B.; Li, K.; Li, J. Jagged1 and epidermal growth factor promoted androgen-suppressed mouse hair growth in vitro and in vivo. Front. Pharmacol. 2020, 10, 1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.H.; Xiang, L.J.; Shi, H.X.; Zhang, J.; Jiang, L.P.; Cai, P.T.; Lin, Z.L.; Lin, B.B.; Huang, Y.; Zhang, H.L.; et al. Fibroblast growth factors stimulate hair growth through β -Catenin and shh expression in C57BL/6 mice. BioMed Res. Int. 2015, 2015, 730139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rishikaysh, P.; Dev, K.; Diaz, D.; Shaikh Qureshi, W.M.; Filip, S.; Mokry, J. Signaling involved in hair follicle morphogenesis and development. Int. J. Mol. Sci. 2014, 15, 1647–1670. [Google Scholar] [CrossRef] [Green Version]
- Wall, D.; Meah, N.; Fagan, N.; York, K.; Sinclair, R. Advances in hair growth. Fac. Rev. 2022, 11, 899095. [Google Scholar] [CrossRef] [PubMed]
- Saxena, N.; Mok, K.W.; Rendl, M. An updated classification of hair follicle morphogenesis. Exp. Dermatol. 2019, 28, 332–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Degenstein, L.; Fuchs, E. Keratinocyte growth factor is required for hair development but not for wound healing. Genes Dev. 1996, 10, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita-Ise, M.; Tsukashima, A.; Kinoshita, T.; Yamazaki, Y.; Ohyama, M. Altered FGF expression profile in human scalp-derived fibroblasts upon WNT activation: Implication of their role to provide folliculogenetic microenvironment. Inflamm. Regen. 2020, 40, 35. [Google Scholar] [CrossRef]
- Alam, M.; Bertolini, M.; Gherardini, J.; Keren, A.; Ponce, L.; Chéret, J.; Alenfall, J.; Dunér, P.; Nilsson, A.H.; Gilhar, A.; et al. An osteopontin-derived peptide inhibits human hair growth at least in part by decreasing fibroblast growth factor-7 production in outer root sheath keratinocytes. Br. J. Dermatol. 2020, 182, 1404–1414. [Google Scholar] [CrossRef]
- Kim, J.E.; Lee, Y.J.; Park, H.R.; Lee, D.G.; Jeong, K.H.; Kang, H. The effect of jak inhibitor on the survival, anagen re-entry, and hair follicle immune privilege restoration in human dermal papilla cells. Int. J. Mol. Sci. 2020, 21, 5137. [Google Scholar] [CrossRef]
- Zhang, H.; Nan, W.; Wang, S.; Zhang, T.; Si, H.; Yang, F.; Li, G. Epidermal Growth Factor Promotes Proliferation and Migration of Follicular Outer Root Sheath Cells via Wnt/β-Catenin Signaling. Cell. Physiol. Biochem. 2016, 39, 360–370. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Nan, W.; Wang, S.; Zhang, T.; Si, H.; Wang, D.; Yang, F.; Li, G. Epidermal growth factor promotes proliferation of dermal papilla cells via Notch signaling pathway. Biochimie 2016, 127, 10–18. [Google Scholar] [CrossRef]
- Choi, N.; Kim, W.S.; Oh, S.H.; Sung, J.H. HB-EGF improves the hair regenerative potential of adipose-derived stem cells via ROS generation and HCK phosphorylation. Int. J. Mol. Sci. 2020, 21, 122. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Fu, X.; Ao, Y.; Nan, M.; Qiu, Z.; Jia, X.; Xiao, Y.; Liu, D.; Guo, X. ANXA1 affects murine hair follicle growth through EGF signaling pathway. Gene 2021, 771, 145343. [Google Scholar] [CrossRef]
- Lee, C.Y.; Yang, C.Y.; Lin, C.C.; Yu, M.C.; Sheu, S.J.; Kuan, Y.H. Hair growth is promoted by BeauTop via expression of EGF and FGF-7. Mol. Med. Rep. 2018, 17, 8047–8052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grothe, T.; Wandrey, F.; Schuerch, C. Short communication: Clinical evaluation of pea sprout extract in the treatment of hair loss. Phytother. Res. 2020, 34, 428–431. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, Q.; Bai, Y.; Yang, K.; Ye, Y.; Wu, K.; Huang, J.; Zhang, Y.; Zhang, X.; Thianthanyakij, T.; et al. Autologous activated platelet-rich plasma in hair growth: A pilot study in male androgenetic alopecia with in vitro bioactivity investigation. J. Cosmet. Dermatol. 2021, 20, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- Richardson, G.D.; Bazzi, H.; Fantauzzo, K.A.; Waters, J.M.; Crawford, H.; Hynd, P.; Christiano, A.M.; Jahoda, C.A.B. KGF and EGF signalling block hair follicle induction and promote interfollicular epidermal fate in developing mouse skin. Development 2009, 136, 2153–2164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, B.; Li, J.; Chen, Q.; Yang, N.; Bao, Z.; Hu, S.; Chen, Y.; Wu, X. A treatment combination of IGF and EGF promotes hair growth in the angora rabbit. Genes 2021, 12, 24. [Google Scholar] [CrossRef]
- Chen, D.; Jarrell, A.; Guo, C.; Lang, R.; Atit, R. Dermal β-catenin activity in response to epidermal Wnt ligands is required for fibroblast proliferation and hair follicle initiation. Development 2012, 139, 1522–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Q.; Yu, W.; Fang, Y.; Yao, M.; Yang, P. Beta-catenin can induce hair follicle stem cell differentiation into transit-amplifying cells through c-myc activation. Tissue Cell 2017, 49, 28–34. [Google Scholar] [CrossRef]
- Ryu, Y.C.; Lee, D.H.; Shim, J.; Park, J.; Kim, Y.R.; Choi, S.; Bak, S.S.; Sung, Y.K.; Lee, S.H.; Choi, K.Y. KY19382, a novel activator of Wnt/β-catenin signalling, promotes hair regrowth and hair follicle neogenesis. Br. J. Pharmacol. 2021, 178, 2533–2546. [Google Scholar] [CrossRef]
- Shin, D.W. The Molecular Mechanism of Natural Products Activating Wnt/β-Catenin Signaling Pathway for Improving Hair Loss. Life 2022, 12, 1856. [Google Scholar] [CrossRef]
- Choi, B.Y. Targeting wnt/β-catenin pathway for developing therapies for hair loss. Int. J. Mol. Sci. 2020, 21, 4915. [Google Scholar] [CrossRef]
- Bejaoui, M.; Villareal, M.O.; Isoda, H. β-catenin-mediated hair growth induction effect of 3,4,5-tri-O-caffeoylquinic acid. Aging 2019, 11, 4216–4237. [Google Scholar] [CrossRef] [PubMed]
- Valenta, T.; Hausmann, G.; Basler, K. Focus Review: The many faces and functions of β-catenin. EMBO J. 2012, 31, 2714–2736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Xiao, Q.; Xiao, J.; Niu, C.; Li, Y.; Zhang, X.; Zhou, Z.; Shu, G.; Yin, G. Wnt/β-catenin signalling: Function, biological mechanisms, and therapeutic opportunities. Signal Transduct. Target. Ther. 2022, 7, 3. [Google Scholar] [CrossRef]
- Yano, K.; Brown, L.F.; Detmar, M. Control of hair growth and follicle size by VEGF-mediated angiogenesis. J. Clin. Investig. 2001, 107, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Bai, T.; Liu, F.; Zou, F.; Zhao, G.; Jiang, Y.; Liu, L.; Shi, J.; Hao, D.; Zhang, Q.; Zheng, T.; et al. Epidermal growth factor induces proliferation of hair follicle-derived mesenchymal stem cells through epidermal growth factor receptor-mediated activation of ERK and AKT signaling pathways associated with upregulation of cyclin d1 and downregulation of p16. Stem Cells Dev. 2017, 26, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fan, Z.; Wang, X.; Mo, M.; Zeng, S.B.; Xu, R.H.; Wang, X.; Wu, Y. PI3K/Akt signaling pathway is essential for de novo hair follicle regeneration. Stem Cell Res. Ther. 2020, 11, 144. [Google Scholar] [CrossRef] [Green Version]
- Russo, B.; Brembilla, N.C.; Chizzolini, C. Interplay Between Keratinocytes and Fibroblasts: A Systematic Review Providing a New Angle for Understanding Skin Fibrotic Disorders. Front. Immunol. 2020, 11, 648. [Google Scholar] [CrossRef]
- Lian, A.A.; Yamaji, Y.; Kajiwara, K.; Takaki, K.; Mori, H.; Liew MW, O.; Kotani, E.; Maruta, R. A Bioengineering Approach for the Development of Fibroblast Growth Factor-7-Functionalized Sericin Biomaterial Applicable for the Cultivation of Keratinocytes. Int. J. Mol. Sci. 2022, 23, 9953. [Google Scholar] [CrossRef]
- Verma, R.; Madaan, A.; Joshi, V.; Kishore, A.; Singh, A.T.; Jaggi, M.; Kwan Sung, Y. Dermal Papilla Cells, Keratinocytes, Proliferation, Oxidative stress, VEGF.1 Dermal Papilla Cells, Keratinocytes, Proliferation, Oxidative stress, VEGF. Am. J. Dermatol. Venereol. 2017, 6, 51–57. [Google Scholar]
- Badri, T.; Nessel, T.A.; Kumar, D.D. Minoxidil. StatPearls. National Library of Medicine National Center for Biotechnology Information. 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK482378/#:~:text=Mechanism%20of%20Action2023,-Topical%20minoxidil%20(empirical&text=Inhibition%20of%20androgen%20effects%20on,of%20the%20beta%2Dcatenin%20pathway (accessed on 20 July 2023).
- Madaan, A.; Joshi, V.; Kishore, A.; Verma, R.; Singh, A.T.; Jaggi, M.; Sung, Y.K. In vitro Hair Growth Promoting Effects of Naringenin and Hesperetin on Human Dermal Papilla Cells and Keratinocytes. Am. J. Dermatol. Venereol. 2017, 6, 51–57. [Google Scholar]
- Brotzu, G.; Fadda, A.M.; Manca, M.L.; Manca, T.; Marongiu, F.; Campisi, M.; Consolaro, F. A liposome-based formulation containing equol, dihomo-γ-linolenic acid and propionyl-l-carnitine to prevent and treat hair loss: A prospective investigation. Dermatol. Ther. 2019, 32, e12778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huh, S.; Lee, J.; Jung, E.; Kim, S.C.; Kang, J.I.; Lee, J.; Kim, Y.W.; Sung, Y.K.; Kang, H.K.; Park, D. A cell-based system for screening hair growth-promoting agents. Arch. Dermatol. Res. 2009, 301, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; He, Z.; Xiang, G.; Cai, L.; Xu, Z.; Mao, C.; Feng, Y. Paeoniflorin promotes angiogenesis and tissue regeneration in a full-thickness cutaneous wound model through the PI3K/AKT pathway. J. Cell. Physiol. 2020, 235, 9933–9945. [Google Scholar] [CrossRef] [PubMed]
- Hua, F.Z.; Ying, J.; Zhang, J.; Wang, X.F.; Hu, Y.H.; Liang, Y.P.; Liu, Q.; Xu, G.H. Naringenin pre-treatment inhibits neuroapoptosis and ameliorates cognitive impairment in rats exposed to isoflurane anesthesia by regulating the PI3/Akt/PTEN signalling pathway and suppressing NF-B-mediated inflammation. Int. J. Mol. Med. 2016, 38, 1271–1280. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.C.; Yang, C.H.; Chiou, Y.L. Citrus flavanone naringenin enhances melanogenesis through the activation of Wnt/β-catenin signalling in mouse melanoma cells. Phytomedicine 2011, 18, 1244–1249. [Google Scholar] [CrossRef]
- Xu, Z.; Xu, J.; Li, S.; Cui, H.; Zhang, G.; Ni, X.; Wang, J. S-Equol enhances osteoblastic bone formation and prevents bone loss through OPG/RANKL via the PI3K/Akt pathway in streptozotocin-induced diabetic rats. Front. Nutr. 2022, 9, 986192. [Google Scholar] [CrossRef]
- Zughaibi, T.A.; Suhail, M.; Tarique, M.; Tabrez, S. Targeting pi3k/akt/mtor pathway by different flavonoids: A cancer chemopreventive approach. Int. J. Mol. Sci. 2021, 22, 12455. [Google Scholar] [CrossRef]
- NOM-062-ZOO-1999. Especificaciones Técnicas para la Producción, Cuidado y Uso de los Animales de Laboratorio. 1999. Available online: https://www.gob.mx/cms/uploads/attachment/file/203498/NOM-062-ZOO-1999_220801.pdf (accessed on 25 July 2023).
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dorsum | Head | |||
|---|---|---|---|---|
| Control | Minoxidil | Control | Minoxidil | |
| EGF | 8.1 | 6 | 3.3 | 3.2 |
| FGF7 | 37.4 | 31.9 | 18.5 | 18.9 |
| p-PI3K | 19.4 | 11.1 | 5.5 | 3.4 |
| AKT | −0.7 | −0.3 | −0.2 | 0.3 |
| p-AKT | 26.1 | 24.9 | 8.2 | 4.9 |
| β-catenin | 4.1 | 2.9 | 0.9 | 1 |
| PCNA | 13.7 | 10.3 | 4.7 | 4.6 |
| KI-67 | 32.6 | 26.8 | 13.9 | 13.3 |
| Cyclin D1 | 7.7 | 4.9 | 3.5 | 1.9 |
| Cyclin E | 16.5 | 13.2 | 8.2 | 8.1 |
| Dorsum | Head | |||
|---|---|---|---|---|
| Control | Minoxidil | Control | Minoxidil | |
| EGF | 1.9 | 1.0 | 5.7 | 4.9 |
| FGF7 | 4.3 | 2.2 | 10.5 | 8.7 |
| PCNA | 2.6 | 1.1 | 4.9 | 4.4 |
| β-catenin | 2.1 | 1.2 | 3.3 | 2.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Mora, S.; Ocampo-López, J.; Gómez-García, M.d.C.; Pérez-Ishiwara, D.G. BFNB Enhances Hair Growth in C57BL/6 Mice through the Induction of EGF and FGF7 Factors and the PI3K-AKT-β-Catenin Pathway. Int. J. Mol. Sci. 2023, 24, 12110. https://doi.org/10.3390/ijms241512110
Pérez-Mora S, Ocampo-López J, Gómez-García MdC, Pérez-Ishiwara DG. BFNB Enhances Hair Growth in C57BL/6 Mice through the Induction of EGF and FGF7 Factors and the PI3K-AKT-β-Catenin Pathway. International Journal of Molecular Sciences. 2023; 24(15):12110. https://doi.org/10.3390/ijms241512110
Chicago/Turabian StylePérez-Mora, Salvador, Juan Ocampo-López, María del Consuelo Gómez-García, and David Guillermo Pérez-Ishiwara. 2023. "BFNB Enhances Hair Growth in C57BL/6 Mice through the Induction of EGF and FGF7 Factors and the PI3K-AKT-β-Catenin Pathway" International Journal of Molecular Sciences 24, no. 15: 12110. https://doi.org/10.3390/ijms241512110