

Pharmacological Screening of Kv7.1 and Kv7.1/KCNE1 Activators as Potential Antiarrhythmic Drugs in the Zebrafish Heart
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
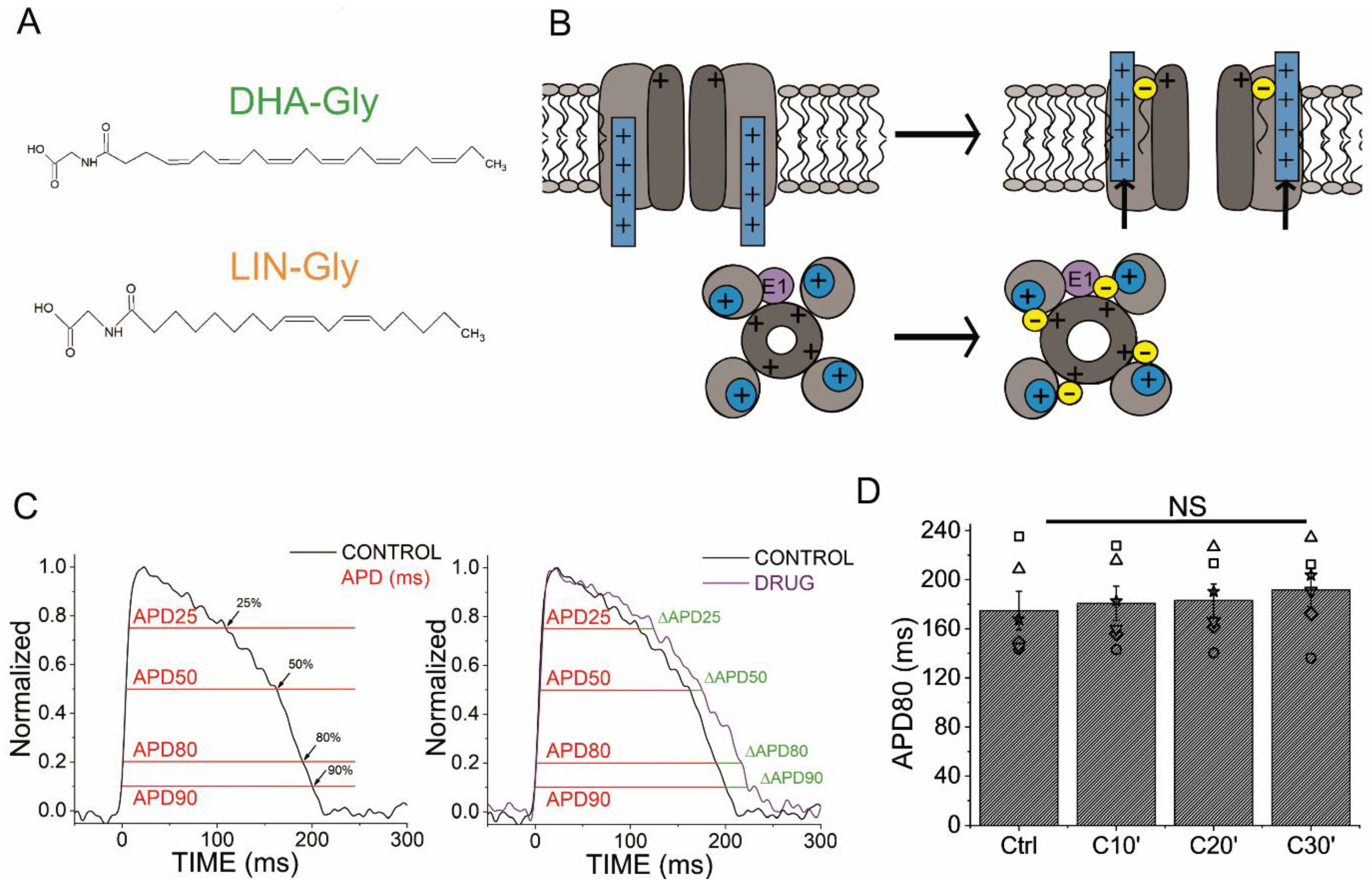
2.1. AP Optical Recordings Are Stable in Control Conditions
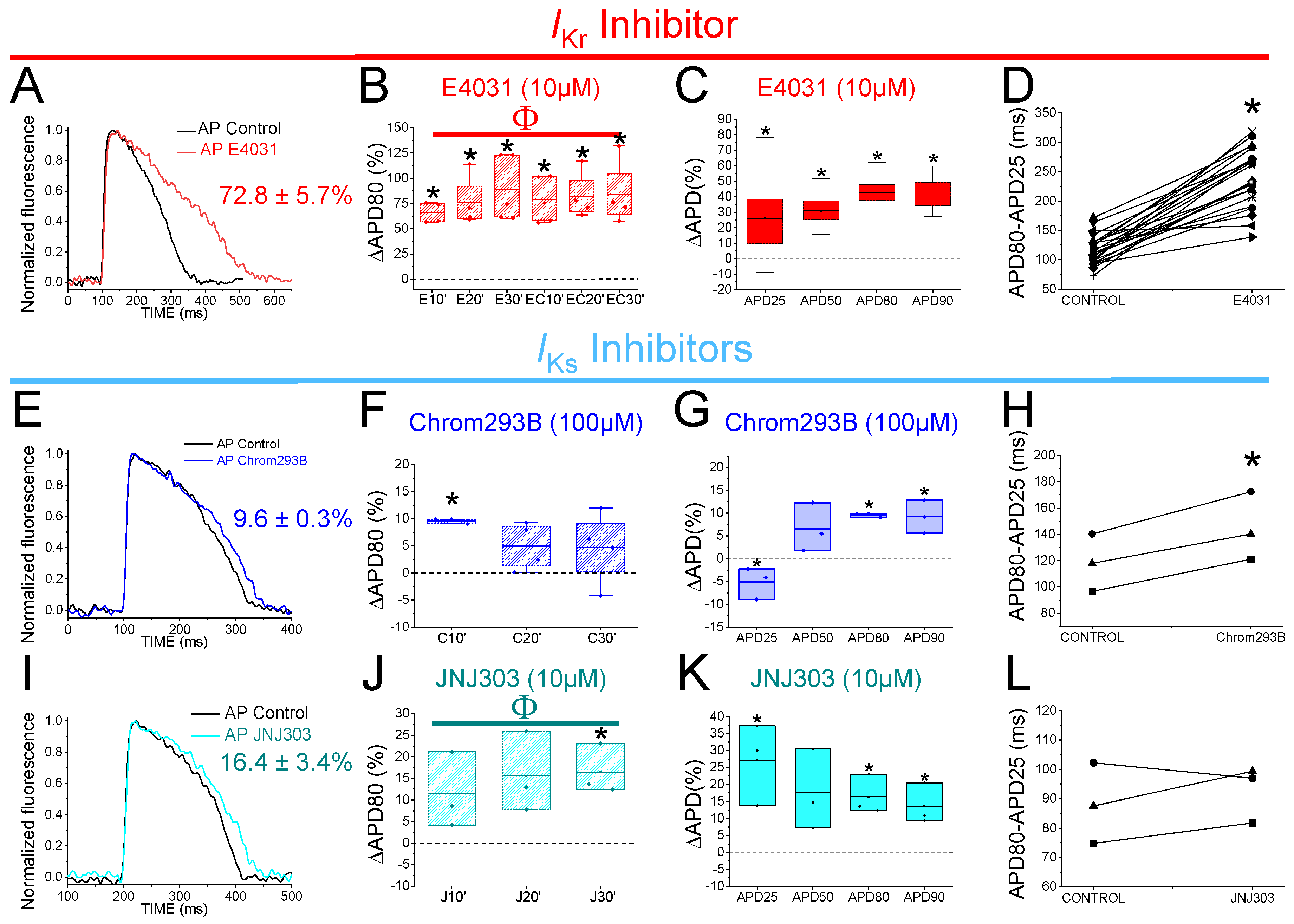
2.2. The IKr Inhibitor E4031 Prolongs Zebrafish APD
2.3. Two Different Human IKs Inhibitors Have Different Effects on Zebrafish AP
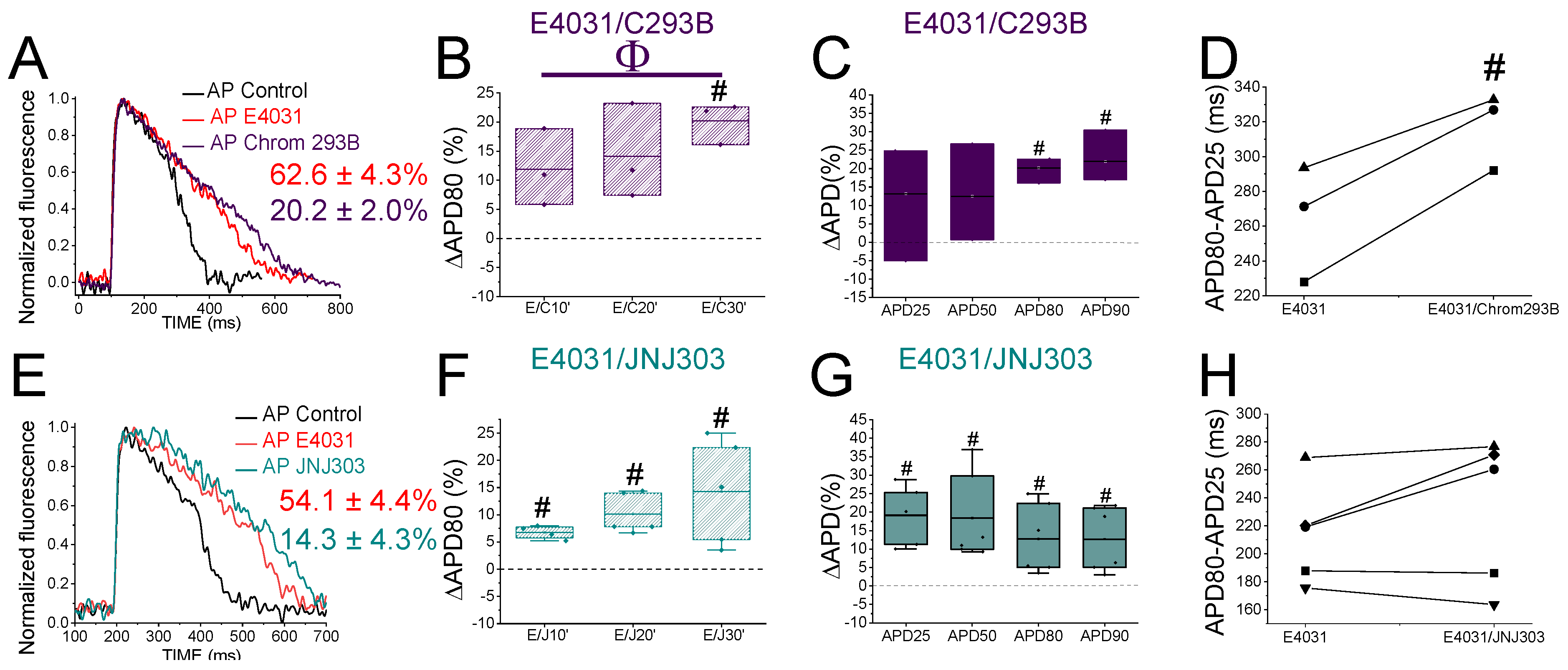
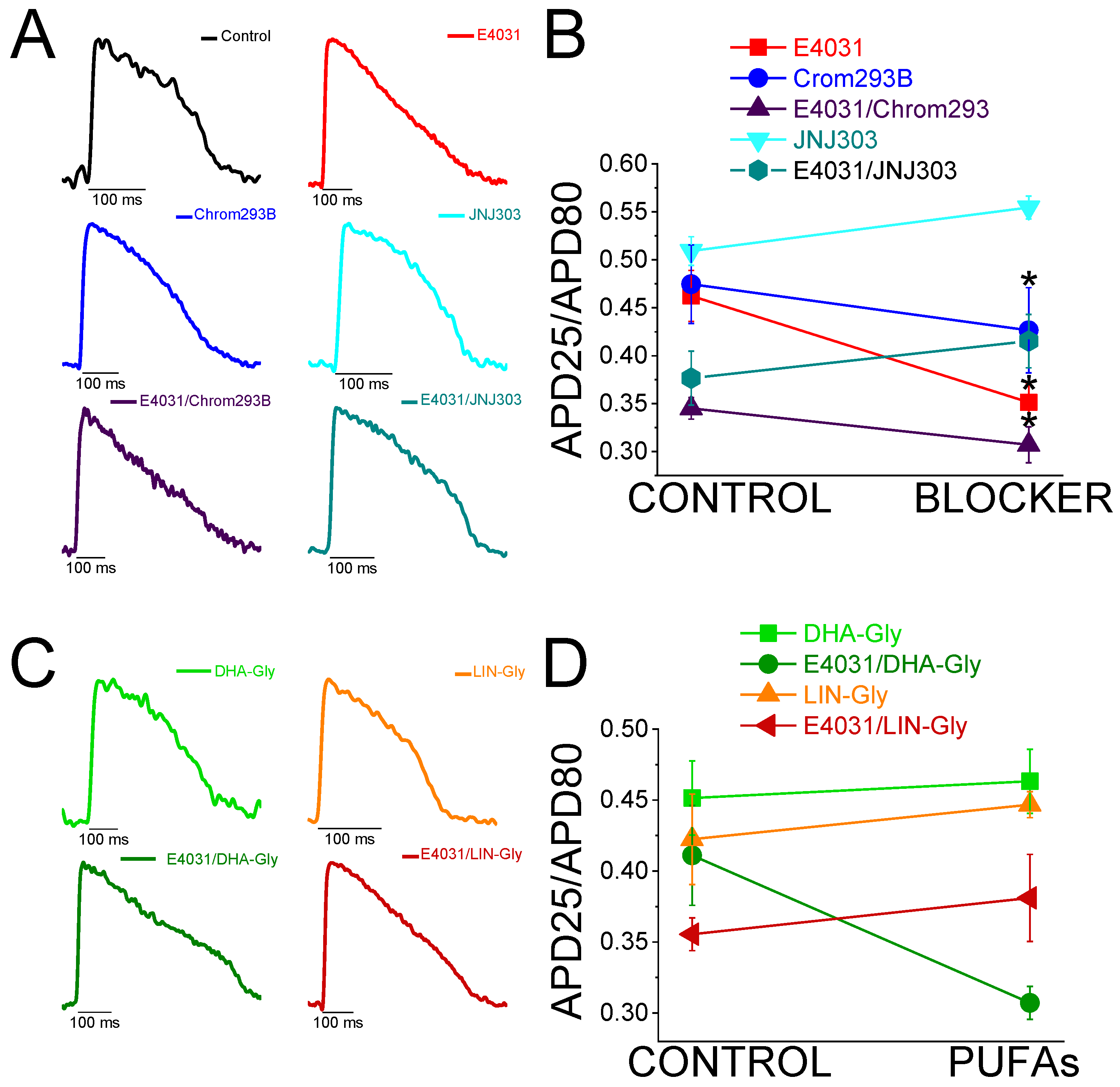
2.4. The IKs Blocker Chromanol 293B Prolongs the APD More after IKr Inhibition
2.5. The Human IKs Inhibitor JNJ303 Does Not Prolong the APD More after IKr Inhibition
2.6. JNJ303 Is Not an Effective IKs Inhibitor of Zebrafish Kv7.1/KCNE1 Currents
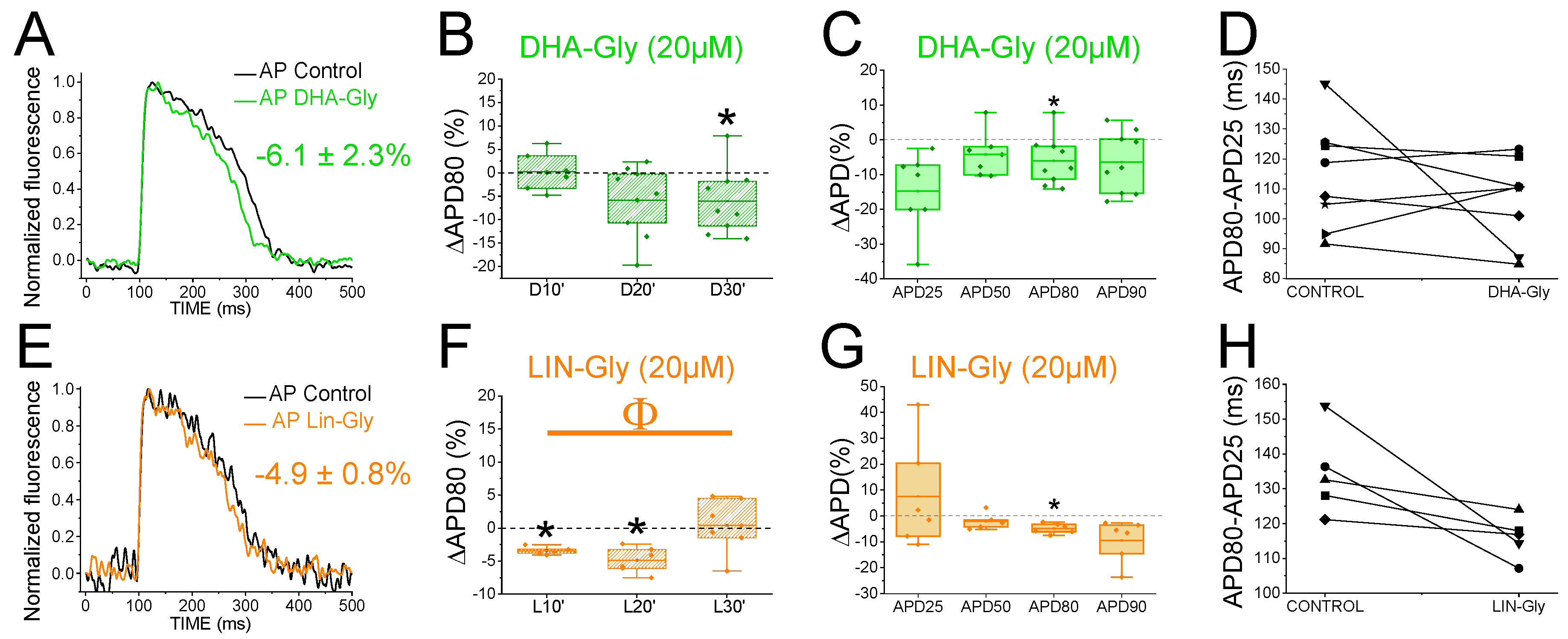
2.7. PUFA Analogues Have Modest Effects on Zebrafish AP
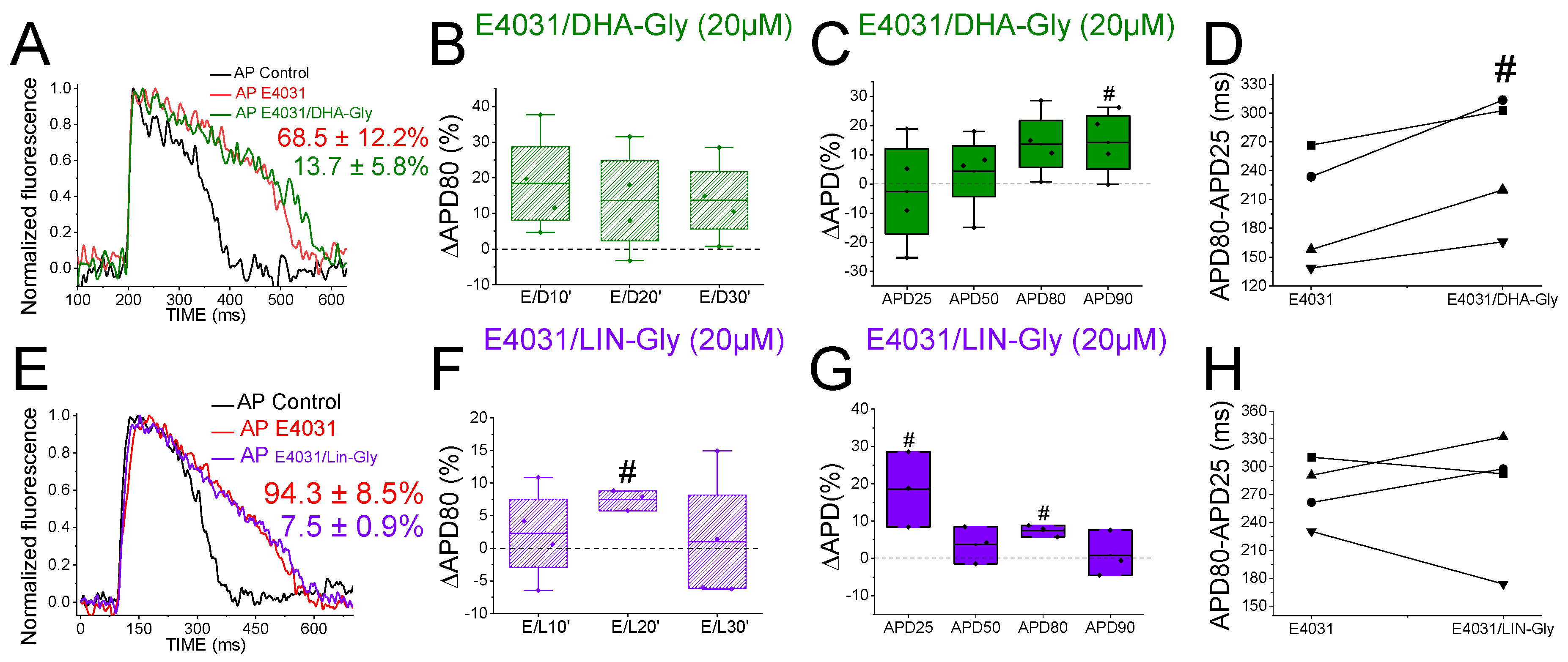
2.8. PUFA Analogues Do Not Reverse APD in Drug-Induced LQTS Zebrafish Hearts
2.9. DHA-Gly and LIN-Gly Have Only Modest Effects on Zebrafish Kv7.1/KCNE1 and Kv7.1 Channels
2.10. DHA-Gly and LIN-Gly Did Not Reverse the Triangulation Effect of Drug-Induced LQT2
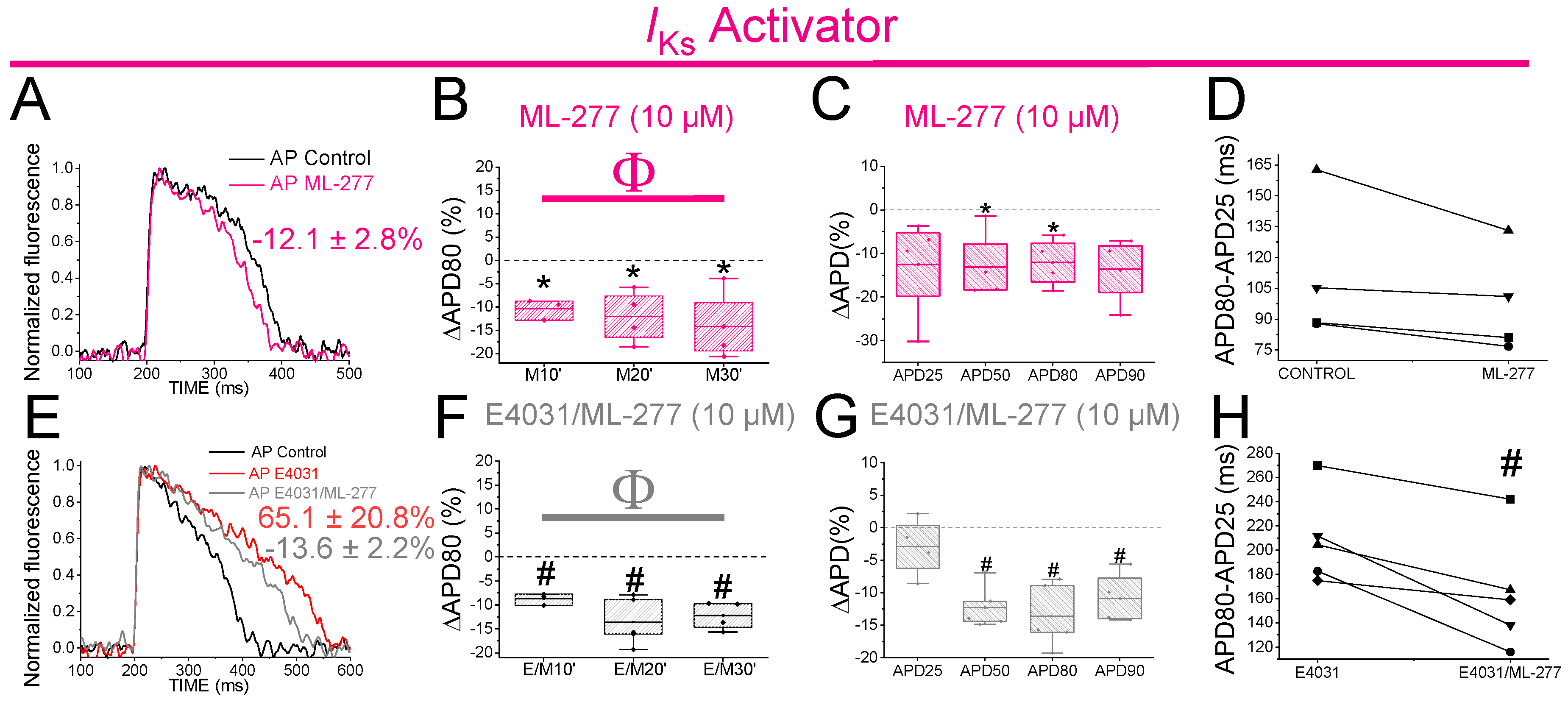
2.11. The IKs Activator ML-277 Partially Restored the APD in Drug-Induced LQT2 Zebrafish Hearts
3. Discussion
3.1. IKr and IKs Contribution to Zebrafish AP Repolarization
3.2. Small Effects of Other Human IKs Inhibitors
3.3. PUFA Analogues Slightly Shorten Zebrafish AP
4. Materials and Methods
4.1. Zebrafish Heart Isolation Procedure
4.2. Action Potential Optical Recordings and Analysis
4.3. Molecular Biology
4.4. Two-Electrode Voltage Clamp
4.5. Statistics
5. Study Limitations, Future Directions, and Clinical Relevance
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Levine, E.; Rosero, S.Z.; Budzikowski, A.S.; Moss, A.J.; Zareba, W.; Daubert, J. Congenital long QT syndrome: Considerations for primary care physicians. Cleve. Clin. J. Med. 2008, 75, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Bohannon, B.M.; de la Cruz, A.; Wu, X.; Jowais, J.J.; Perez, M.E.; Dykxhoorn, D.M.; Liin, S.I.; Larsson, H.P. Polyunsaturated fatty acid analogues differentially affect cardiac NaV, CaV, and KV channels through unique mechanisms. Elife 2020, 9, e51453. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, P.J.; Stramba-Badiale, M.; Crotti, L.; Pedrazzini, M.; Besana, A.; Bosi, G.; Gabbarini, F.; Goulene, K.; Insolia, R.; Mannarino, S.; et al. Prevalence of the Congenital Long-QT Syndrome. Circulation 2009, 120, 1761–1767. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.E.; Venkateshappa, R.; Pang, Z.K.; Faizi, S.; Tibbits, G.F.; Claydon, T.W. Utility of Zebrafish Models of Acquired and Inherited Long QT Syndrome. Front. Physiol. 2020, 11, 624129. [Google Scholar] [CrossRef]
- Chi, N.C.; Shaw, R.M.; Jungblut, B.; Huisken, J.; Ferrer, T.; Arnaout, R.; Scott, I.; Beis, D.; Xiao, T.; Baier, H.; et al. Genetic and physiologic dissection of the vertebrate cardiac conduction system. PLoS Biol. 2008, 6, e109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemtsas, P.; Wettwer, E.; Christ, T.; Weidinger, G.; Ravens, U. Adult zebrafish heart as a model for human heart? An electrophysiological study. J. Mol. Cell. Cardiol. 2010, 48, 161–171. [Google Scholar] [CrossRef]
- Milan, D.J.; Jones, I.L.; Ellinor, P.T.; MacRae, C.A.; Gut, P.; Reischauer, S.; Stainier, D.Y.R.; Arnaout, R.; Lin, E.; Craig, C.; et al. In vivo recording of adult zebrafish electrocardiogram and assessment of drug-induced QT prolongation. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H269–H273. [Google Scholar] [CrossRef]
- Zhao, Y.; James, N.; Beshay, A.R.; Chang, E.E.; Lin, A.; Bashar, F.; Wassily, A.; Nguyen, B.; Nguyen, T.P. Adult zebrafish ventricular electrical gradients as tissue mechanisms of ECG patterns under baseline vs. oxidative stress. Cardiovasc. Res. 2020, 117, 1891–1907. [Google Scholar] [CrossRef]
- Vornanen, M.; Hassinen, M. Zebrafish heart as a model for human cardiac electrophysiology. Channels 2016, 10, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Tristani-Firouzi, M.; Chen, J.; Mitcheson, J.S.; Sanguinetti, M.C. Molecular biology of K(+) channels and their role in cardiac arrhythmias. Am. J. Med. 2001, 110, 50–59. [Google Scholar] [CrossRef]
- Trudeau, M.C.; Warmke, J.W.; Ganetzky, B.; Robertson, G.A. HERG, a human inward rectifier in the voltage-gated potassium channel family. Science 1995, 269, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Banyasz, T.; Jian, Z.; Horvath, B.; Khabbaz, S.; Izu, L.T.; Chen-Izu, Y. Beta-adrenergic stimulation reverses the I Kr-I Ks dominant pattern during cardiac action potential. Pflug. Arch. 2014, 466, 2067–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Gao, X.; Wang, Y.; Peng, L.; Zhu, Y.; Wang, S. IKs protects from ventricular arrhythmia during cardiac ischemia and reperfusion in rabbits by preserving the repolarization reserve. PLoS ONE 2012, 7, e31545. [Google Scholar] [CrossRef] [PubMed]
- Giudicessi, J.R.; Wilde, A.A.M.; Ackerman, M.J. The genetic architecture of long QT syndrome: A critical reappraisal. Trends Cardiovasc. Med. 2018, 28, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Ravens, U. Ionic basis of cardiac electrophysiology in zebrafish compared to human hearts. Prog. Biophys. Mol. Biol. 2018, 138, 38–44. [Google Scholar] [CrossRef]
- Alday, A.; Alonso, H.; Gallego, M.; Urrutia, J.; Letamendia, A.; Callol, C.; Casis, O. Ionic channels underlying the ventricular action potential in zebrafish embryo. Pharmacol. Res. 2014, 84, 26–31. [Google Scholar] [CrossRef]
- Wu, C.; Sharma, K.; Laster, K.; Hersi, M.; Torres, C.; Lukas, T.J.; Moore, E.J. Kcnq1-5 (Kv7.1-5) potassium channel expression in the adult zebrafish. BMC Physiol. 2014, 14, 1. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.T.; Wu, C.-K.; Chiang, F.-T.; Tseng, C.-D.; Lee, J.-K.; Yu, C.-C.; Wang, Y.-C.; Lai, L.-P.; Lin, J.-L.; Hwang, J.-J. In-vitro recording of adult zebrafish heart electrocardiogram—A platform for pharmacological testing. Clin. Chim. Acta 2011, 412, 1963–1967. [Google Scholar] [CrossRef]
- Abramochkin, D.V.; Hassinen, M.; Vornanen, M. Transcripts of Kv7.1 and MinK channels and slow delayed rectifier K(+) current (I(Ks)) are expressed in zebrafish (Danio rerio) heart. Pflug. Arch. 2018, 470, 1753–1764. [Google Scholar] [CrossRef]
- Moss, A.J.; Windle, J.R.; Hall, W.J.; Zareba, W.; Robinson, J.L.; McNitt, S.; Severski, P.; Rosero, S.; Daubert, J.P.; Qi, M.; et al. Safety and efficacy of flecainide in subjects with Long QT-3 syndrome (DeltaKPQ mutation): A randomized, double-blind, placebo-controlled clinical trial. Ann. Noninvasive Electrocardiol. 2005, 10 (Suppl. S4), 59–66. [Google Scholar] [CrossRef]
- Groh, W.J.; Silka, M.J.; Oliver, R.P.; Halperin, B.D.; McAnulty, J.H.; Kron, J. Use of implantable cardioverter-defibrillators in the congenital long QT syndrome. Am. J. Cardiol. 1996, 78, 703–706. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, Q.-F.; Du, R.; Xu, Q.-M.; Ke, Q.-M.; Wang, B.; Chen, X.-L.; Tian, L.; Zhang, S.-Y.; Kang, C.-L.; et al. Congenital long QT syndrome caused by the F275S KCNQ1 mutation: Mechanism of impaired channel function. Biochem. Biophys. Res. Commun. 2009, 380, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Moss, A.J.; McDonald, J. Unilateral cervicothoracic sympathetic ganglionectomy for the treatment of long QT interval syndrome. N. Engl. J. Med. 1971, 285, 903–904. [Google Scholar] [CrossRef] [PubMed]
- Sgro, A.; Drake, T.M.; Lopez-Ayala, P.; Phan, K. Left cardiac sympathetic denervation in the management of long QT syndrome and catecholaminergic polymorphic ventricular tachycardia: A meta-regression. Congenit. Heart Dis. 2019, 14, 1102–1112. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.J., Jr.; Houle, M.S.; Stump, G.L.; Wallace, A.A.; Gilberto, D.B.; Jahansouz, H.; Smith, G.R.; Tebben, A.J.; Liverton, N.J.; Selnick, H.G.; et al. Antiarrhythmic efficacy of selective blockade of the cardiac slowly activating delayed rectifier current, I(Ks), in canine models of malignant ischemic ventricular arrhythmia. Circulation 1999, 100, 1917–1922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, P.J.; Crotti, L.; Insolia, R. Long-QT syndrome: From genetics to management. Circ. Arrhythm. Electrophysiol. 2012, 5, 868–877. [Google Scholar] [CrossRef] [Green Version]
- Varshneya, M.; Devenyi, R.A.; Sobie, E.A. Slow Delayed Rectifier Current Protects Ventricular Myocytes from Arrhythmic Dynamics Across Multiple Species: A Computational Study. Circ. Arrhythm. Electrophysiol. 2018, 11, e006558. [Google Scholar] [CrossRef]
- Wu, X.; Larsson, H. Insights into Cardiac IKs (KCNQ1/KCNE1) Channels Regulation. Int. J. Mol. Sci. 2020, 21, 9440. [Google Scholar] [CrossRef]
- Benatti, P.; Peluso, G.; Nicolai, R.; Calvani, M. Polyunsaturated fatty acids: Biochemical, nutritional and epigenetic properties. J. Am. Coll. Nutr. 2004, 23, 281–302. [Google Scholar] [CrossRef]
- Borjesson, S.I.; Hammarstrom, S.; Elinder, F. Lipoelectric modification of ion channel voltage gating by polyunsaturated fatty acids. Biophys. J. 2008, 95, 2242–2253. [Google Scholar] [CrossRef] [Green Version]
- Liin, S.I.; Yazdi, S.; Ramentol, R.; Barro-Soria, R.; Larsson, H.P. Mechanisms Underlying the Dual Effect of Polyunsaturated Fatty Acid Analogs on Kv7.1. Cell Rep. 2018, 24, 2908–2918. [Google Scholar] [CrossRef] [Green Version]
- Skarsfeldt, M.A.; Liin, S.I.; Larsson, H.P.; Bentzen, B.H. Polyunsaturated fatty acid-derived IKs channel activators shorten the QT interval ex-vivo and in-vivo. Acta Physiol. 2020, 229, e13471. [Google Scholar] [CrossRef] [PubMed]
- Lerche, C.; Bruhova, I.; Lerche, H.; Steinmeyer, K.; Wei, A.D.; Strutz-Seebohm, N.; Lang, F.; Busch, A.E.; Zhorov, B.S.; Seebohm, G. Chromanol 293B binding in KCNQ1 (Kv7.1) channels involves electrostatic interactions with a potassium ion in the selectivity filter. Mol. Pharmacol. 2007, 71, 1503–1511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrobel, E.; Rothenberg, I.; Krisp, C.; Hundt, F.; Fraenzel, B.; Eckey, K.; Linders, J.T.M.; Gallacher, D.J.; Towart, R.; Pott, L.; et al. KCNE1 induces fenestration in the Kv7.1/KCNE1 channel complex that allows for highly specific pharmacological targeting. Nat. Commun. 2016, 7, 12795. [Google Scholar] [CrossRef] [PubMed]
- Kannankeril, P.; Roden, D.M.; Darbar, D. Drug-induced long QT syndrome. Pharmacol. Rev. 2010, 62, 760–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hara, T.; Rudy, Y. Quantitative comparison of cardiac ventricular myocyte electrophysiology and response to drugs in human and nonhuman species. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H1023–H1030. [Google Scholar] [CrossRef] [Green Version]
- Jost, N.; Virág, L.; Bitay, M.; Takács, J.; Lengyel, C.; Biliczki, P.; Nagy, Z.; Bogáts, G.; Lathrop, D.A.; Papp, J.G.; et al. Restricting excessive cardiac action potential and QT prolongation: A vital role for IKs in human ventricular muscle. Circulation 2005, 112, 1392–1399. [Google Scholar] [CrossRef] [Green Version]
- Sanguinetti, M.C.; Jurkiewicz, N.K.; Scott, A.; Siegl, P.K. Isoproterenol antagonizes prolongation of refractory period by the class III antiarrhythmic agent E-4031 in guinea pig myocytes. Mechanism of action. Circ. Res. 1991, 68, 77–84. [Google Scholar] [CrossRef]
- Wang, Z.; Fermini, B.; Nattel, S. Rapid and slow components of delayed rectifier current in human atrial myocytes. Cardiovasc. Res. 1994, 28, 1540–1546. [Google Scholar] [CrossRef]
- Biliczki, P.; Virág, L.; Iost, N.; Papp, J.G.; Varró, A. Interaction of different potassium channels in cardiac repolarization in dog ventricular preparations: Role of repolarization reserve. Br. J. Pharmacol. 2002, 137, 361–368. [Google Scholar] [CrossRef] [Green Version]
- Haverinen, J.; Hassinen, M.; Dash, S.N.; Vornanen, M. Expression of calcium channel transcripts in the zebrafish heart: Dominance of T-type channels. J. Exp. Biol. 2018, 221 Pt 10, jeb179226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liin, S.I.; Ejneby, M.S.; Barro-Soria, R.; Skarsfeldt, M.A.; Larsson, J.E.; Starck Härlin, F.; Parkkari, T.; Bentzen, B.H.; Schmitt, N.; Larsson, H.P.; et al. Polyunsaturated fatty acid analogs act antiarrhythmically on the cardiac IKs channel. Proc. Natl. Acad. Sci. USA 2015, 112, 5714–5719. [Google Scholar] [CrossRef] [PubMed]
- Bohannon, B.M.; Jowais, J.J.; Nyberg, L.; Olivier-Meo, V.; Corradi, V.; Tieleman, D.P.; Liin, S.I.; Larsson, H.P. Mechanistic insights into robust cardiac I(Ks) potassium channel activation by aromatic polyunsaturated fatty acid analogues. Elife 2023, 12, e85773. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| APD80 (ms) | APD80 (ms) | (n, p) | ||
|---|---|---|---|---|
| CTRL | 174.9 ± 15.7 | Long CTRL | 185.2 ± 13.5 | n = 6, NS, p > 0.05 |
| E4031 | 332.2 ± 19.2 | Long E4031 | 338.6 ± 17.8 | n = 3, NS, p > 0.05 |
| E4031 CTRL | 217.6 ± 7.1 | E4031 | 350.5 ± 11.4 | n = 22, p < 0.001 |
| C293B CTRL | 225.9 ± 33.5 | C293B | 243.0 ± 39.8 | n = 3, NS, p > 0.05 |
| E4031 | 392.8 ± 14.0 | E4031/C293B | 463.5 ± 16.2 | n = 3, p < 0.05 |
| JNJ303 CTRL | 180.2 ± 17.3 | JNJ303 | 204.7 ± 13.5 | n = 3, p < 0.05 |
| E4031 | 325.6 ± 14.3 | E4031/JNJ303 | 369.8 ± 28.7 | n = 4, NS, p > 0.05 |
| DHA-Gly CTRL | 212.3 ± 9.1 | DHA-Gly | 206.1 ± 6.7 | n = 7, NS, p > 0.05 |
| LIN-Gly CTRL | 210.8 ± 14.9 | LIN-Gly | 204.8 ± 15.1 | n = 5, p < 0.05 |
| E4031 | 325.3 ± 36.3 | E4031/DHA-Gly | 438.6 ± 54.7 | n = 4, NS, p > 0.05 |
| E4031 | 397.9 ± 14.6 | E4031/LIN-Gly | 433.0 ± 19.7 | n = 3, NS, p > 0.05 |
| ML-277 CTRL | 236.4 ± 26.6 | ML-277 CTRL | 211.1 ± 22.3 | n = 3, p < 0.05 |
| E4031 | 301.5 ± 13.1 | E4031/ML-277 | 272.2 ± 8.4 | n = 3, p < 0.05 |
| ΔAPD25 (%) | (n, p) | ΔAPD50 (%) | (n, p) | ΔAPD80 (%) | (n, p) | ΔAPD90 (%) | (n, p) | |
|---|---|---|---|---|---|---|---|---|
| E4031 | 48.3 ± 14.6 | n = 26, p < 0.001 | 49.6 ± 4.8 | n = 26, p < 0.001 | 72.8 ± 5.7 | n = 26, p < 0.001 | 71.6 ± 5.8 | n = 26, p < 0.001 |
| C293B | −5.1 ± 2.0 | n = 3, p < 0.05 | 6.5 ± 3.1 | n = 3, NS, p > 0.05 | 9.6 ± 0.3 | n = 3, p < 0.05 | 9.2 ± 2.1 | n = 3, p < 0.01 |
| JNJ303 | 27.1 ± 7.0 | n = 3, p < 0.05 | 17.5 ± 6.8 | n = 3, NS, p > 0.05 | 16.4 ± 3.4 | n = 3, p < 0.01 | 13.6 ± 3.5 | n = 3, p < 0.05 |
| E4031/C293B | 13.2 ± 9.2 | n = 3, NS, p > 0.05 | 12.5 ± 7.6 | n = 3, NS, p > 0.05 | 20.2 ± 2.0 | n = 3, p < 0.01 | 22.0 ± 4.3 | n = 3, p < 0.05 |
| E4031/JNJ303 | 27.5 ± 10.5 | n = 5, p < 0.05 | 20.2 ± 5.5 | n = 5, p < 0.05 | 14.3 ± 4.3 | n = 5, p < 0.05 | 14.0 ± 4.1 | n = 5, p < 0.05 |
| ML-277 | −12.5 ± 6.0 | n = 4, NS, p > 0.05 | −13.1 ± 4.0 | n = 4, p < 0.05 | 12.1 ± 2.3 | n = 4, p < 0.05 | −13.6 ± 3.8 | n = 3, NS, p > 0.05 |
| E4031/ML-277 | 3.9 ± 7.1 | n = 5, NS, p > 0.05 | −12.3 ± 1.5 | n = 5, p < 0.01 | −13.6 ± 2.2 | n = 5, p < 0.01 | −14.0 ± 3.5 | n = 5, p < 0.05 |
| ΔAPD25 (%) | (n, p) | ΔAPD50 (%) | (n, p) | ΔAPD80 (%) | (n, p) | ΔAPD90 (%) | (n, p) | |
|---|---|---|---|---|---|---|---|---|
| DHA-Gly | −0.7 ± 10.5 | n = 9, NS, p > 0.05 | −4.5 ± 3.4 | n = 9, NS, p > 0.05 | −6.1 ± 2.3 | n = 9, p < 0.05 | −6.4 ± 3.0 | n = 9, NS, p > 0.05 |
| LIN-Gly | 7.6 ± 8.4 | n = 6, NS, p > 0.05 | −6.4 ± 4.5 | n = 6, NS, p > 0.05 | −4.9 ± 0.8 | n = 6, p < 0.01 | −9.4 ± 3.3 | n = 6, NS, p > 0.05 |
| E4031/DHA-Gly | −2.6 ± 9.5 | n = 4, NS, p > 0.05 | 4.4 ± 6.9 | n = 4, NS, p > 0.05 | 13.7 ± 5.8 | n = 4, NS, p > 0.05 | 14.2 ± 5.8 | n = 4, p < 0.05 |
| E4031/LIN-Gly | 18.6 ± 5.8 | n = 3, p < 0.05 | 3.7 ± 2.9 | n = 3, NS, p > 0.05 | 7.5 ± 0.9 | n = 3, p < 0.05 | 0.8 ± 3.6 | n = 3, NS, p > 0.05 |
| APD25/APD80 | APD25/APD80 | (n, p) | ||
|---|---|---|---|---|
| E4031 CTRL | 0.48 ± 0.03 | E4031 | 0.36 ± 0.02 | n = 24, p < 0.001 |
| C293B CTRL | 0.47 ± 0.14 | C293B | 0.41 ± 0.06 | n = 3, p < 0.01 |
| E4031 | 0.35 ± 0.01 | E4031/C293B | 0.31 ± 0.02 | n = 3, p < 0.05 |
| JNJ303 CTRL | 0.51 ± 0.02 | JNJ303 | 0.55 ± 0.01 | n = 3, NS, p > 0.05 |
| E4031 | 0.38 ± 0.03 | E4031/JNJ303 | 0.42 ± 0.03 | n = 5, NS, p > 0.05 |
| DHA-Gly CTRL | 0.45 ± 0.02 | DHA-Gly | 0.46 ± 0.02 | n = 9, NS, p > 0.05 |
| LIN-Gly CTRL | 0.41 ± 0.04 | LIN-Gly | 0.46 ± 0.01 | n = 5, NS, p > 0.05 |
| E4031 | 0.38 ± 0.11 | E4031/DHA-Gly | 0.33 ± 0.1 | n = 3, NS, p > 0.05 |
| E4031 | 0.34 ± 0.02 | E4031/LIN-Gly | 0.36 ± 0.04 | n = 5, NS, p > 0.05 |
| ML-277 CTRL | 0.51 ± 0.03 | ML-277 CTRL | 0.51 ± 0.03 | n = 4, NS, p > 0.05 |
| E4031 | 0.37 ± 0.03 | E4031/ML-277 | 0.44 ± 0.03 | n = 5, NS, p > 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De la Cruz, A.; Wu, X.; Rainer, Q.C.; Hiniesto-Iñigo, I.; Perez, M.E.; Edler, I.; Liin, S.I.; Larsson, H.P. Pharmacological Screening of Kv7.1 and Kv7.1/KCNE1 Activators as Potential Antiarrhythmic Drugs in the Zebrafish Heart. Int. J. Mol. Sci. 2023, 24, 12092. https://doi.org/10.3390/ijms241512092
De la Cruz A, Wu X, Rainer QC, Hiniesto-Iñigo I, Perez ME, Edler I, Liin SI, Larsson HP. Pharmacological Screening of Kv7.1 and Kv7.1/KCNE1 Activators as Potential Antiarrhythmic Drugs in the Zebrafish Heart. International Journal of Molecular Sciences. 2023; 24(15):12092. https://doi.org/10.3390/ijms241512092
Chicago/Turabian StyleDe la Cruz, Alicia, Xiaoan Wu, Quinn C. Rainer, Irene Hiniesto-Iñigo, Marta E. Perez, Isak Edler, Sara I. Liin, and H. Peter Larsson. 2023. "Pharmacological Screening of Kv7.1 and Kv7.1/KCNE1 Activators as Potential Antiarrhythmic Drugs in the Zebrafish Heart" International Journal of Molecular Sciences 24, no. 15: 12092. https://doi.org/10.3390/ijms241512092