

Gating of β-Barrel Protein Pores, Porins, and Channels: An Old Problem with New Facets


Abstract
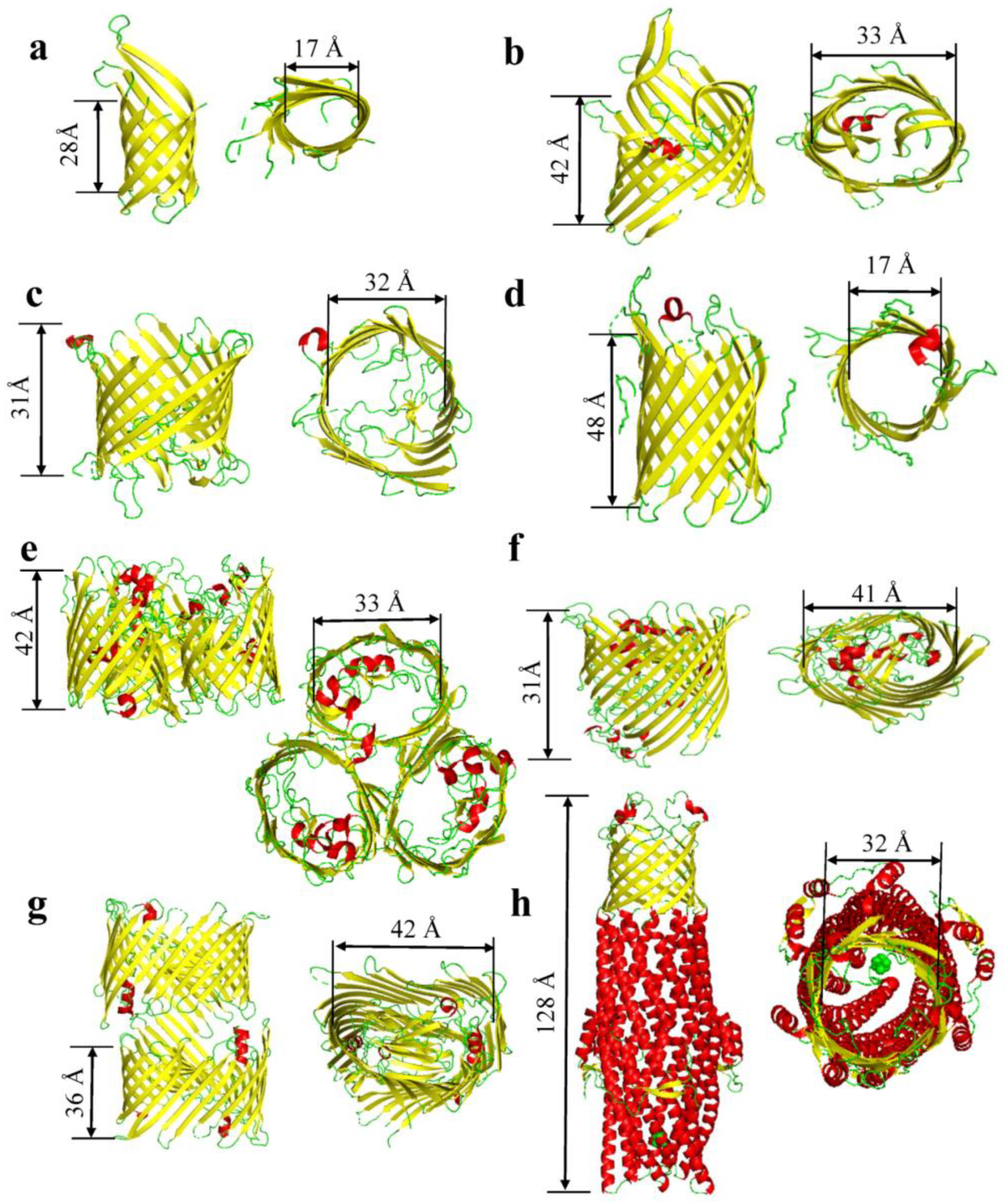
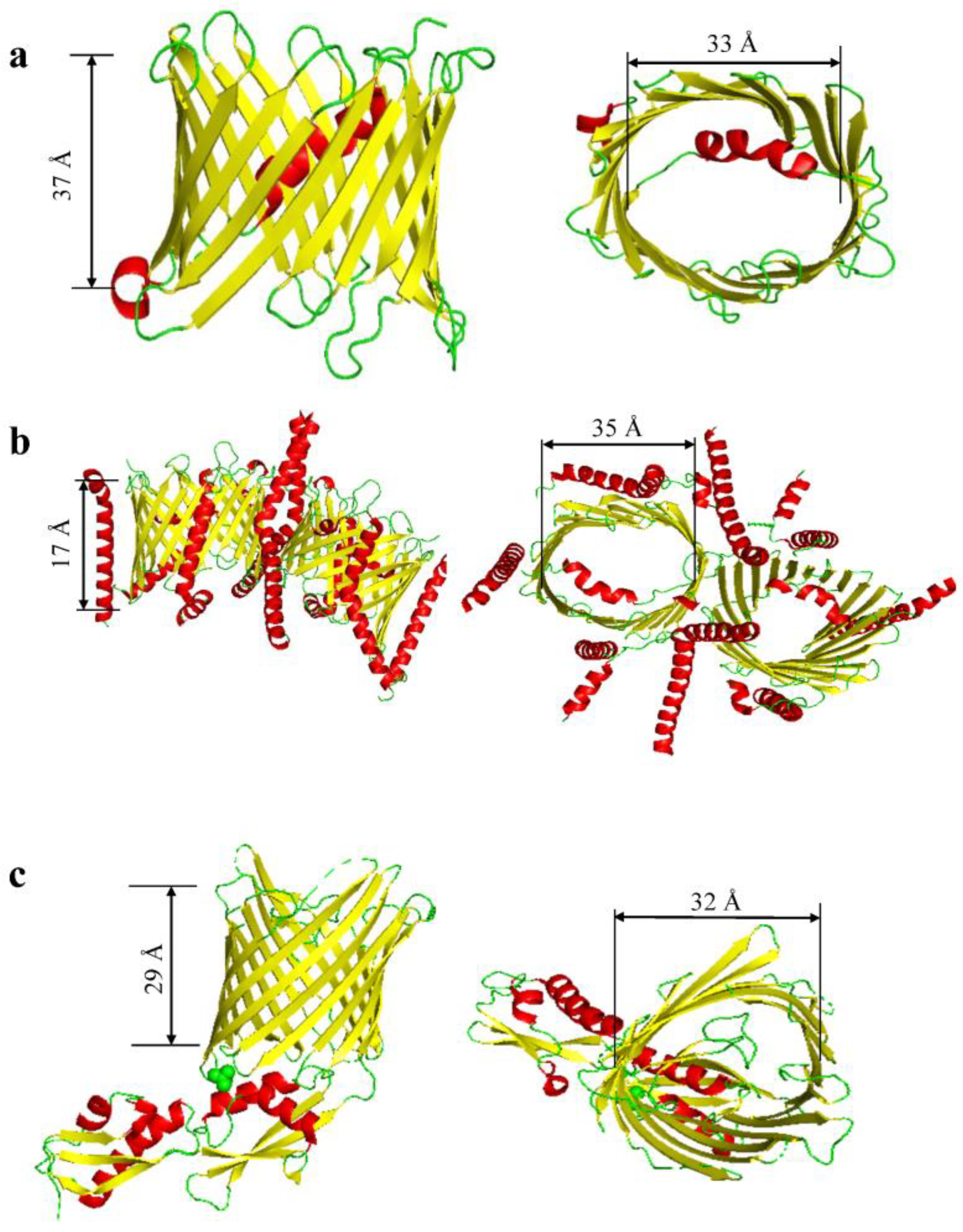
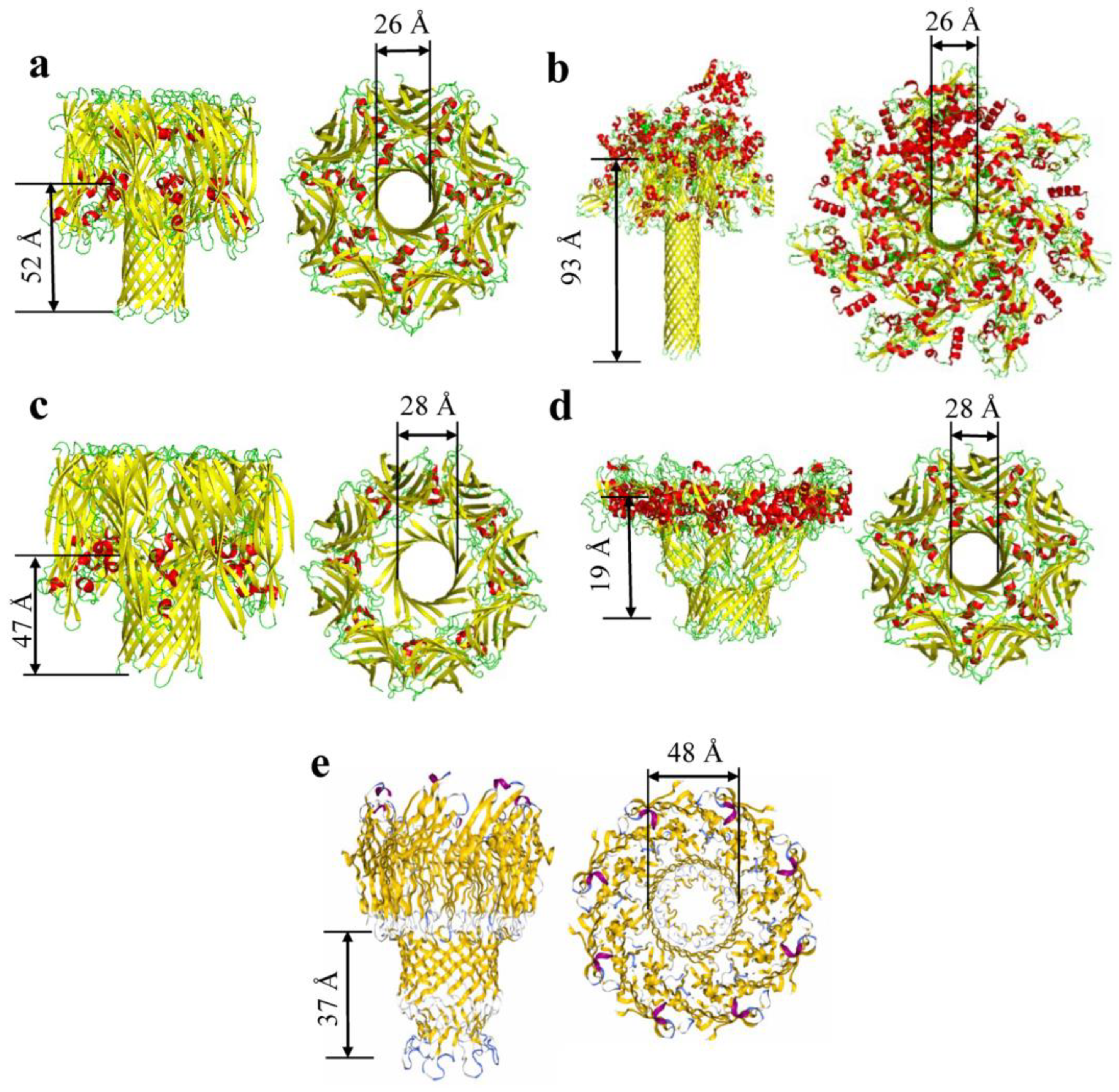
:1. The Structure and Composition of β Barrels




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteins | PDB Code | Average Molecular Weight(kDa) | Oligomeric State | Number of β-Strands per Monomer | Average Internal Diameter (Å) | Length of Barrel (Å) | Corks and Loops | Citation |
|---|---|---|---|---|---|---|---|---|
| OmpA | 1QJP | 37 | Mono/Dimer | 8 | 17 | 28 | 4 Loops | [15] |
| OmpW | 2F1V/2F1T | 21 | Monomer | 8 | 17 | 23 | 4 Loops | [24,53] |
| OprD | 3SY7 | 48.4 | Monomer | 8 | 35 | 34 | 4 loops | [40] |
| OmpT | 6EHD | 40 | Trimeric | 10 | 33 | 42 | 8 Loops | [39] |
| OmpG | 2F1C | 35 | Monomer | 14 | 17 | 48 | 7 loops | [31] |
| OmpF | 2ZFG | 40 | Trimer | 16 | 33 | 42 | 8 Loops | [41] |
| OmpC | 2J1N | 40.4 | Trimer | 16 | 32 | 35 | 8 loops | [21] |
| PhoE | 1PHO | 39.5 | Trimer | 16 | 32 | 39 | 8 loops | [54] |
| Maltoporin | 1AF6 | 49.9 | Trimer | 18 | 37 | 35 | 9 loops | [55] |
| FhuA | 1BY3 | 82 | Monomer | 22 | 41 | 31 | 1 Cork & 11 loops | [17] |
| PapC | 3FIP | 91.5 | Dimer | 24 | 42 | 36 | 1 cork & 12 loops | [18] |
| TolC | 7NG9 | 162 | Trimer | 6 | 32 | 128 | 6 loops | [42] |
| Proteins | PDB Code | Average Molecular Weight(kDa) | Oligomeric State | Number of β-Strands per Monomer | Average Internal Diameter (Å) | Length of Barrel (Å) | Corks and Loops | Citation |
|---|---|---|---|---|---|---|---|---|
| VDAC | 6TIQ | 31 | Dynamic (Dimer, Trimer, Tetramer) | 19 | 33 | 37 | 9 loops | [43] |
| TOM complex | 7VD2 | 38 | Dimer | 19 | 35 | 17 | 9 loops | [44] |
| Fhac/Sam50 | 4QKY | 54.4 | Hexamer | 16 | 32 | 29 | 8 loops | [45,46,47] |
| Toxins | PDB | Average Molecular Weight (kDa) | Number of Chains | Internal Diameter (Å) | Length of Barrel (Å) | Number of β-Strands | Citation |
|---|---|---|---|---|---|---|---|
| Cytolysin (Sticholysin II) | 1GWY | 19.3 | 1 | 14 | 23 | 10 | [56] |
| α-hemolysin | 3ANZ | 33 | 7 | 26 | 52 | 14 | [48,49,50] |
| γ-hemolysin | 3B07 | 36.7 | 8 | 28 | 47 | 16 | [9] |
| Bi-component Toxin LukGH | 4TW1 | 36.8 | 8 | 30 | 39 | 16 | [57] |
| Aerolysin | 5JZH/5JZW | 54.3 | 7 | 28 | 19 | 14 | [12] |
| Epsilon toxin | 6RB9 | 36.3 | 7 | 30 | 68 | 14 | [58] |
| Anthrax Toxin | 6PSN | 90 | 7 | 26 | 93 | 14 | [51] |
| Lysenin | 5GAQ | 33.4 | 9 | 34 | 85 | 18 | [59] |
| MspA | 1UUN | 22.1 | 8 | 48 | 37 | 16 | [52] |
2. Early Observations of Voltage Gating of β Barrels
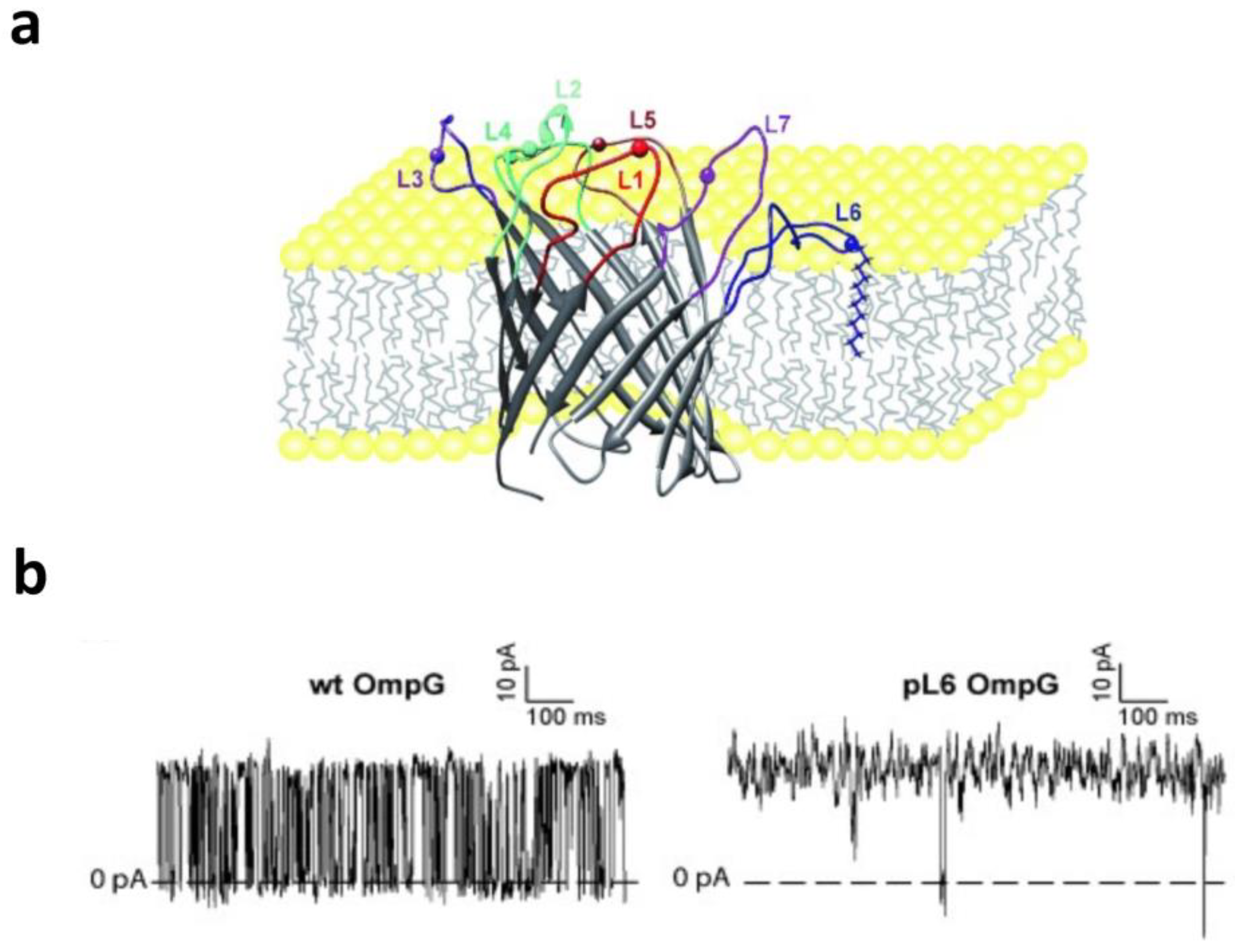
3. Gating Activity Produced by Loops and Plugs
4. Gating Activity Modulated by the N-Terminal Tail
5. Modulation of the Voltage-Dependent Gating by Environmental Conditions
5.1. Effect of pH
5.2. Effect of Temperature

5.3. Effect of Lipid Composition and Bilayer Asymmetry
6. Applications in Biotechnology
7. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pocanschi, C.L.; Kleinschmidt, J.H. The Thermodynamic Stability of Membrane Proteins in Micelles and Lipid Bilayers Investigated with the Ferrichrom Receptor FhuA. J. Membr. Biol. 2022, 255, 485–502. [Google Scholar] [CrossRef] [PubMed]
- Horne, J.E.; Radford, S.E. A growing toolbox of techniques for studying β-barrel outer membrane protein folding and biogenesis. Biochem. Soc. Trans. 2016, 44, 802–809. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, D.; Mahalakshmi, R. Transmembrane β-barrels: Evolution, folding and energetics. Biochim. Biophys. Acta Biomembr. 2017, 1859, 2467–2482. [Google Scholar] [CrossRef] [PubMed]
- Slusky, J.S. Outer membrane protein design. Curr. Opin. Struct. Biol. 2017, 45, 45–52. [Google Scholar] [CrossRef]
- Thoma, J.; Sapra, K.T.; Müller, D.J. Single-Molecule Force Spectroscopy of Transmembrane β-Barrel Proteins. Annu. Rev. Anal. Chem. 2018, 11, 375–395. [Google Scholar] [CrossRef] [PubMed]
- Vergalli, J.; Bodrenko, I.V.; Masi, M.; Moynié, L.; Acosta-Gutiérrez, S.; Naismith, J.H.; Davin-Regli, A.; Ceccarelli, M.; van den Berg, B.; Winterhalter, M.; et al. Porins and small-molecule translocation across the outer membrane of Gram-negative bacteria. Nat. Rev. Microbiol. 2020, 18, 164–176. [Google Scholar] [CrossRef]
- Hermansen, S.; Linke, D.; Leo, J.C. Transmembrane β-barrel proteins of bacteria: From structure to function. Adv. Protein Chem. Struct. Biol. 2022, 128, 113–161. [Google Scholar] [CrossRef]
- Sayyed, U.M.H.; Mahalakshmi, R. Mitochondrial protein translocation machinery: From TOM structural biogenesis to functional regulation. J. Biol. Chem. 2022, 298, 101870. [Google Scholar] [CrossRef]
- Yamashita, K.; Kawai, Y.; Tanaka, Y.; Hirano, N.; Kaneko, J.; Tomita, N.; Ohta, M.; Kamio, Y.; Yao, M.; Tanaka, I. Crystal structure of the octameric pore of staphylococcal γ-hemolysin reveals the β-barrel pore formation mechanism by two components. Proc. Natl. Acad. Sci. USA 2011, 108, 17314–17319. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, D.; Sugawara, T.; Takeshita, M.; Kaneko, J.; Kamio, Y.; Tanaka, I.; Tanaka, Y.; Yao, M. Molecular basis of transmembrane beta-barrel formation of staphylococcal pore-forming toxins. Nat. Commun. 2014, 5, 4897. [Google Scholar] [CrossRef] [Green Version]
- Dal Peraro, M.; van der Goot, F.G. Pore-forming toxins: Ancient, but never really out of fashion. Nat. Rev. Microbiol. 2016, 14, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Iacovache, I.; De Carlo, S.; Cirauqui, N.; Dal Peraro, M.; van der Goot, F.G.; Zuber, B. Cryo-EM structure of aerolysin variants reveals a novel protein fold and the pore-formation process. Nat. Commun. 2016, 7, 12062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, M.W.; Slusky, J.S.G. Tight Turns of Outer Membrane Proteins: An Analysis of Sequence, Structure, and Hydrogen Bonding. J. Mol. Biol. 2018, 430, 3251–3265. [Google Scholar] [CrossRef]
- Kleinschmidt, J.H. Folding of β-barrel membrane proteins in lipid bilayers—Unassisted and assisted folding and insertion. Biochim. Biophys. Acta 2015, 1848, 1927–1943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pautsch, A.; Schulz, G.E. High-resolution structure of the OmpA membrane domain. J. Mol. Biol. 2000, 298, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, A.D.; Hofmann, E.; Coulton, J.W.; Diederichs, K.; Welte, W. Siderophore-mediated iron transport: Crystal structure of FhuA with bound lipopolysaccharide. Science 1998, 282, 2215–2220. [Google Scholar] [CrossRef] [Green Version]
- Locher, K.P.; Rees, B.; Koebnik, R.; Mitschler, A.; Moulinier, L.; Rosenbusch, J.P.; Moras, D. Transmembrane signaling across the ligand-gated FhuA receptor: Crystal structures of free and ferrichrome-bound states reveal allosteric changes. Cell 1998, 95, 771–778. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Smith, B.S.; Chen, L.X.; Baxter, R.H.; Deisenhofer, J. Insights into pilus assembly and secretion from the structure and functional characterization of usher PapC. Proc. Natl. Acad. Sci. USA 2009, 106, 7403–7407. [Google Scholar] [CrossRef]
- Cowan, S.W.; Garavito, R.M.; Jansonius, J.N.; Jenkins, J.A.; Karlsson, R.; Konig, N.; Pai, E.F.; Pauptit, R.A.; Rizkallah, P.J.; Rosenbusch, J.P.; et al. The structure of OmpF porin in a tetragonal crystal form. Structure 1995, 3, 1041–1050. [Google Scholar] [CrossRef]
- Pebay-Peyroula, E.; Garavito, R.M.; Rosenbusch, J.P.; Zulauf, M.; Timmins, P.A. Detergent structure in tetragonal crystals of OmpF porin. Structure 1995, 3, 1051–1059. [Google Scholar] [CrossRef] [Green Version]
- Basle, A.; Rummel, G.; Storici, P.; Rosenbusch, J.P.; Schirmer, T. Crystal structure of osmoporin OmpC from E. coli at 2.0 A. J. Mol. Biol. 2006, 362, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Koronakis, V.; Sharff, A.; Koronakis, E.; Luisi, B.; Hughes, C. Crystal structure of the bacterial membrane protein TolC central to multidrug efflux and protein export. Nature 2000, 405, 914–919. [Google Scholar] [CrossRef]
- Buchanan, S.K. Type I secretion and multidrug efflux: Transport through the TolC channel-tunnel. Trends Biochem. Sci. 2001, 26, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Patel, D.R.; Tamm, L.K.; van den Berg, B. The outer membrane protein OmpW forms an eight-stranded beta-barrel with a hydrophobic channel. J. Biol. Chem. 2006, 281, 7568–7577. [Google Scholar] [CrossRef] [Green Version]
- Horst, R.; Stanczak, P.; Wüthrich, K. NMR polypeptide backbone conformation of the E. coli outer membrane protein W. Structure 2014, 22, 1204–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, J.; Schulz, G.E. The structure of the outer membrane protein OmpX from Escherichia coli reveals possible mechanisms of virulence. Structure 1999, 7, 1301–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, V.E.; Lo, E.I.; Engel, C.K.; Chen, L.; Hwang, P.M.; Kay, L.E.; Bishop, R.E.; Privé, G.G. A hydrocarbon ruler measures palmitate in the enzymatic acylation of endotoxin. EMBO J. 2004, 23, 2931–2941. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Suarez, M.L.; Samsudin, F.; Piggot, T.J.; Bond, P.J.; Khalid, S. Full-Length OmpA: Structure, Function, and Membrane Interactions Predicted by Molecular Dynamics Simulations. Biophys. J. 2016, 111, 1692–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Bernstein, H.D. The Escherichia coli outer membrane protein OmpA acquires secondary structure prior to its integration into the membrane. J. Biol. Chem. 2022, 298, 101802. [Google Scholar] [CrossRef]
- Vandeputte-Rutten, L.; Kramer, R.A.; Kroon, J.; Dekker, N.; Egmond, M.R.; Gros, P. Crystal structure of the outer membrane protease OmpT from Escherichia coli suggests a novel catalytic site. EMBO J. 2001, 20, 5033–5039. [Google Scholar] [CrossRef] [Green Version]
- Subbarao, G.V.; van den Berg, B. Crystal structure of the monomeric porin OmpG. J. Mol. Biol 2006, 360, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, O.; Vinothkumar, K.R.; Goswami, P.; Kuhlbrandt, W. Structure of the monomeric outer-membrane porin OmpG in the open and closed conformation. EMBO J. 2006, 25, 3702–3713. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Tamm, L.K. Structure of outer membrane protein G by solution NMR spectroscopy. Proc. Natl. Acad. Sci. USA 2007, 104, 16140–16145. [Google Scholar] [CrossRef] [PubMed]
- Botos, I.; Majdalani, N.; Mayclin, S.J.; McCarthy, J.G.; Lundquist, K.; Wojtowicz, D.; Barnard, T.J.; Gumbart, J.C.; Buchanan, S.K. Structural and Functional Characterization of the LPS Transporter LptDE from Gram-Negative Pathogens. Structure 2016, 24, 965–976. [Google Scholar] [CrossRef] [Green Version]
- Lauber, F.; Deme, J.C.; Lea, S.M.; Berks, B.C. Type 9 secretion system structures reveal a new protein transport mechanism. Nature 2018, 564, 77–82. [Google Scholar] [CrossRef]
- Biswas, S.; Mohammad, M.M.; Patel, D.R.; Movileanu, L.; van den Berg, B. Structural insight into OprD substrate specificity. Nat. Struct. Mol. Biol. 2007, 14, 1108–1109. [Google Scholar] [CrossRef]
- Biswas, S.; Mohammad, M.M.; Movileanu, L.; van den Berg, B. Crystal structure of the outer membrane protein OpdK from Pseudomonas aeruginosa. Structure 2008, 16, 1027–1035. [Google Scholar] [CrossRef] [Green Version]
- Snijder, H.J.; Ubarretxena-Belandia, I.; Blaauw, M.; Kalk, K.H.; Verheij, H.M.; Egmond, M.R.; Dekker, N.; Dijkstra, B.W. Structural evidence for dimerization-regulated activation of an integral membrane phospholipase. Nature 1999, 401, 717–721. [Google Scholar] [CrossRef]
- Pathania, M.; Acosta-Gutierrez, S.; Bhamidimarri, S.P.; Baslé, A.; Winterhalter, M.; Ceccarelli, M.; van den Berg, B. Unusual Constriction Zones in the Major Porins OmpU and OmpT from Vibrio cholerae. Structure 2018, 26, 708–721.e704. [Google Scholar] [CrossRef] [Green Version]
- Eren, E.; Vijayaraghavan, J.; Liu, J.; Cheneke, B.R.; Touw, D.S.; Lepore, B.W.; Indic, M.; Movileanu, L.; van den Berg, B. Substrate specificity within a family of outer membrane carboxylate channels. PLoS Biol. 2012, 10, e1001242. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, E.; Zhalnina, M.V.; Zakharov, S.D.; Sharma, O.; Cramer, W.A. Crystal structures of the OmpF porin: Function in a colicin translocon. EMBO J. 2008, 27, 2171–2180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Housden, N.G.; Webby, M.N.; Lowe, E.D.; El-Baba, T.J.; Kaminska, R.; Redfield, C.; Robinson, C.V.; Kleanthous, C. Toxin import through the antibiotic efflux channel TolC. Nat. Commun. 2021, 12, 4625. [Google Scholar] [CrossRef] [PubMed]
- Böhm, R.; Amodeo, G.F.; Murlidaran, S.; Chavali, S.; Wagner, G.; Winterhalter, M.; Brannigan, G.; Hiller, S. The Structural Basis for Low Conductance in the Membrane Protein VDAC upon β-NADH Binding and Voltage Gating. Structure 2020, 28, 206–214.e204. [Google Scholar] [CrossRef]
- Su, J.; Liu, D.; Yang, F.; Zuo, M.Q.; Li, C.; Dong, M.Q.; Sun, S.; Sui, S.F. Structural basis of Tom20 and Tom22 cytosolic domains as the human TOM complex receptors. Proc. Natl. Acad. Sci. USA 2022, 119, e2200158119. [Google Scholar] [CrossRef] [PubMed]
- Höhr, A.I.; Straub, S.P.; Warscheid, B.; Becker, T.; Wiedemann, N. Assembly of β-barrel proteins in the mitochondrial outer membrane. Biochim. Biophys. Acta 2015, 1853, 74–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Höhr, A.I.C.; Lindau, C.; Wirth, C.; Qiu, J.; Stroud, D.A.; Kutik, S.; Guiard, B.; Hunte, C.; Becker, T.; Pfanner, N.; et al. Membrane protein insertion through a mitochondrial β-barrel gate. Science 2018, 359, 6373. [Google Scholar] [CrossRef] [Green Version]
- Maier, T.; Clantin, B.; Gruss, F.; Dewitte, F.; Delattre, A.S.; Jacob-Dubuisson, F.; Hiller, S.; Villeret, V. Conserved Omp85 lid-lock structure and substrate recognition in FhaC. Nat. Commun. 2015, 6, 7452. [Google Scholar] [CrossRef] [Green Version]
- Song, L.Z.; Hobaugh, M.R.; Shustak, C.; Cheley, S.; Bayley, H.; Gouaux, J.E. Structure of Staphylococcal Alpha-Hemolysin, a Heptameric Transmembrane Pore. Science 1996, 274, 1859–1866. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Hirano, N.; Kaneko, J.; Kamio, Y.; Yao, M.; Tanaka, I. 2-Methyl-2,4-pentanediol induces spontaneous assembly of staphylococcal α-hemolysin into heptameric pore structure. Protein Sci. 2011, 20, 448–456. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, T.; Yamashita, D.; Kato, K.; Peng, Z.; Ueda, J.; Kaneko, J.; Kamio, Y.; Tanaka, Y.; Yao, M. Structural basis for pore-forming mechanism of staphylococcal α-hemolysin. Toxicon 2015, 108, 226–231. [Google Scholar] [CrossRef] [Green Version]
- Hardenbrook, N.J.; Liu, S.; Zhou, K.; Ghosal, K.; Hong Zhou, Z.; Krantz, B.A. Atomic structures of anthrax toxin protective antigen channels bound to partially unfolded lethal and edema factors. Nat. Commun. 2020, 11, 840. [Google Scholar] [CrossRef]
- Faller, M.; Niederweis, M.; Schulz, G.E. The structure of a mycobacterial outer-membrane channel. Science 2004, 303, 1189–1192. [Google Scholar] [CrossRef] [PubMed]
- Pali, T.; Marsh, D. Tilt, twist, and coiling in beta-barrel membrane proteins: Relation to infrared dichroism. Biophys. J. 2001, 80, 2789–2797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowan, S.W.; Schirmer, T.; Rummel, G.; Steiert, M.; Ghosh, R.; Pauptit, R.A.; Jansonius, J.N.; Rosenbusch, J.P. Crystal structures explain functional properties of two E. coli porins. Nature 1992, 358, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.F.; Dutzler, R.; Rizkallah, P.J.; Rosenbusch, J.P.; Schirmer, T. Channel specificity: Structural basis for sugar discrimination and differential flux rates in maltoporin. J. Mol. Biol. 1997, 272, 56–63. [Google Scholar] [CrossRef]
- Mancheño, J.M.; Martín-Benito, J.; Martínez-Ripoll, M.; Gavilanes, J.G.; Hermoso, J.A. Crystal and electron microscopy structures of sticholysin II actinoporin reveal insights into the mechanism of membrane pore formation. Structure 2003, 11, 1319–1328. [Google Scholar] [CrossRef] [Green Version]
- Badarau, A.; Rouha, H.; Malafa, S.; Logan, D.T.; Håkansson, M.; Stulik, L.; Dolezilkova, I.; Teubenbacher, A.; Gross, K.; Maierhofer, B.; et al. Structure-function analysis of heterodimer formation, oligomerization, and receptor binding of the Staphylococcus aureus bi-component toxin LukGH. J. Biol. Chem. 2015, 290, 142–156. [Google Scholar] [CrossRef] [Green Version]
- Savva, C.G.; Clark, A.R.; Naylor, C.E.; Popoff, M.R.; Moss, D.S.; Basak, A.K.; Titball, R.W.; Bokori-Brown, M. The pore structure of Clostridium perfringens epsilon toxin. Nat. Commun. 2019, 10, 2641. [Google Scholar] [CrossRef] [Green Version]
- Bokori-Brown, M.; Martin, T.G.; Naylor, C.E.; Basak, A.K.; Titball, R.W.; Savva, C.G. Cryo-EM structure of lysenin pore elucidates membrane insertion by an aerolysin family protein. Nat. Commun. 2016, 7, 11293. [Google Scholar] [CrossRef] [Green Version]
- Hiller, S.; Garces, R.G.; Malia, T.J.; Orekhov, V.Y.; Colombini, M.; Wagner, G. Solution structure of the integral human membrane protein VDAC-1 in detergent micelles. Science 2008, 321, 1206–1210. [Google Scholar] [CrossRef] [Green Version]
- Colombini, M. The VDAC channel: Molecular basis for selectivity. Biochim. Biophys. Acta 2016, 1863, 2498–2502. [Google Scholar] [CrossRef] [PubMed]
- Naghdi, S.; Hajnóczky, G. VDAC2-specific cellular functions and the underlying structure. Biochim. Biophys. Acta 2016, 1863, 2503–2514. [Google Scholar] [CrossRef] [PubMed]
- Zeth, K.; Zachariae, U. Ten Years of High Resolution Structural Research on the Voltage Dependent Anion Channel (VDAC)-Recent Developments and Future Directions. Front. Physiol. 2018, 9, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benz, R. Historical Perspective of Pore-Forming Activity Studies of Voltage-Dependent Anion Channel (Eukaryotic or Mitochondrial Porin) Since Its Discovery in the 70th of the Last Century. Front. Physiol. 2021, 12, 734226. [Google Scholar] [CrossRef] [PubMed]
- Reina, S.; Checchetto, V. Voltage-Dependent Anion Selective Channel 3: Unraveling Structural and Functional Features of the Least Known Porin Isoform. Front. Physiol. 2021, 12, 784867. [Google Scholar] [CrossRef]
- Zinghirino, F.; Pappalardo, X.G.; Messina, A.; Nicosia, G.; De Pinto, V.; Guarino, F. VDAC Genes Expression and Regulation in Mammals. Front. Physiol. 2021, 12, 708695. [Google Scholar] [CrossRef]
- Ulrich, T.; Gross, L.E.; Sommer, M.S.; Schleiff, E.; Rapaport, D. Chloroplast β-barrel proteins are assembled into the mitochondrial outer membrane in a process that depends on the TOM and TOB complexes. J. Biol. Chem. 2012, 287, 27467–27479. [Google Scholar] [CrossRef] [Green Version]
- Jores, T.; Rapaport, D. Early stages in the biogenesis of eukaryotic β-barrel proteins. FEBS Lett. 2017, 591, 2671–2681. [Google Scholar] [CrossRef]
- Day, P.M.; Inoue, K.; Theg, S.M. Chloroplast Outer Membrane β-Barrel Proteins Use Components of the General Import Apparatus. Plant Cell 2019, 31, 1845–1855. [Google Scholar] [CrossRef]
- Gross, L.E.; Klinger, A.; Spies, N.; Ernst, T.; Flinner, N.; Simm, S.; Ladig, R.; Bodensohn, U.; Schleiff, E. Insertion of plastidic β-barrel proteins into the outer envelopes of plastids involves an intermembrane space intermediate formed with Toc75-V/OEP80. Plant Cell 2021, 33, 1657–1681. [Google Scholar] [CrossRef]
- Jiang, J.; Pentelute, B.L.; Collier, R.J.; Zhou, Z.H. Atomic structure of anthrax protective antigen pore elucidates toxin translocation. Nature 2015, 521, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Alonzo, F., III; Torres, V.J. The bicomponent pore-forming leucocidins of Staphylococcus aureus. Microbiol. Mol. Biol. Rev. 2014, 78, 199–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaan, A.N.; van Strijp, J.A.G.; Torres, V.J. Leukocidins: Staphylococcal bi-component pore-forming toxins find their receptors. Nat. Rev. Microbiol. 2017, 15, 435–447. [Google Scholar] [CrossRef]
- Tromp, A.T.; van Strijp, J.A.G. Studying Staphylococcal Leukocidins: A Challenging Endeavor. Front. Microbiol. 2020, 11, 611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miles, G.; Movileanu, L.; Bayley, H. Subunit composition of a bicomponent toxin: Staphylococcal leukocidin forms an octameric transmembrane pore. Protein Sci. 2002, 11, 894–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gugel, J.F.; Movileanu, L. Staphylococcal beta-barrel Pore-Forming Toxins: Mushrooms That Breach the Greasy Barrier. In Electrophysiology of Uncoventional Channels and Pores; Delcour, A.H., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; Volume 18, pp. 241–266. [Google Scholar]
- Goyal, P.; Krasteva, P.V.; Van Gerven, N.; Gubellini, F.; Van den Broeck, I.; Troupiotis-Tsailaki, A.; Jonckheere, W.; Pehau-Arnaudet, G.; Pinkner, J.S.; Chapman, M.R.; et al. Structural and mechanistic insights into the bacterial amyloid secretion channel CsgG. Nature 2014, 516, 250–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Z.; Yin, M.; Xu, D.; Zhu, Y.; Li, X. Structural insights into the secretin translocation channel in the type II secretion system. Nat. Struct. Mol. Biol. 2017, 24, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.J.; Ortega, D.R.; Sazinsky, M.H.; Dalia, T.N.; Dalia, A.B.; Jensen, G.J. CryoEM structure of the type IVa pilus secretin required for natural competence in Vibrio cholerae. Nat. Commun. 2020, 11, 5080. [Google Scholar] [CrossRef]
- Phale, P.S.; Schirmer, T.; Prilipov, A.; Lou, K.L.; Hardmeyer, A.; Rosenbusch, J.P. Voltage gating of Escherichia coli porin channels: Role of the constriction loop. Proc. Natl. Acad. Sci. USA 1997, 94, 6741–6745. [Google Scholar] [CrossRef]
- Van Gelder, P.; Saint, N.; Phale, P.; Eppens, E.F.; Prilipov, A.; van Boxtel, R.; Rosenbusch, J.P.; Tommassen, J. Voltage sensing in the PhoE and OmpF outer membrane porins of Escherichia coli: Role of charged residues. J. Mol. Biol. 1997, 269, 468–472. [Google Scholar] [CrossRef]
- Bainbridge, G.; Gokce, I.; Lakey, J.H. Voltage gating is a fundamental feature of porin and toxin beta-barrel membrane channels. FEBS Lett. 1998, 431, 305–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bainbridge, G.; Mobasheri, H.; Armstrong, G.A.; Lea, E.J.A.; Lakey, J.H. Voltage-gating of Escherichia coli porin: A cystine-scanning mutagenesis study of loop 3. J. Mol. Biol. 1998, 275, 171–176. [Google Scholar] [CrossRef]
- Mathes, A.; Engelhardt, H. Voltage-dependent closing of porin channels: Analysis of relaxation kinetics. J. Membr. Biol. 1998, 165, 11–18. [Google Scholar] [CrossRef]
- Mathes, A.; Engelhardt, H. Nonlinear and asymmetric open channel characteristics of an ion-selective porin in planar membranes. Biophys. J. 1998, 75, 1255–1262. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Delcour, A.H. The spontaneous gating activity of OmpC porin is affected by mutations of a putative hydrogen bond network or of a salt bridge between the L3 loop and the barrel. Protein Eng. 1998, 11, 797–802. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Delcour, A.H. Inhibitory effect of acidic pH on OmpC porin: Wild-type and mutant studies. FEBS Lett. 1998, 434, 160–164. [Google Scholar] [CrossRef] [Green Version]
- Samartzidou, H.; Delcour, A.H. E. coli PhoE porin has an opposite voltage-dependence to the homologous OmpF. EMBO J. 1998, 17, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basle, A.; Iyer, R.; Delcour, A.H. Subconductance states in OmpF gating. Biochim. Biophys. Acta 2004, 1664, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Rosenbusch, J.; Schirmer, T.; Karplus, M. Computer simulations of the OmpF porin from the outer membrane of Escherichia coli. Biophys. J. 1997, 72, 2094–2102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schirmer, T. General and specific porins from bacterial outer membranes. J. Struct. Biol. 1998, 121, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Phale, P.S.; Philippsen, A.; Kiefhaber, T.; Koebnik, R.; Phale, V.P.; Schirmer, T.; Rosenbusch, J.P. Stability of trimeric OmpF porin: The contributions of the latching loop L2. Biochemistry 1998, 37, 15663–15670. [Google Scholar] [CrossRef] [PubMed]
- Wager, B.; Basle, A.; Delcour, A.H. Disulfide bond tethering of extracellular loops does not affect the closure of OmpF porin at acidic pH. Proteins 2010, 78, 2886–2894. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Samartzidou, H.; Lee, K.W.; Briggs, J.M.; Delcour, A.H. Effects of pore mutations and permeant ion concentration on the spontaneous gating activity of OmpC porin. Protein Eng. 2000, 13, 491–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mobasheri, H.; Lea, E.J. Biophysics of gating phenomena in voltage-dependent OmpC mutant porin channels (R74C and R37C) of Escherichia coli outer membranes. Eur. Biophys. J. 2002, 31, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Eppens, E.F.; Saint, N.; Van Gelder, P.; van Boxtel, R.; Tommassen, J. Role of the constriction loop in the gating of outer membrane porin PhoE of Escherichia coli. FEBS Lett. 1997, 415, 317–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basle, A.; Qutub, R.; Mehrazin, M.; Wibbenmeyer, J.; Delcour, A.H. Deletions of single extracellular loops affect pH sensitivity, but not voltage dependence, of the Escherichia coli porin OmpF. Protein Eng. Des. Sel. 2004, 17, 665–672. [Google Scholar] [CrossRef]
- Alcaraz, A.; Nestorovich, E.M.; Aguilella-Arzo, M.; Aguilella, V.M.; Bezrukov, S.M. Salting out the ionic selectivity of a wide channel: The asymmetry of OmpF. Biophys. J. 2004, 87, 943–957. [Google Scholar] [CrossRef] [Green Version]
- Alcaraz, A.; Queralt-Martin, M.; Garcia-Gimenez, E.; Aguilella, V.M. Increased salt concentration promotes competitive block of OmpF channel by protons. Biochim. Biophys. Acta 2012, 1818, 2777–2782. [Google Scholar] [CrossRef] [Green Version]
- Rostovtseva, T.K.; Kazemi, N.; Weinrich, M.; Bezrukov, S.M. Voltage gating of VDAC is regulated by nonlamellar lipids of mitochondrial membranes. J. Biol. Chem. 2006, 281, 37496–37506. [Google Scholar] [CrossRef] [Green Version]
- Tomita, N.; Mohammad, M.M.; Niedzwiecki, D.J.; Ohta, M.; Movileanu, L. Does the lipid environment impact the open-state conductance of an engineered beta-barrel protein nanopore? Biochim. Biophys. Acta Biomembr. 2013, 1828, 1057–1065. [Google Scholar] [CrossRef] [Green Version]
- Hong, H.; Szabo, G.; Tamm, L.K. Electrostatic couplings in OmpA ion-channel gating suggest a mechanism for pore opening. Nat. Chem. Biol. 2006, 2, 627–635. [Google Scholar] [CrossRef]
- Robertson, K.M.; Tieleman, D.P. Molecular basis of voltage gating of OmpF porin. Biochem. Cell Biol. 2002, 80, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Tieleman, D.P.; Leontiadou, H.; Mark, A.E.; Marrink, S.J. Simulation of pore formation in lipid bilayers by mechanical stress and electric fields. J. Am. Chem. Soc. 2003, 125, 6382–6383. [Google Scholar] [CrossRef] [Green Version]
- Tieleman, D.P. Computer simulations of transport through membranes: Passive diffusion, pores, channels and transporters. Clin. Exp. Pharmacol. Physiol. 2006, 33, 893–903. [Google Scholar] [CrossRef]
- Khalid, S.; Bond, P.J.; Deol, S.S.; Sansom, M.S. Modeling and simulations of a bacterial outer membrane protein: OprF from Pseudomonas aeruginosa. Proteins 2006, 63, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Bond, P.J.; Derrick, J.P.; Sansom, M.S. Membrane simulations of OpcA: Gating in the loops? Biophys. J. 2007, 92, L23–L25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bond, P.J.; Holyoake, J.; Ivetac, A.; Khalid, S.; Sansom, M.S. Coarse-grained molecular dynamics simulations of membrane proteins and peptides. J. Struct. Biol. 2007, 157, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Khalid, S.; Bond, P.J.; Carpenter, T.; Sansom, M.S. OmpA: Gating and dynamics via molecular dynamics simulations. Biochim. Biophys. Acta 2008, 1778, 1871–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nestorovich, E.M.; Sugawara, E.; Nikaido, H.; Bezrukov, S.M. Pseudomonas aeruginosa porin OprF: Properties of the channel. J. Biol. Chem. 2006, 281, 16230–16237. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, E.; Nestorovich, E.M.; Bezrukov, S.M.; Nikaido, H. Pseudomonas aeruginosa porin OprF exists in two different conformations. J. Biol. Chem. 2006, 281, 16220–16229. [Google Scholar] [CrossRef] [Green Version]
- Conlan, S.; Bayley, H. Folding of a monomeric porin, OmpG, in detergent solution. Biochemistry 2003, 42, 9453–9465. [Google Scholar] [CrossRef] [PubMed]
- Conlan, S.; Zhang, Y.; Cheley, S.; Bayley, H. Biochemical and biophysical characterization of OmpG: A monomeric porin. Biochemistry 2000, 39, 11845–11854. [Google Scholar] [CrossRef] [PubMed]
- Levadny, V.; Colombini, M.; Li, X.X.; Aguilella, V.M. Electrostatics explains the shift in VDAC gating with salt activity gradient. Biophys. J. 2002, 82, 1773–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammad, M.M.; Movileanu, L. Impact of Distant Charge Reversals within a Robust Beta-Barrel Protein Pore. J. Phys. Chem. B 2010, I114, 8750–8759. [Google Scholar] [CrossRef] [Green Version]
- Damaghi, M.; Sapra, K.T.; Koster, S.; Yildiz, O.; Kuhlbrandt, W.; Muller, D.J. Dual energy landscape: The functional state of the beta-barrel outer membrane protein G molds its unfolding energy landscape. Proteomics 2010, 10, 4151–4162. [Google Scholar] [CrossRef]
- Damaghi, M.; Bippes, C.; Köster, S.; Yildiz, O.; Mari, S.A.; Kühlbrandt, W.; Muller, D.J. pH-dependent interactions guide the folding and gate the transmembrane pore of the beta-barrel membrane protein OmpG. J. Mol. Biol. 2010, 397, 878–882. [Google Scholar] [CrossRef]
- Runke, G.; Maier, E.; Summers, W.A.; Bay, D.C.; Benz, R.; Court, D.A. Deletion variants of Neurospora mitochondrial porin: Electrophysiological and spectroscopic analysis. Biophys. J. 2006, 90, 3155–3164. [Google Scholar] [CrossRef] [Green Version]
- Acharya, A.; Ghai, I.; Piselli, C.; Prajapati, J.D.; Benz, R.; Winterhalter, M.; Kleinekathöfer, U. Conformational Dynamics of Loop L3 in OmpF: Implications toward Antibiotic Translocation and Voltage Gating. J. Chem. Inf. Model. 2023, 63, 910–927. [Google Scholar] [CrossRef]
- Eren, E.; Parkin, J.; Adelanwa, A.; Cheneke, B.R.; Movileanu, L.; Khalid, S.; van den Berg, B. Towards understanding the outer membrane uptake of small molecules by Pseudomonas aeruginosa. J. Biol. Chem 2013, 288, 12042–12053. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Eren, E.; Vijayaraghavan, J.; Cheneke, B.R.; Indic, M.; van den Berg, B.; Movileanu, L. OccK Channels from Pseudomonas aeruginosa Exhibit Diverse Single-channel Electrical Signatures, but Conserved Anion Selectivity. Biochemistry 2012, 51, 2319–2330. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wolfe, A.J.; Eren, E.; Vijayaraghavan, J.; Indic, M.; van den Berg, B.; Movileanu, L. Cation Selectivity is a Conserved Feature in the OccD Subfamily of Pseudomonas aeruginosa. Biochim. Biophys. Acta Biomembr. 2012, 1818, 2908–2916. [Google Scholar] [CrossRef] [Green Version]
- Cheneke, B.R.; van den Berg, B.; Movileanu, L. Analysis of gating transitions among the three major open states of the OpdK channel. Biochemistry 2011, 50, 4987–4997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, M.; Yang, J. Engineered Ion Channels as Emerging Tools for Chemical Biology. Acc. Chem. Res. 2013, 46, 2998–3008. [Google Scholar] [CrossRef] [Green Version]
- Stoddart, D.; Ayub, M.; Hofler, L.; Raychaudhuri, P.; Klingelhoefer, J.W.; Maglia, G.; Heron, A.; Bayley, H. Functional truncated membrane pores. Proc. Natl. Acad. Sci. USA 2014, 111, 2425–2430. [Google Scholar] [CrossRef] [PubMed]
- Ayub, M.; Bayley, H. Engineered transmembrane pores. Curr. Opin. Chem. Biol. 2016, 34, 117–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, Y.L.; Hu, Z.L.; Zhang, S.; Qing, Y.; Fragasso, A.; Maglia, G.; Meller, A.; Bayley, H.; Dekker, C.; Long, Y.T. Nanopore-based technologies beyond DNA sequencing. Nat. Nanotechnol. 2022, 17, 1136–1146. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, T.; Tamm, L.K. Control of the Conductance of Engineered Protein Nanopores through Concerted Loop Motions. Angew. Chem. Int. Ed. Engl. 2014, 53, 5897–5902. [Google Scholar] [CrossRef] [PubMed]
- Sanganna Gari, R.R.; Montalvo-Acosta, J.J.; Heath, G.R.; Jiang, Y.; Gao, X.; Nimigean, C.M.; Chipot, C.; Scheuring, S. Correlation of membrane protein conformational and functional dynamics. Nat. Commun. 2021, 12, 4363. [Google Scholar] [CrossRef]
- Retel, J.S.; Nieuwkoop, A.J.; Hiller, M.; Higman, V.A.; Barbet-Massin, E.; Stanek, J.; Andreas, L.B.; Franks, W.T.; van Rossum, B.J.; Vinothkumar, K.R.; et al. Structure of outer membrane protein G in lipid bilayers. Nat. Commun. 2017, 8, 2073. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Khalid, S.; Sansom, M.S.; Bayley, H. Outer membrane protein G: Engineering a quiet pore for biosensing. Proc. Natl. Acad. Sci. USA 2008, 105, 6272–6277. [Google Scholar] [PubMed]
- Eisenberg, B. Engineering channels: Atomic biology. Proc. Natl. Acad. Sci. USA 2008, 105, 6211–6212. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, T.; Chisholm, C.; Chen, M.; Tamm, L.K. NMR-based conformational ensembles explain pH-gated opening and closing of OmpG channel. J. Am. Chem. Soc. 2013, 135, 15101–15113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosse, W.; Psakis, G.; Mertins, B.; Reiss, P.; Windisch, D.; Brademann, F.; Bürck, J.; Ulrich, A.; Koert, U.; Essen, L.O. Structure-based engineering of a minimal porin reveals loop-independent channel closure. Biochemistry 2014, 53, 4826–4838. [Google Scholar] [CrossRef]
- Villinger, S.; Briones, R.; Giller, K.; Zachariae, U.; Lange, A.; de Groot, B.L.; Griesinger, C.; Becker, S.; Zweckstetter, M. Functional dynamics in the voltage-dependent anion channel. Proc. Natl. Acad. Sci. USA 2010, 107, 22546–22551. [Google Scholar] [CrossRef]
- Zachariae, U.; Schneider, R.; Briones, R.; Gattin, Z.; Demers, J.P.; Giller, K.; Maier, E.; Zweckstetter, M.; Griesinger, C.; Becker, S.; et al. β-Barrel mobility underlies closure of the voltage-dependent anion channel. Structure 2012, 20, 1540–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosse, W.; Reiss, P.; Reitz, S.; Cebi, M.; Lübben, W.; Koert, U.; Essen, L.O. Structural and functional characterization of a synthetically modified OmpG. Bioorg. Med. Chem. 2010, 18, 7716–7723. [Google Scholar] [CrossRef] [PubMed]
- Grosse, W.; Essen, L.O.; Koert, U. Strategies and perspectives in ion-channel engineering. Chembiochem 2011, 12, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Fahie, M.; Chisholm, C.; Chen, M. Resolved single-molecule detection of individual species within a mixture of anti-biotin antibodies using an engineered monomeric nanopore. ACS Nano 2015, 9, 1089–1098. [Google Scholar] [CrossRef] [Green Version]
- Fahie, M.A.; Chen, M. Electrostatic Interactions between OmpG Nanopore and Analyte Protein Surface Can Distinguish between Glycosylated Isoforms. J. Phys. Chem. B 2015, 119, 10198–10206. [Google Scholar] [CrossRef]
- Fahie, M.A.; Yang, B.; Mullis, M.; Holden, M.A.; Chen, M. Selective Detection of Protein Homologues in Serum Using an OmpG Nanopore. Anal. Chem. 2015, 87, 11143–11149. [Google Scholar] [CrossRef] [Green Version]
- Fahie, M.A.; Yang, B.; Pham, B.; Chen, M. Tuning the selectivity and sensitivity of an OmpG nanopore sensor by adjusting ligand tether length. ACS Sens. 2016, 1, 614–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanganna Gari, R.R.; Seelheim, P.; Liang, B.; Tamm, L.K. Quiet Outer Membrane Protein G (OmpG) Nanopore for Biosensing. ACS Sens. 2019, 4, 1230–1235. [Google Scholar] [CrossRef] [PubMed]
- Killmann, H.; Benz, R.; Braun, V. Properties of the FhuA channel in the Escherichia coli outer membrane after deletion of FhuA portions within and outside the predicted gating loop. J. Bacteriol. 1996, 178, 6913–6920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, M.; Killmann, H.; Maier, E.; Benz, R.; Braun, V. Diffusion through channel derivatives of the Escherichia coli FhuA transport protein. Eur. J. Biochem. 2002, 269, 4948–4959. [Google Scholar] [CrossRef] [PubMed]
- Udho, E.; Jakes, K.S.; Buchanan, S.K.; James, K.J.; Jiang, X.; Klebba, P.E.; Finkelstein, A. Reconstitution of bacterial outer membrane TonB-dependent transporters in planar lipid bilayer membranes. Proc. Natl. Acad. Sci. USA 2009, 106, 21990–21995. [Google Scholar] [CrossRef]
- Mohammad, M.M.; Howard, K.R.; Movileanu, L. Redesign of a plugged beta-barrel membrane protein. J. Biol. Chem. 2011, 286, 8000–8013. [Google Scholar] [CrossRef] [Green Version]
- Udho, E.; Jakes, K.S.; Finkelstein, A. TonB-dependent transporter FhuA in planar lipid bilayers: Partial exit of its plug from the barrel. Biochemistry 2012, 51, 6753–6759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawelek, P.D.; Croteau, N.; Ng-Thow-Hing, C.; Khursigara, C.M.; Moiseeva, N.; Allaire, M.; Coulton, J.W. Structure of TonB in complex with FhuA, E. coli outer membrane receptor. Science 2006, 312, 1399–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, A.K.; Movileanu, L. Real-Time Measurement of Protein-Protein Interactions at Single-Molecule Resolution using a Biological Nanopore. Nat. Biotechnol. 2019, 37, 96–101. [Google Scholar] [CrossRef]
- Thakur, A.K.; Movileanu, L. Single-Molecule Protein Detection in a Biofluid Using a Quantitative Nanopore Sensor. ACS Sens. 2019, 4, 2320–2326. [Google Scholar] [CrossRef]
- Sun, J.; Thakur, A.K.; Movileanu, L. Protein Ligand-Induced Amplification in the 1/f Noise of a Protein-Selective Nanopore. Langmuir 2020, 36, 15247–15257. [Google Scholar] [CrossRef]
- Sun, J.; Thakur, A.K.; Movileanu, L. Current noise of a protein-selective biological nanopore. Proteomics 2021, 22, e2100077. [Google Scholar] [CrossRef]
- Mayse, L.A.; Imran, A.; Larimi, M.G.; Cosgrove, M.S.; Wolfe, A.J.; Movileanu, L. Disentangling the recognition complexity of a protein hub using a nanopore. Nat. Commun. 2022, 13, 978. [Google Scholar] [CrossRef]
- Ahmad, M.; Ha, J.H.; Mayse, L.A.; Presti, M.F.; Wolfe, A.J.; Moody, K.J.; Loh, S.N.; Movileanu, L. A generalizable nanopore sensor for highly specific protein detection at single-molecule precision. Nat. Commun. 2023, 14, 1374. [Google Scholar] [CrossRef]
- Mayse, L.A.; Imran, A.; Wang, Y.; Ahmad, M.; Oot, R.A.; Wilkens, S.; Movileanu, L. Evaluation of Nanopore Sensor Design Using Electrical and Optical Analyses. ACS Nano 2023, 17, 10857–10871. [Google Scholar] [CrossRef] [PubMed]
- Volkan, E.; Ford, B.A.; Pinkner, J.S.; Dodson, K.W.; Henderson, N.S.; Thanassi, D.G.; Waksman, G.; Hultgren, S.J. Domain activities of PapC usher reveal the mechanism of action of an Escherichia coli molecular machine. Proc. Natl. Acad. Sci. USA 2012, 109, 9563–9568. [Google Scholar]
- Farabella, I.; Pham, T.; Henderson, N.S.; Geibel, S.; Phan, G.; Thanassi, D.G.; Delcour, A.H.; Waksman, G.; Topf, M. Allosteric signalling in the outer membrane translocation domain of PapC usher. eLife 2014, 3, 03532. [Google Scholar] [CrossRef] [Green Version]
- Mapingire, O.S.; Henderson, N.S.; Duret, G.; Thanassi, D.G.; Delcour, A.H. Modulating effects of the plug, helix, and N- and C-terminal domains on channel properties of the PapC usher. J. Biol. Chem. 2009, 284, 36324–36333. [Google Scholar] [CrossRef] [Green Version]
- Volkan, E.; Kalas, V.; Pinkner, J.S.; Dodson, K.W.; Henderson, N.S.; Pham, T.; Waksman, G.; Delcour, A.H.; Thanassi, D.G.; Hultgren, S.J. Molecular basis of usher pore gating in Escherichia coli pilus biogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, 20741–20746. [Google Scholar] [PubMed]
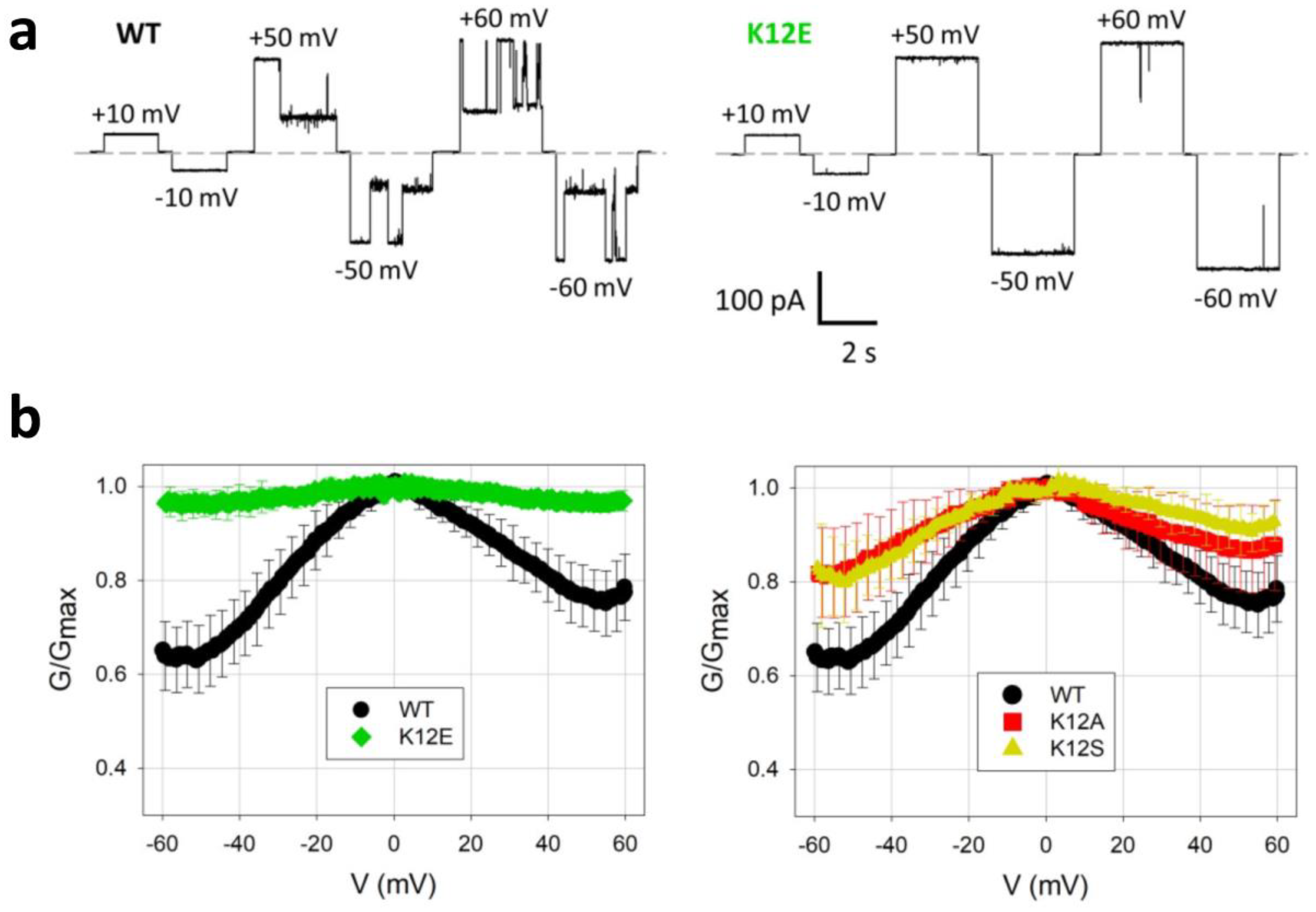
- Ngo, V.A.; Queralt-Martín, M.; Khan, F.; Bergdoll, L.; Abramson, J.; Bezrukov, S.M.; Rostovtseva, T.K.; Hoogerheide, D.P.; Noskov, S.Y. The Single Residue K12 Governs the Exceptional Voltage Sensitivity of Mitochondrial Voltage-Dependent Anion Channel Gating. J. Am. Chem. Soc. 2022, 144, 14564–14577. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, O.P.; Ujwal, R.; Kowallis, W.; Coalson, R.; Abramson, J.; Grabe, M. The electrostatics of VDAC: Implications for selectivity and gating. J. Mol. Biol. 2010, 396, 580–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurnev, P.A.; Rostovtseva, T.K.; Bezrukov, S.M. Tubulin-blocked state of VDAC studied by polymer and ATP partitioning. FEBS Lett. 2011, 585, 2363–2366. [Google Scholar] [CrossRef] [Green Version]
- Gurnev, P.A.; Queralt-Martin, M.; Aguilella, V.M.; Rostovtseva, T.K.; Bezrukov, S.M. Probing Tubulin-Blocked State of VDAC by Varying Membrane Surface Charge. Biophys. J. 2012, 102, 2070–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rostovtseva, T.K.; Bezrukov, S.M. VDAC inhibition by tubulin and its physiological implications. Biochim. Biophys. Acta 2012, 1818, 1526–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombini, M. VDAC structure, selectivity, and dynamics. Biochim. Biophys. Acta 2012, 1818, 1457–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villinger, S.; Giller, K.; Bayrhuber, M.; Lange, A.; Griesinger, C.; Becker, S.; Zweckstetter, M. Nucleotide interactions of the human voltage-dependent anion channel. J. Biol. Chem. 2014, 289, 13397–13406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vander Heiden, M.G.; Li, X.X.; Gottleib, E.; Hill, R.B.; Thompson, C.B.; Colombini, M. Bcl-xL promotes the open configuration of the voltage-dependent anion channel and metabolite passage through the outer mitochondrial membrane. J. Biol. Chem. 2001, 276, 19414–19419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rostovtseva, T.K.; Antonsson, B.; Suzuki, M.; Youle, R.J.; Colombini, M.; Bezrukov, S.M. Bid, but not Bax, regulates VDAC channels. J. Biol. Chem. 2004, 279, 13575–13583. [Google Scholar] [CrossRef] [Green Version]
- Noskov, S.Y.; Rostovtseva, T.K.; Chamberlin, A.C.; Teijido, O.; Jiang, W.; Bezrukov, S.M. Current state of theoretical and experimental studies of the voltage-dependent anion channel (VDAC). Biochim. Biophys. Acta 2016, 1858, 1778–1790. [Google Scholar] [CrossRef]
- Rostovtseva, T.K.; Gurnev, P.A.; Hoogerheide, D.P.; Rovini, A.; Sirajuddin, M.; Bezrukov, S.M. Sequence diversity of tubulin isotypes in regulation of the mitochondrial voltage-dependent anion channel. J. Biol. Chem. 2018, 293, 10949–10962. [Google Scholar] [CrossRef] [Green Version]
- Colombini, M. The published 3D structure of the VDAC channel: Native or not? Trends Biochem. Sci. 2009, 34, 382–389. [Google Scholar] [CrossRef]
- Hiller, S.; Abramson, J.; Mannella, C.; Wagner, G.; Zeth, K. The 3D structures of VDAC represent a native conformation. Trends Biochem. Sci. 2010, 35, 514–521. [Google Scholar] [CrossRef] [Green Version]
- Zeth, K. Structure and evolution of mitochondrial outer membrane proteins of beta-barrel topology. Biochim. Biophys. Acta 2010, 1797, 1292–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martynowycz, M.W.; Khan, F.; Hattne, J.; Abramson, J.; Gonen, T. MicroED structure of lipid-embedded mammalian mitochondrial voltage-dependent anion channel. Proc. Natl. Acad. Sci. USA 2020, 117, 32380–32385. [Google Scholar] [CrossRef] [PubMed]
- Najbauer, E.E.; Tekwani Movellan, K.; Giller, K.; Benz, R.; Becker, S.; Griesinger, C.; Andreas, L.B. Structure and Gating Behavior of the Human Integral Membrane Protein VDAC1 in a Lipid Bilayer. J. Am. Chem. Soc. 2022, 144, 2953–2967. [Google Scholar] [CrossRef] [PubMed]
- Briones, R.; Weichbrodt, C.; Paltrinieri, L.; Mey, I.; Villinger, S.; Giller, K.; Lange, A.; Zweckstetter, M.; Griesinger, C.; Becker, S.; et al. Voltage Dependence of Conformational Dynamics and Subconducting States of VDAC-1. Biophys. J. 2016, 111, 1223–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, L.; Villinger, S.; Mari, S.A.; Giller, K.; Griesinger, C.; Becker, S.; Müller, D.J.; Zweckstetter, M. Molecular Plasticity of the Human Voltage-Dependent Anion Channel Embedded Into a Membrane. Structure 2016, 24, 585–594. [Google Scholar] [CrossRef] [Green Version]
- Shuvo, S.R.; Ferens, F.G.; Court, D.A. The N-terminus of VDAC: Structure, Mutational Analysis, and a Potential Role in Regulating Barrel Shape. Biochim. Biophys. Acta 2016, 1858, 1350–1361. [Google Scholar] [CrossRef] [PubMed]
- Shuvo, S.R.; Kovaltchouk, U.; Zubaer, A.; Kumar, A.; Summers, W.A.T.; Donald, L.J.; Hausner, G.; Court, D.A. Functional characterization of an N-terminally-truncated mitochondrial porin expressed in Neurospora crassa. Can. J. Microbiol. 2017, 63, 730–738. [Google Scholar] [CrossRef]
- Reif, M.M.; Fischer, M.; Fredriksson, K.; Hagn, F.; Zacharias, M. The N-Terminal Segment of the Voltage-Dependent Anion Channel: A Possible Membrane-Bound Intermediate in Pore Unbinding. J. Mol. Biol. 2019, 431, 223–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Pinto, V. Renaissance of VDAC: New Insights on a Protein Family at the Interface between Mitochondria and Cytosol. Biomolecules 2021, 11, 107. [Google Scholar] [CrossRef] [PubMed]
- Preto, J.; Krimm, I. The intrinsically disordered N-terminus of the voltage-dependent anion channel. PLoS Comput. Biol. 2021, 17, e1008750. [Google Scholar] [CrossRef]
- Preto, J.; Gorny, H.; Krimm, I. A Deep Dive into VDAC1 Conformational Diversity Using All-Atom Simulations Provides New Insights into the Structural Origin of the Closed States. Int. J. Mol. Sci. 2022, 23, 1175. [Google Scholar] [CrossRef]
- De Pinto, V.; Guarino, F.; Guarnera, A.; Messina, A.; Reina, S.; Tomasello, F.M.; Palermo, V.; Mazzoni, C. Characterization of human VDAC isoforms: A peculiar function for VDAC3? Biochim. Biophys. Acta 2010, 1797, 1268–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reina, S.; Magrì, A.; Lolicato, M.; Guarino, F.; Impellizzeri, A.; Maier, E.; Benz, R.; Ceccarelli, M.; De Pinto, V.; Messina, A. Deletion of β-strands 9 and 10 converts VDAC1 voltage-dependence in an asymmetrical process. Biochim. Biophys. Acta 2013, 1827, 793–805. [Google Scholar] [CrossRef]
- Amodeo, G.F.; Scorciapino, M.A.; Messina, A.; De Pinto, V.; Ceccarelli, M. Charged residues distribution modulates selectivity of the open state of human isoforms of the voltage dependent anion-selective channel. PLoS ONE 2014, 9, e103879. [Google Scholar] [CrossRef] [Green Version]
- Rappaport, S.M.; Teijido, O.; Hoogerheide, D.P.; Rostovtseva, T.K.; Berezhkovskii, A.M.; Bezrukov, S.M. Conductance hysteresis in the voltage-dependent anion channel. Eur. Biophys. J. 2015, 44, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Queralt-Martín, M.; Bergdoll, L.; Jacobs, D.; Bezrukov, S.M.; Abramson, J.; Rostovtseva, T.K. Assessing the role of residue E73 and lipid headgroup charge in VDAC1 voltage gating. Biochim. Biophys. Acta Bioenerg. 2019, 1860, 22–29. [Google Scholar] [CrossRef]
- Rovini, A.; Gurnev, P.A.; Beilina, A.; Queralt-Martín, M.; Rosencrans, W.; Cookson, M.R.; Bezrukov, S.M.; Rostovtseva, T.K. Molecular mechanism of olesoxime-mediated neuroprotection through targeting α-synuclein interaction with mitochondrial VDAC. Cell. Mol. Life. Sci. 2020, 77, 3611–3626. [Google Scholar] [CrossRef]
- Rostovtseva, T.K.; Queralt-Martín, M.; Rosencrans, W.M.; Bezrukov, S.M. Targeting the Multiple Physiologic Roles of VDAC with Steroids and Hydrophobic Drugs. Front. Physiol. 2020, 11, 446. [Google Scholar] [CrossRef] [PubMed]
- Rostovtseva, T.K.; Gurnev, P.A.; Protchenko, O.; Hoogerheide, D.P.; Yap, T.L.; Philpott, C.C.; Lee, J.C.; Bezrukov, S.M. α-Synuclein Shows High Affinity Interaction with Voltage-dependent Anion Channel, Suggesting Mechanisms of Mitochondrial Regulation and Toxicity in Parkinson Disease. J. Biol. Chem. 2015, 290, 18467–18477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camara, A.K.S.; Zhou, Y.; Wen, P.C.; Tajkhorshid, E.; Kwok, W.M. Mitochondrial VDAC1: A Key Gatekeeper as Potential Therapeutic Target. Front. Physiol. 2017, 8, 460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heslop, K.A.; Milesi, V.; Maldonado, E.N. VDAC Modulation of Cancer Metabolism: Advances and Therapeutic Challenges. Front. Physiol. 2021, 12, 742839. [Google Scholar] [CrossRef]
- Rajendran, M.; Queralt-Martín, M.; Gurnev, P.A.; Rosencrans, W.M.; Rovini, A.; Jacobs, D.; Abrantes, K.; Hoogerheide, D.P.; Bezrukov, S.M.; Rostovtseva, T.K. Restricting α-synuclein transport into mitochondria by inhibition of α-synuclein-VDAC complexation as a potential therapeutic target for Parkinson’s disease treatment. Cell. Mol. Life Sci. 2022, 79, 368. [Google Scholar] [CrossRef] [PubMed]
- Rosencrans, W.M.; Queralt-Martin, M.; Lessen, H.J.; Larimi, M.G.; Rajendran, M.; Chou, T.F.; Mahalakshmi, R.; Sodt, A.J.; Yu, T.Y.; Bezrukov, S.M.; et al. Defining the roles and regulation of the mitochondrial VDAC isoforms one molecule at a time. Biophys. J. 2023, 122, 93a. [Google Scholar] [CrossRef]
- Bayrhuber, M.; Meins, T.; Habeck, M.; Becker, S.; Giller, K.; Villinger, S.; Vonrhein, C.; Griesinger, C.; Zweckstetter, M.; Zeth, K. Structure of the human voltage-dependent anion channel. Proc. Natl. Acad. Sci. USA 2008, 105, 15370–15375. [Google Scholar] [CrossRef]
- Ujwal, R.; Cascio, D.; Colletier, J.P.; Faham, S.; Zhang, J.; Toro, L.; Ping, P.; Abramson, J. The crystal structure of mouse VDAC1 at 2.3 A resolution reveals mechanistic insights into metabolite gating. Proc. Natl. Acad. Sci. USA 2008, 105, 17742–17747. [Google Scholar] [CrossRef]
- Hiller, S.; Wagner, G. The role of solution NMR in the structure determinations of VDAC-1 and other membrane proteins. Curr. Opin. Struct. Biol. 2009, 19, 396–401. [Google Scholar] [CrossRef] [Green Version]
- Rostovtseva, T.K.; Liu, T.T.; Colombini, M.; Parsegian, V.A.; Bezrukov, S.M. Positive cooperativity without domains or subunits in a monomeric membrane channel. Proc. Natl. Acad. Sci. USA 2000, 97, 7819–7822. [Google Scholar] [CrossRef]
- Colombini, M.; Mannella, C.A. VDAC, the early days. Biochim. Biophys. Acta 2012, 1818, 1438–1443. [Google Scholar] [CrossRef] [Green Version]
- Teijido, O.; Ujwal, R.; Hillerdal, C.O.; Kullman, L.; Rostovtseva, T.K.; Abramson, J. Affixing N-terminal α-helix to the wall of the voltage-dependent anion channel does not prevent its voltage gating. J. Biol. Chem. 2012, 287, 11437–11445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teijido, O.; Rappaport, S.M.; Chamberlin, A.; Noskov, S.Y.; Aguilella, V.M.; Rostovtseva, T.K.; Bezrukov, S.M. Acidification asymmetrically affects voltage-dependent anion channel implicating the involvement of salt bridges. J. Biol. Chem. 2014, 289, 23670–23682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queralt-Martín, M.; Hoogerheide, D.P.; Noskov, S.Y.; Berezhkovskii, A.M.; Rostovtseva, T.K.; Bezrukov, S.M. VDAC Gating Thermodynamics, but Not Gating Kinetics, Are Virtually Temperature Independent. Biophys. J. 2020, 119, 2584–2592. [Google Scholar] [CrossRef]
- Bezrukov, S.M.; Kasianowicz, J.J. Current noise reveals protonation kinetics and number of ionizable sites in an open protein ion channel. Phys. Rev. Lett. 1993, 70, 2352–2355. [Google Scholar] [CrossRef] [PubMed]
- Kasianowicz, J.J.; Bezrukov, S.M. Protonation dynamics of the alpha-toxin ion channel from spectral analysis of pH-dependent current fluctuations. Biophys. J. 1995, 69, 94–105. [Google Scholar] [CrossRef] [Green Version]
- Korchev, Y.E.; Bashford, C.L.; Alder, G.M.; Kasianowicz, J.J.; Pasternak, C.A. Low-conductance states of a single-ion channel are not closed. J. Membr. Biol. 1995, 147, 233–239. [Google Scholar] [CrossRef]
- Mohammad, M.M.; Iyer, R.; Howard, K.R.; McPike, M.P.; Borer, P.N.; Movileanu, L. Engineering a Rigid Protein Tunnel for Biomolecular Detection. J. Am. Chem. Soc. 2012, 134, 9521–9531. [Google Scholar] [CrossRef] [Green Version]
- Nestorovich, E.M.; Rostovtseva, T.K.; Bezrukov, S.M. Residue ionization and ion transport through OmpF channels. Biophys. J. 2003, 85, 3718–3729. [Google Scholar] [CrossRef] [Green Version]
- Queralt-Martín, M.; Peiró-González, C.; Aguilella-Arzo, M.; Alcaraz, A. Effects of extreme pH on ionic transport through protein nanopores: The role of ion diffusion and charge exclusion. Phys. Chem. Chem. Phys. 2016, 18, 21668–21675. [Google Scholar] [CrossRef]
- Alcaraz, A.; Queralt-Martín, M. On the different sources of cooperativity in pH titrating sites of a membrane protein channel. Eur. Phys. J. E Soft Matter. 2016, 39, 29. [Google Scholar] [CrossRef] [Green Version]
- Perez-Rathke, A.; Fahie, M.A.; Chisholm, C.; Liang, J.; Chen, M. Mechanism of OmpG pH-Dependent Gating from Loop Ensemble and Single Channel Studies. J. Am. Chem. Soc. 2018, 140, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Fahie, M.A.V.; Li, F.; Palmer, C.; Yoon, C.; Chen, M. Modifying the pH sensitivity of OmpG nanopore for improved detection at acidic pH. Biophys. J. 2022, 121, 731–741. [Google Scholar] [CrossRef]
- Clapham, D.E.; Miller, C. A thermodynamic framework for understanding temperature sensing by transient receptor potential (TRP) channels. Proc. Natl. Acad. Sci. USA 2011, 108, 19492–19497. [Google Scholar] [CrossRef]
- Mills, A.; Le, H.T.; Coulton, J.W.; Duong, F. FhuA interactions in a detergent-free nanodisc environment. Biochim. Biophys. Acta 2014, 1838, 364–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Susac, L.; Horst, R.; Wuthrich, K. Solution-NMR characterization of outer-membrane protein A from E. coli in lipid bilayer nanodiscs and detergent micelles. Chembiochem 2014, 15, 995–1000. [Google Scholar] [CrossRef] [Green Version]
- Chimerel, C.; Movileanu, L.; Pezeshki, S.; Winterhalter, M.; Kleinekathofer, U. Transport at the nanoscale: Temperature dependence of ion conductance. Eur. Biophys. J. 2008, 38, 121–125. [Google Scholar] [CrossRef]
- Pezeshki, S.; Chimerel, C.; Bessonov, A.N.; Winterhalter, M.; Kleinekathofer, U. Understanding ion conductance on a molecular level: An all-atom modeling of the bacterial porin OmpF. Biophys. J. 2009, 97, 1898–1906. [Google Scholar] [CrossRef] [Green Version]
- Biro, I.; Pezeshki, S.; Weingart, H.; Winterhalter, M.; Kleinekathofer, U. Comparing the temperature-dependent conductance of the two structurally similar E. coli porins OmpC and OmpF. Biophys. J. 2010, 98, 1830–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakharian, E.; Reusch, R.N. Outer membrane protein A of Escherichia coli forms temperature-sensitive channels in planar lipid bilayers. FEBS Lett. 2003, 555, 229–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheneke, B.R.; van den Berg, B.; Movileanu, L. Quasithermodynamic contributions to the fluctuations of a protein nanopore. ACS Chem. Biol. 2015, 10, 784–794. [Google Scholar] [CrossRef] [PubMed]
- Sackmann, B.; Neher, E. Single-Channel Recording, 2nd ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 1995. [Google Scholar]
- Jaikaran, D.C.J.; Woolley, G.A. Characterization of thermal-isomerization at the single-molecule level. J. Phys. Chem. 1995, 99, 13352–13355. [Google Scholar] [CrossRef]
- Andersen, O.S. Graphic representation of the results of kinetic analyses. J. Gen. Physiol. 1999, 114, 589–590. [Google Scholar] [PubMed]
- Howorka, S.; Movileanu, L.; Braha, O.; Bayley, H. Kinetics of duplex formation for individual DNA strands within a single protein nanopore. Proc. Natl. Acad. Sci. USA 2001, 98, 12996–13001. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.F.; Gu, L.Q.; Cheley, S.; Bayley, H. Single Protein Pores Containing Molecular Adapters at High Temperatures. Angew. Chem. Int. Ed. Engl. 2005, 44, 1495–1499. [Google Scholar] [CrossRef]
- Cheneke, B.R.; Indic, M.; van den Berg, B.; Movileanu, L. An Outer Membrane Protein undergoes Enthalpy- and Entropy-driven Transitions. Biochemistry 2012, 51, 5348–5358. [Google Scholar] [CrossRef] [Green Version]
- Movileanu, L.; Schiff, E.A. Entropy-enthalpy Compensation of Biomolecular Systems in Aqueous Phase: A Dry Perspective. Monatsh. Chem. 2013, 144, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Alcaraz, A.; Queralt-Martín, M.; Verdiá-Báguena, C.; Aguilella, V.M.; Mafé, S. Entropy-enthalpy compensation at the single protein level: pH sensing in the bacterial channel OmpF. Nanoscale 2014, 6, 15210–15215. [Google Scholar] [CrossRef]
- Jung, Y.; Bayley, H.; Movileanu, L. Temperature-responsive protein pores. J. Am. Chem. Soc. 2006, 128, 15332–15340. [Google Scholar] [CrossRef]
- Phillips, R.; Ursell, T.; Wiggins, P.; Sens, P. Emerging roles for lipids in shaping membrane-protein function. Nature 2009, 459, 379–385. [Google Scholar] [CrossRef] [Green Version]
- Eddy, M.T.; Ong, T.C.; Clark, L.; Teijido, O.; van der Wel, P.C.; Garces, R.; Wagner, G.; Rostovtseva, T.K.; Griffin, R.G. Lipid dynamics and protein-lipid interactions in 2D crystals formed with the beta-barrel integral membrane protein VDAC1. J. Am. Chem. Soc. 2012, 134, 6375–6387. [Google Scholar] [CrossRef] [Green Version]
- Goldfine, H. Bacterial membranes and lipid packing theory. J. Lipid Res. 1984, 25, 1501–1507. [Google Scholar] [CrossRef] [PubMed]
- Gruner, S.M. Intrinsic curvature hypothesis for biomembrane lipid composition: A role for nonbilayer lipids. Proc. Natl. Acad. Sci. USA 1985, 82, 3665–3669. [Google Scholar] [CrossRef]
- Cullis, P.R.; Hope, M.J.; Tilcock, C.P. Lipid polymorphism and the roles of lipids in membranes. Chem. Phys. Lipids 1986, 40, 127–144. [Google Scholar] [CrossRef]
- Frolov, V.A.; Shnyrova, A.V.; Zimmerberg, J. Lipid polymorphisms and membrane shape. Cold Spring Harb. Perspect. Biol. 2011, 3, a004747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laganowsky, A.; Reading, E.; Allison, T.M.; Ulmschneider, M.B.; Degiacomi, M.T.; Baldwin, A.J.; Robinson, C.V. Membrane proteins bind lipids selectively to modulate their structure and function. Nature 2014, 510, 172–175. [Google Scholar] [CrossRef] [Green Version]
- Liko, I.; Degiacomi, M.T.; Lee, S.; Newport, T.D.; Gault, J.; Reading, E.; Hopper, J.T.S.; Housden, N.G.; White, P.; Colledge, M.; et al. Lipid binding attenuates channel closure of the outer membrane protein OmpF. Proc. Natl. Acad. Sci. USA 2018, 115, 6691–6696. [Google Scholar] [CrossRef] [Green Version]
- Perini, D.A.; Alcaraz, A.; Queralt-Martín, M. Lipid Headgroup Charge and Acyl Chain Composition Modulate Closure of Bacterial β-Barrel Channels. Int. J. Mol. Sci. 2019, 20, 674. [Google Scholar] [CrossRef] [Green Version]
- Hwang, W.L.; Chen, M.; Cronin, B.; Holden, M.A.; Bayley, H. Asymmetric droplet interface bilayers. J. Am. Chem. Soc. 2008, 130, 5878–5879. [Google Scholar] [CrossRef] [PubMed]
- Howorka, S. Building membrane nanopores. Nat. Nanotechnol. 2017, 12, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Cressiot, B.; Bacri, L.; Pelta, J. The Promise of Nanopore Technology: Advances in the Discrimination of Protein Sequences and Chemical Modifications. Small Methods 2020, 4, 2000090. [Google Scholar] [CrossRef]
- Tanimoto, I.M.F.; Cressiot, B.; Greive, S.J.; Le Pioufle, B.; Bacri, L.; Pelta, J. Focus on using nanopore technology for societal health, environmental, and energy challenges. Nano Res. 2022, 15, 9906–9920. [Google Scholar] [CrossRef] [PubMed]
- Howorka, S.; Siwy, Z. Nanopores and Nanochannels: From Gene Sequencing to Genome Mapping. ACS Nano 2016, 10, 9768–9771. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, Z.; Haque, F.; Guo, P. Engineering of protein nanopores for sequencing, chemical or protein sensing and disease diagnosis. Curr. Opin. Biotechnol. 2018, 51, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.W.F.; Reiner, J.E. The Utility of Nanopore Technology for Protein and Peptide Sensing. Proteomics 2018, 18, e18000262018. [Google Scholar] [CrossRef] [PubMed]
- Schmid, S.; Dekker, C. Nanopores: A versatile tool to study protein dynamics. Essays Biochem. 2021, 65, 93–107. [Google Scholar] [CrossRef]
- Nova, I.C.; Ritmejeris, J.; Brinkerhoff, H.; Koenig, T.J.R.; Gundlach, J.H.; Dekker, C. Detection of phosphorylation post-translational modifications along single peptides with nanopores. Nat. Biotechnol. 2023. [Google Scholar] [CrossRef]
- Restrepo-Perez, L.; Joo, C.; Dekker, C. Paving the way to single-molecule protein sequencing. Nat. Nanotechnol. 2018, 13, 786–796. [Google Scholar] [CrossRef] [Green Version]
- Brinkerhoff, H.; Kang, A.S.W.; Liu, J.; Aksimentiev, A.; Dekker, C. Multiple rereads of single proteins at single-amino acid resolution using nanopores. Science 2021, 374, 1509–1513. [Google Scholar] [CrossRef]
- Laszlo, A.H.; Derrington, I.M.; Gundlach, J.H. MspA nanopore as a single-molecule tool: From sequencing to SPRNT. Methods 2016, 105, 75–89. [Google Scholar] [CrossRef]
- Cherf, G.M.; Lieberman, K.R.; Rashid, H.; Lam, C.E.; Karplus, K.; Akeson, M. Automated forward and reverse ratcheting of DNA in a nanopore at 5-A precision. Nat. Biotechnol. 2012, 30, 344–348. [Google Scholar] [CrossRef] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayse, L.A.; Movileanu, L. Gating of β-Barrel Protein Pores, Porins, and Channels: An Old Problem with New Facets. Int. J. Mol. Sci. 2023, 24, 12095. https://doi.org/10.3390/ijms241512095
Mayse LA, Movileanu L. Gating of β-Barrel Protein Pores, Porins, and Channels: An Old Problem with New Facets. International Journal of Molecular Sciences. 2023; 24(15):12095. https://doi.org/10.3390/ijms241512095
Chicago/Turabian StyleMayse, Lauren A., and Liviu Movileanu. 2023. "Gating of β-Barrel Protein Pores, Porins, and Channels: An Old Problem with New Facets" International Journal of Molecular Sciences 24, no. 15: 12095. https://doi.org/10.3390/ijms241512095