
Effect of Exogenous Plant Growth Regulators and Rejuvenation Measures on the Endogenous Hormone and Enzyme Activity Responses of Acer mono Maxim in Cuttage Rooting

Abstract
:1. Introduction
2. Results
2.1. Rooting Characteristics, Rooting Rate, and Callus Rate of A. mono Cuttings
2.2. Changes in IAA Content during the Rooting Process of A. mono Cuttings
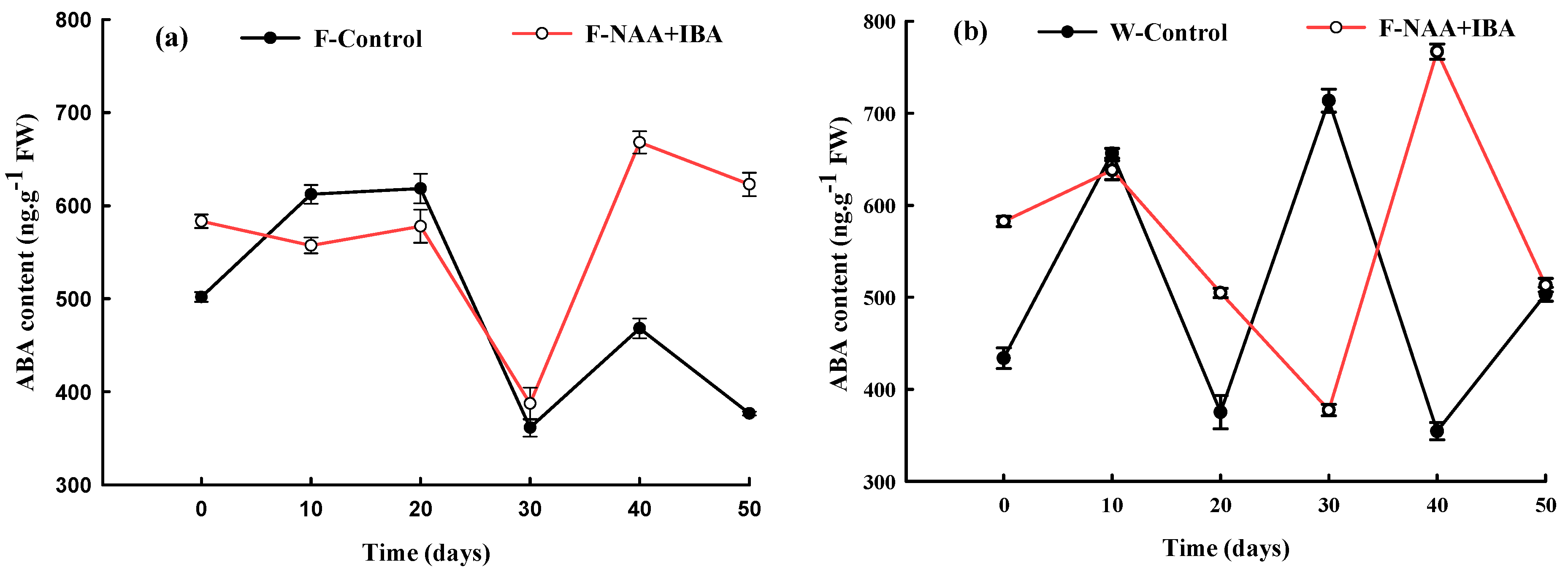
2.3. Changes in ABA Content during the Rooting Process of A. mono Cuttings
2.4. Changes in GA3 Content during the Rooting Process of A. mono Cuttings
2.5. Changes in ZR Content during the Rooting Process of A. mono Cuttings
2.6. Changes in POD Activity during the Rooting Process of A. mono Cuttings
2.7. Changes in PPO Activity during the Rooting Process of A. mono Cuttings
2.8. Changes in IAAO Activity during the Rooting Process of A. mono Cuttings
3. Discussion
3.1. Endogenous Hormone IAA Could Promote the Formation of Adventitious Roots in the Cuttings of A. mono
3.2. Endogenous Hormone ABA on Adventitious Roots in the Cuttings of A. mono
3.3. Endogenous Hormone GA3 Have Different Effects of Adventitious Roots in the Cuttings of A. mono
3.4. Endogenous Hormone ZR Conducive to the Growth and Development of Adventitious Roots of A. mono
3.5. POD Is Closely Related to the Induction and Growth of Adventitious Roots of A. mono
3.6. PPO Could Promote the Elongation of Adventitious Roots of A. mono
3.7. IAAO Could Control IAA Concentration and Promote the Growth and Development of Cuttings of A. mono
4. Materials and Methods
4.1. Plant Materials
4.2. Cutting Bed and Test Conditions
4.3. Collection and Production of Cuttings
4.4. Index Observation
4.5. Measurement Index and Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Z.J.; Sun, Y.H.; Li, Y. Plant rejuvenation: From phenotypes to mechanisms. Plant Cell Rep. 2020, 39, 1249–1262. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.P.; Tsuda, Y.; Shen, H.L.; Hu, L.J.; Saito, Y.; Ide, Y. Genetic structure and hierarchical population divergence history of Acer mono var. mono in South and Northeast China. PLoS ONE 2014, 9, e87187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, S.Q.; Zhang, M.Z.; Luo, Y.; Zhang, P.; Yang, L.; Shen, H.L. Investigation of germplasm resource of mono maple with purple-red autumn leaf traits. J. Chin. Urban For. 2013, 11, 20–23. [Google Scholar]
- Kang, W.; Li, R.; Wang, L.Y.; Cheng, H. Auxin-Induced adventitious root formation in nodal cuttings of camellia sinensis. Int. J. Mol. Sci. 2019, 20, 4817. [Google Scholar]
- Zhou, X.X.; Li, R.Y.; Zang, S.Y.; Kun, P.; Yang, L. Effects of exogenous auxin mass concentration and cuttings substrate on the rooting of the softwood cuttings of Acer mono. For. Eng. 2022, 38, 1–5. [Google Scholar]
- Shao, F.X.; Wang, S.; Huang, W.; Liu, Z.M. Effects of IBA on the rooting of branch cuttings of Chinese jujube (Zizyphus jujuba Mill.) and changes to nutrients and endogenous hormones. J. For. Res. 2018, 29, 1557–1567. [Google Scholar] [CrossRef]
- Li, H.H.; Pan, R.C. Hormone Control of Adventitious Rooting in Mung Bean Stem Cuttings: XV International Botanical Congress; Japan Pacifico Yokohama: Yokohama, Japan, 1993. [Google Scholar]
- Hou, J.W.; Guo, S.J.; Wang, G.Y. Effects of in vitro subculture on the physiological characteristics of adventitious root formation in microshoots of Castanea mollissima cv. ‘yanshanhong’. J. For. Res. 2010, 21, 155–160. [Google Scholar] [CrossRef]
- Rout, G.R. Effect of auxins on adventitious root development from single node cuttings of Camellia sinensis (L.) Kuntze and associated biochemical changes. Plant Growth Regul. 2006, 48, 111–117. [Google Scholar] [CrossRef]
- Gao, J.; Liu, X.; Yuan, Y. Water cultured propagation of Polygonum multiflorum and dynamic changes of physiological and biochemical characteristics during adventitious roots formation. China J. Chin. Mater. Med. 2011, 36, 375–378. [Google Scholar]
- Zhao, Y.Q.; Chen, Y.J.; Jiang, C.; Lu, M.Z.; Zhang, J. Exogenous hormones supplementation improve adventitious root formation in woody plants. Front. Bioeng. Biotechnol. 2022, 10, 1009531. [Google Scholar] [CrossRef]
- Wolters, H.; Jürgens, G. Survival of the flexible: Hormonal growth control and adaptation in plant development. Nat. Rev. Genet. 2009, 10, 305–317. [Google Scholar] [CrossRef]
- Zhou, A.P.; Zong, D.; Gan, P.H.; Zou, X.L.; Fei, X.; Zhong, Y.Y.; He, C.Z. Physiological analysis and transcriptome profiling of inverted cuttings of Populus yunnanensis reveal that cell wall metabolism plays a crucial role in responding to inversion. Genes 2018, 9, 572. [Google Scholar] [CrossRef] [Green Version]
- Husen, A.; Pal, M. Metabolic changes during adventitious root primordium development in Tectona grandis Linn. f.(teak) cuttings as affected by age of donor plants and auxin (IBA and NAA) treatment. New For. 2007, 33, 309–323. [Google Scholar] [CrossRef]
- Taramino, G.; Sauer, M.; Stauffer, J.L., Jr.; Multani, D.; Niu, X.; Sakai, H.; Hochholdinger, F. The maize (Zea mays L.) RTCS gene encodes a LOB domain protein that is a key regulator of embryonic seminal and post-embryonic shoot-borne root initiation. Plant J. 2007, 50, 649–659. [Google Scholar] [CrossRef]
- Caplan, D.; Stemeroff, J.; Dixon, M.; Zheng, Y.B. Vegetative propagation of cannabis by stem cuttings: Effects of leaf number, cutting position, rooting hormone, and leaf tip removal. Can. J. Plant Sci. 2018, 98, 1126–1132. [Google Scholar] [CrossRef]
- Shang, C.Q.; Yang, H.L.; Ma, S.; Shen, Q.D.; Liu, L.; Hou, C.X.; Cao, X.; Cheng, J.L. Physiological and transcriptomic changes during the early phases of adventitious root formation in mulberry stem hardwood cuttings. Int. J. Mol. Sci. 2019, 20, 3707. [Google Scholar] [CrossRef] [Green Version]
- Mu, H.Z.; Jin, X.H.; Ma, X.Y.; Zhao, A.Q.; Gao, Y.T.; Lin, L. Ortet Age Effect, Anatomy and Physiology of Adventitious Rooting in Tilia mandshurica Softwood Cuttings. Forests 2022, 13, 1427. [Google Scholar] [CrossRef]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.H.; Cao, F.; Liu, Z.Z.; Peng, F.R.; Liang, Y.W.; Tan, P.P. Effects of NAA treatment on the endogenous hormone changes in cuttings of Carya illinoinensis during rooting. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2016, 40, 191–196. [Google Scholar]
- Li, C.X.; Bian, B.; Gong, T.Y.; Liao, W.B. Comparative proteomic analysis of key proteins during abscisic acid-hydrogen peroxide-induced adventitious rooting in cucumber (Cucumis sativus L.) under drought stress. J. Plant Physiol. 2018, 229, 185–194. [Google Scholar] [CrossRef]
- Bollmark, M.; Eliasson, L. Ethylene accelerates the breakdown of cytokinins and thereby stimulates rooting in Norway spruce hypocotyl cuttings. Physiol. Plant. 1990, 80, 534–540. [Google Scholar] [CrossRef]
- Mauriat, M.; Petterle, A.; Bellini, C.; Moritz, T. Gibberellins inhibit adventitious rooting in hybrid aspen and Arabidopsis by affecting auxin transport. Plant J. 2014, 78, 372–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez-Carvajal, G.A.; Morse, A.M.; Dervinis, C.; Davis, J.M. The cytokinin type-B response regulator PtRR13 is a negative regulator of adventitious root development in Populus. Plant Physiol. 2009, 150, 759–771. [Google Scholar] [CrossRef] [Green Version]
- Uwe, D.; Philipp, F.; Hajirezaei, M.R. Plant hormone homeostasis, signaling, and function during adventitious root formation in cuttings. Front. Plant 2016, 7, 381. [Google Scholar]
- Han, J.H.; Shao, W.H.; Liu, J.F.; Diao, S.F. Content changes of endogenous hormone and polyphenols during hardwood cuttage progress in Sapindus ukorossi. Non-Wood For. Res. 2019, 37, 37–43. [Google Scholar]
- Lv, G.X.; Qing, J.; Du, H.Y.; Du, Q.X.; Meng, Y.D.; He, F.; Liu, P.F.; Du, L.Y.; Wang, L. Comparing rooting ability and physiological changes of two Eucommia ulmoides improved varieties. Forests 2021, 12, 1267. [Google Scholar] [CrossRef]
- Yilmaz, H.; Taşkin, T.; Otludil, B. Polyphenol oxidase activity during rooting in cuttings of grape (Vitis vinifera L.) varieties. Turk. J. Bot. 2003, 27, 495–498. [Google Scholar]
- Syros, T.; Yupsanis, T.; Zafiriadis, H.; Economou, A. Activity and isoforms of peroxidases, lignin and anatomy, during adventitious rooting in cuttings of Ebenus cretica L. J. Plant Physiol. 2004, 161, 69–77. [Google Scholar] [CrossRef]
- Xiong, F.; Liao, J.R.; Ma, Y.C.; Wang, Y.H.; Fang, W.P.; Zhu, X.J. The Protective Effect of Exogenous Putrescine in the Response of Tea Plants (Camellia sinensis) to Salt Stress. HortScience 2018, 53, 1640–1646. [Google Scholar] [CrossRef] [Green Version]
- Massolo, J.F.; Concellón, A.; Chaves, A.R.; Vicente, A.R. 1-Methylcyclopropene (1-MCP) delays senescence, maintains quality and reduces browning of non-climacteric eggplant (Solanum melongena L.) fruit. Postharvest Biol. Technol. 2011, 59, 10–15. [Google Scholar] [CrossRef]
- Qaddoury, A.; Amssa, M. Endogenous phenolic contents, peroxidase and polyphenoloxidase activities in date palm (Phoenix dactylifera L.) offshoots related to rooting ability. Acta Physiol. Plant. 2003, 25, 417–421. [Google Scholar] [CrossRef]
- Pan, X.; Zhao, Q.; Zhao, T. Plant growth, antioxidative enzyme, and cadmium tolerance responses to cadmium stress in Canna orchioides. Hortic. Plant J. 2021, 7, 256–266. [Google Scholar]
- Nag, S.; Saha, K.; Choudhuri, M.A. Role of auxin and polyamines in adventitious root formation in relation to changes in compounds involved in rooting. J. Plant Growth Regul. 2001, 20, 182–194. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Rooting Rate (%) | Callus Rate (%) |
|---|---|---|
| Rejuvenation Control | 4.17 ± 4.17 c | 25.00 ± 4.81 b |
| Rejuvenation NAA + IBA | 29.17 ± 4.17 b | 37.50 ± 7.98 b |
| Non-Rejuvenation Control | 4.17 ± 4.17 c | 16.67 ± 6.80 b |
| Non-Rejuvenation NAA + IBA | 62.50 ± 4.17 a | 66.67 ± 6.80 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Li, R.; Shen, H.; Yang, L. Effect of Exogenous Plant Growth Regulators and Rejuvenation Measures on the Endogenous Hormone and Enzyme Activity Responses of Acer mono Maxim in Cuttage Rooting. Int. J. Mol. Sci. 2023, 24, 11883. https://doi.org/10.3390/ijms241511883
Zhou X, Li R, Shen H, Yang L. Effect of Exogenous Plant Growth Regulators and Rejuvenation Measures on the Endogenous Hormone and Enzyme Activity Responses of Acer mono Maxim in Cuttage Rooting. International Journal of Molecular Sciences. 2023; 24(15):11883. https://doi.org/10.3390/ijms241511883
Chicago/Turabian StyleZhou, Xinxin, Ruyue Li, Hailong Shen, and Ling Yang. 2023. "Effect of Exogenous Plant Growth Regulators and Rejuvenation Measures on the Endogenous Hormone and Enzyme Activity Responses of Acer mono Maxim in Cuttage Rooting" International Journal of Molecular Sciences 24, no. 15: 11883. https://doi.org/10.3390/ijms241511883