
Transcriptome and Small RNA Sequencing Reveals the Basis of Response to Salinity, Alkalinity and Hypertonia in Quinoa (Chenopodium quinoa Willd.)
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
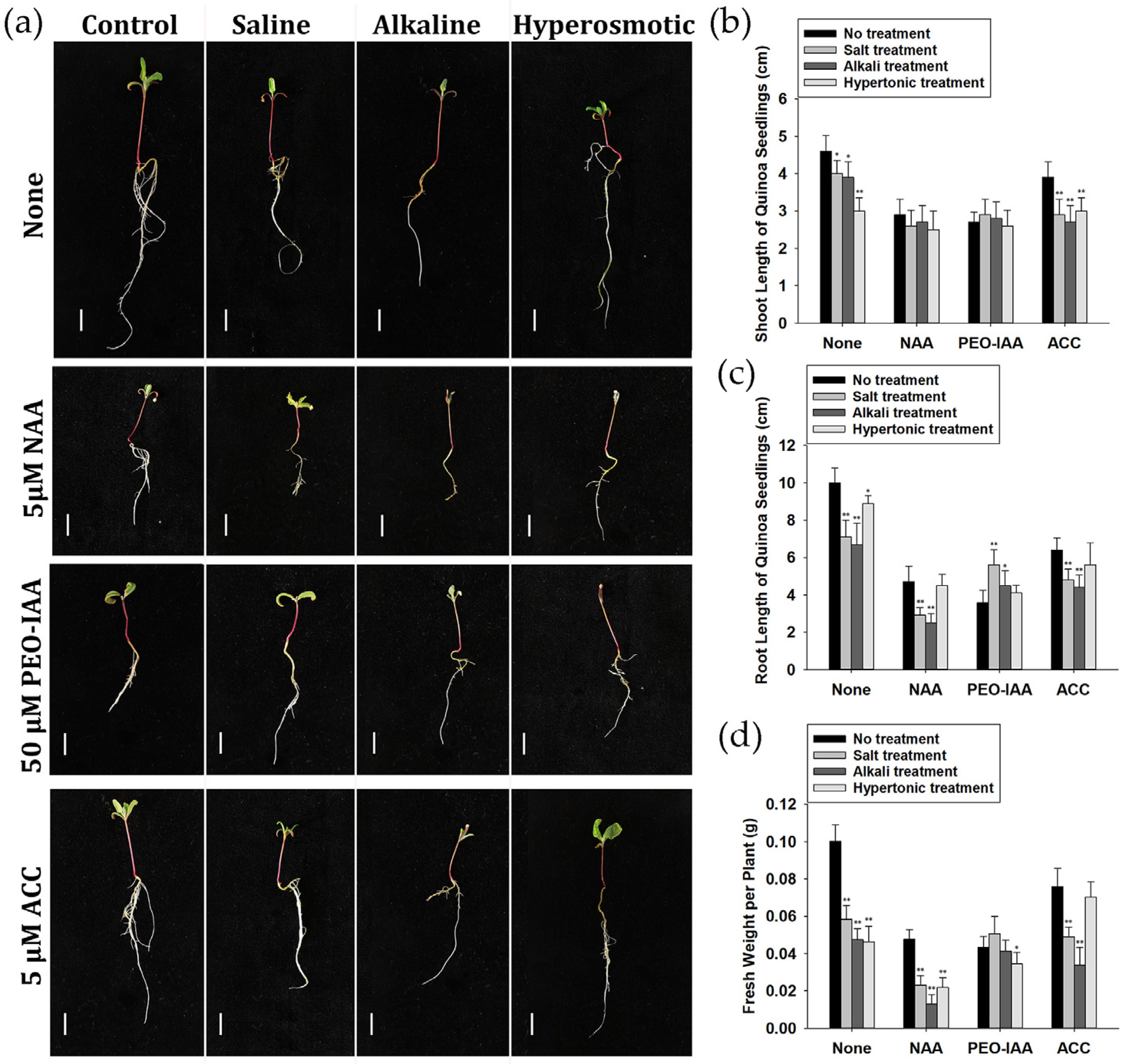
2.1. Response of Quinoa Strain CM499 to Saline, Alkaline and Hyperosmotic Stresses
2.2. Transcriptome Sequencing of CM499 Seedlings
2.3. Differential Expression Analysis of Quinoa Genes
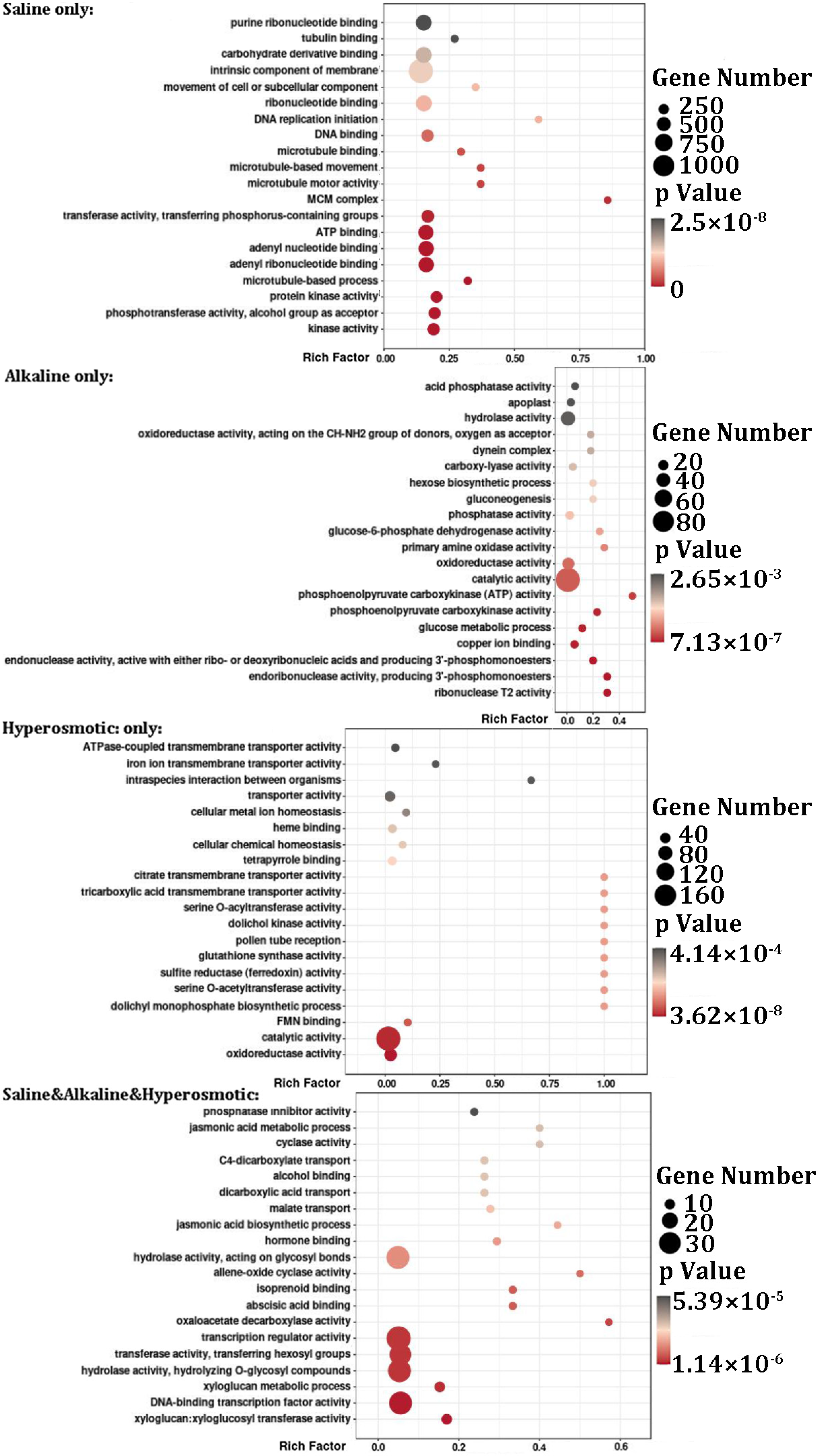
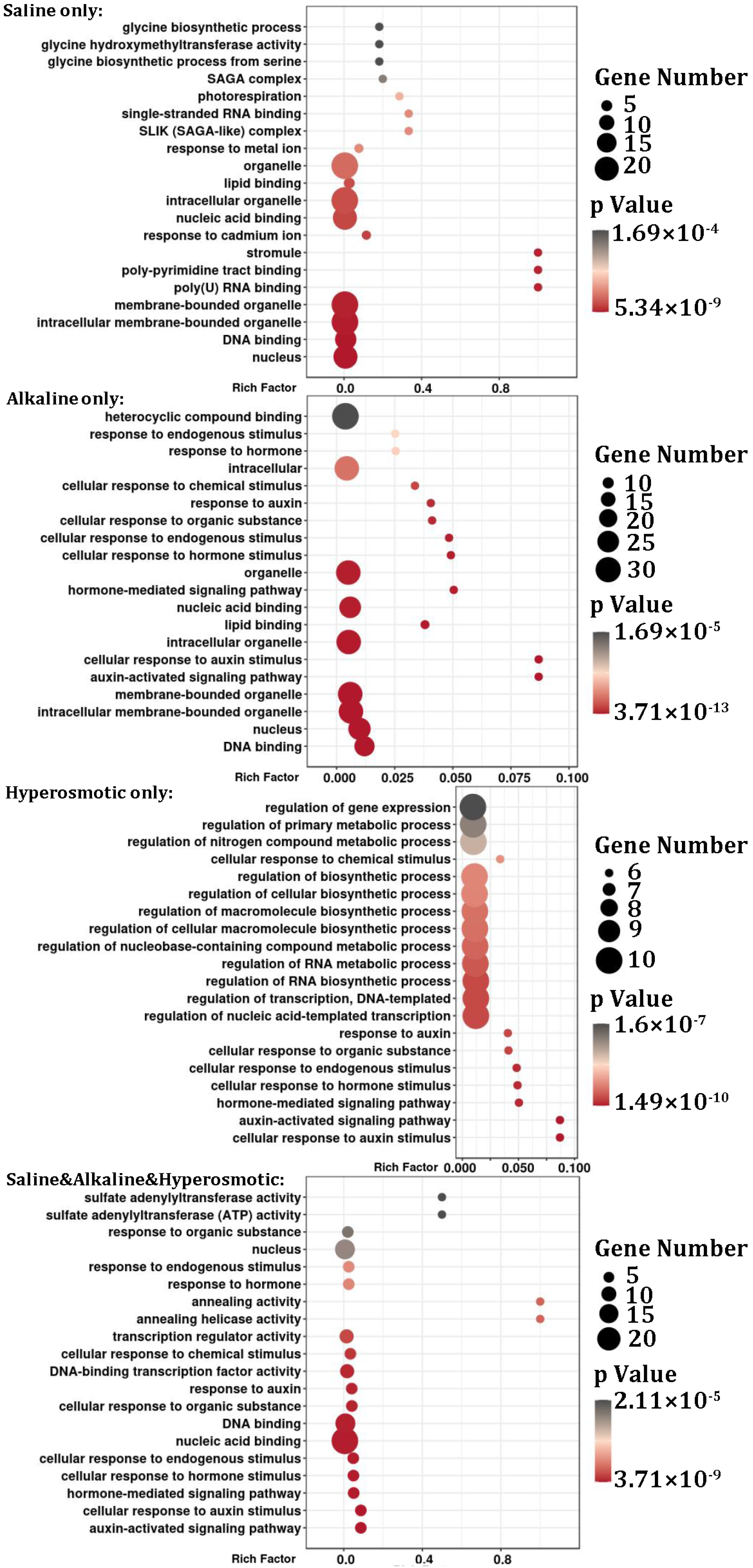
2.4. Functional Analysis of DEGs in Quinoa Stress Response
2.5. The Acquisition of Small RNA Sequence
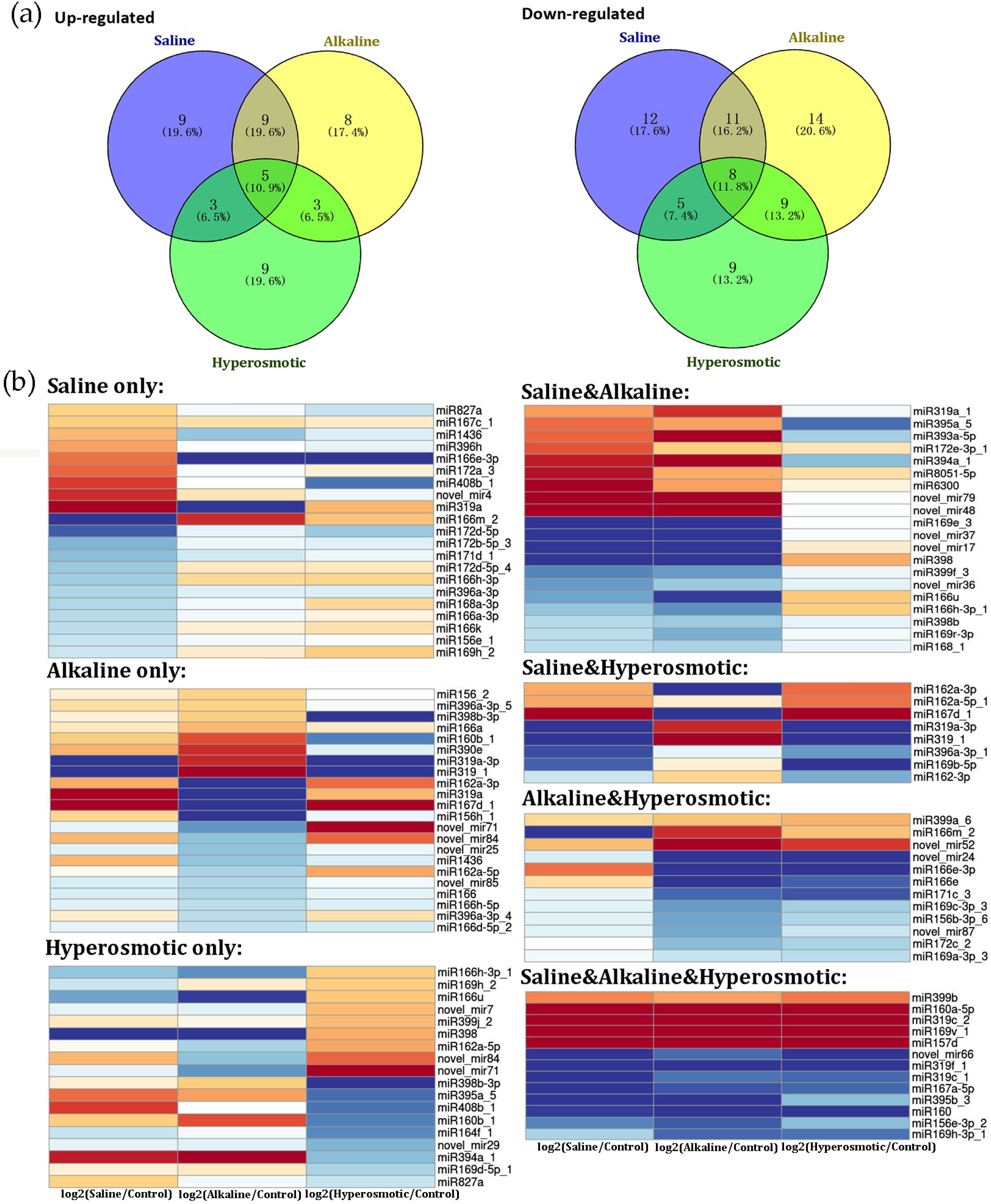
2.6. MiRNA Responses to Different Stresses in Quinoa
2.7. Functional Analysis of DEMs in Quinoa
2.8. ROS Accumulation in Quinoa Seedlings under Stress Treatment
2.9. Auxin and Ethylene Were Involved in Quinoa Stress Responses
2.10. Metabolism of Hemicellulose in Quinoa Stress Responses
3. Discussion
3.1. Plant Hormones Played Important Roles in Quinoa Stress Responses
3.2. Transcription Factors Were Important Regulators of Abiotic Stress Response in Quinoa
3.3. ROS Metabolism in Abiotic Stress Response of Quinoa
3.4. Cytoskeleton Dynamics Might Be Involved in Stress Responses in Quinoa
3.5. Cell Wall Organization Was Associated with Stress Response in Quinoa
4. Materials and Methods
4.1. Plant Materials and Growing Conditions
4.2. Growth Phenotype Observation
4.3. Transcriptome and Small RNA Sequencing
4.4. qRT-PCR Analysis
4.5. ROS Stain
4.6. Hemicellulose Determination
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ruiz, K.B.; Maldonado, J.; Biondi, S.; Silva, H. RNA-Seq Analysis of Salt-Stressed versus Non Salt-Stressed Transcriptomes of Chenopodium quinoa Landrace R49. Genes 2019, 10, 1042. [Google Scholar] [CrossRef] [Green Version]
- Peterson, A.; Murphy, K. Tolerance of Lowland Quinoa Cultivars to Sodium Chloride and Sodium Sulfate Salinity. Crop. Sci. 2015, 55, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Adolf, V.I.; Jacobsen, S.E.; Shabala, S. Salt Tolerance Mechanisms in Quinoa (Chenopodium quinoa Willd.). Environ. Exp. Bot. 2013, 92, 43–54. [Google Scholar] [CrossRef]
- Hinojosa, L.; Gonzalez, J.A.; Barrios-Masias, F.H.; Fuentes, F.; Murphy, K.M. Quinoa Abiotic Stress Responses: A Review. Plants 2018, 7, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanji, K.K.; American Society of Civil Engineers; Irrigation and Drainage Division; Water Quality Technical Committee. Agricultural Salinity Assessment and Management; Wallender, W.W., Tanji, K.K., Eds.; American Society of Civil Engineers: New York, NY, USA, 1990; p. xi. 619p. [Google Scholar]
- Wahome, P.K. Mechanisms of Salt (NaCl) Stress Tolerance in Horticultural Crops—A Mini Review. Acta Hortic. 2003, 609, 127–131. [Google Scholar] [CrossRef]
- Yang, C.W.; Xu, H.H.; Wang, L.L.; Liu, J.; Shi, D.C.; Wang, D.L. Comparative Effects of Salt-Stress and Alkali-Stress on the Growth, Photosynthesis, Solute Accumulation, and Ion Balance of Barley Plants. Photosynthetica 2009, 47, 79–86. [Google Scholar] [CrossRef]
- Farooqi, Z.U.R.; Ayub, M.A.; Zia ur Rehman, M.; Sohail, M.I.; Usman, M.; Khalid, H.; Naz, K. Regulation of Drought Stress in Plants. In Plant Life under Changing Environment; Tripathi, D.K., Pratap Singh, V., Chauhan, D.K., Sharma, S., Prasad, S.M., Dubey, N.K., Ramawat, N., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 77–104. [Google Scholar]
- Almeida, D.M.; Oliveira, M.M.; Saibo, N.J.M. Regulation of Na+ and K+ Homeostasis in Plants: Towards Improved Salt Stress Tolerance in Crop Plants. Genet. Mol. Biol. 2017, 40, 326–345. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.W.; Wang, C.; Sun, Y.; Wang, N.; Li, X.W.; Dong, Y.Y.; Yao, N.; Liu, X.M.; Chen, H.; Chen, X.F.; et al. Overexpression of Vacuolar Proton Pump Atpase (V-H+-Atpase) Subunits B, C and H Confers Tolerance to Salt and Saline-Alkali Stresses in Transgenic Alfalfa (Medicago sativa L.). J. Integr. Agric. 2016, 15, 2279–2289. [Google Scholar] [CrossRef]
- Guo-Hui, Y. Alkali Stress Induced the Accumulation and Secretion of Organic Acids in Wheat. Afr. J. Agric. Res. 2012, 7, 2844–2852. [Google Scholar] [CrossRef]
- You, J.; Chan, Z.L. ROS Regulation During Abiotic Stress Responses in Crop Plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [Green Version]
- Bakshi, P.; Handa, N.; Gautam, V.; Kaur, P.; Sareen, S.; Mir, B.; Bhardwaj, R. Role and Regulation of Plant Hormones as a Signal Molecule in Response to Abiotic Stresses. In Plant Signaling Molecules; Khan, M.I.R., Reddy, P.S., Ferrante, A., Khan, N.A., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 303–317. [Google Scholar]
- Sun, X.; Lin, L.; Sui, N. Regulation Mechanism of MicroRNA in Plant Response to Abiotic Stress and Breeding. Mol. Biol. Rep. 2019, 46, 1447–1457. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, D.E.; Ho, Y.S.; Lightfoot, D.J.; Schmockel, S.M.; Li, B.; Borm, T.J.A.; Ohyanagi, H.; Mineta, K.; Michell, C.T.; Saber, N.; et al. The Genome of Chenopodium quinoa. Nature 2017, 542, 307–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz, K.B.; Biondi, S.; Martinez, E.A.; Orsini, F.; Antognoni, F.; Jacobsen, S.E. Quinoa—A Model Crop for Understanding Salt-Tolerance Mechanisms in Halophytes. Plant Biosyst. 2016, 150, 357–371. [Google Scholar] [CrossRef]
- Maughan, P.J.; Turner, T.B.; Coleman, C.E.; Elzinga, D.B.; Jellen, E.N.; Morales, J.A.; Udall, J.A.; Fairbanks, D.J.; Bonifacio, A. Characterization of Salt Overly Sensitive 1 (SOS1) Gene Homoeologs in Quinoa (Chenopodium quinoa Willd.). Genome 2009, 52, 647–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orsini, F.; Accorsi, M.; Gianquinto, G.; Dinelli, G.; Antognoni, F.; Carrasco, K.B.R.; Martinez, E.A.; Alnayef, M.; Marotti, I.; Bosi, S.; et al. Beyond the Ionic and Osmotic Response to Salinity in Chenopodium quinoa: Functional Elements of Successful Halophytism. Funct. Plant Biol. 2011, 38, 818–831. [Google Scholar] [CrossRef]
- Becker, V.I.; Goessling, J.W.; Duarte, B.; Cacador, I.; Liu, F.L.; Rosenqvist, E.; Jacobsen, S.E. Combined Effects of Soil Salinity and High Temperature on Photosynthesis and Growth of Quinoa Plants (Chenopodium quinoa). Funct. Plant Biol. 2017, 44, 665–678. [Google Scholar] [CrossRef]
- Panuccio, M.R.; Jacobsen, S.E.; Akhtar, S.S.; Muscolo, A. Effect of Saline Water on Seed Germination and Early Seedling Growth of the Halophyte Quinoa. Aob Plants 2014, 6, plu047. [Google Scholar] [CrossRef]
- Pottosin, I.; Bonales-Alatorre, E.; Shabala, S. Choline but Not Its Derivative Betaine Blocks Slow Vacuolar Channels in the Halophyte Chenopodium quinoa: Implications for Salinity Stress Responses. FEBS Lett. 2014, 588, 3918–3923. [Google Scholar] [CrossRef]
- Ruiz, K.B.; Rapparini, F.; Bertazza, G.; Silva, H.; Torrigiani, P.; Biondi, S. Comparing Salt-Induced Responses at the Transcript Level in a Salares and Coastal-Lowlands Landrace of Quinoa (Chenopodium quinoa Willd). Environ. Exp. Bot. 2017, 140, 150. [Google Scholar] [CrossRef]
- Morales, A.; Bajgain, P.; Garver, Z.; Maughan, J.; Udall, J. Physiological Responses of Chenopodium quinoa to Salt Stress. Int. J. Plant Physiol. Biochem. 2011, 3, 219–232. [Google Scholar]
- Del Rio, L.A. ROS and RNS in Plant Physiology: An Overview. J. Exp. Bot. 2015, 66, 2827–2837. [Google Scholar] [CrossRef] [Green Version]
- Peleg, Z.; Blumwald, E. Hormone Balance and Abiotic Stress Tolerance in Crop Plants. Curr. Opin. Plant Biol. 2011, 14, 290–295. [Google Scholar] [CrossRef]
- Waadt, R.; Seller, C.A.; Hsu, P.K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant Hormone Regulation of Abiotic Stress Responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant Hormone-Mediated Regulation of Stress Responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [Green Version]
- Waśkiewicz, A.; Beszterda, M.; Goliński, P. Aba: Role in Plant Signaling under Salt Stress. In Salt Stress in Plants: Signalling, Omics and Adaptations; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer New York: New York, NY, USA, 2013; pp. 175–196. [Google Scholar]
- Pei, H.; Wang, H.; Wang, L.; Zheng, F.; Dong, C.-H. Regulatory Function of Ethylene in Plant Responses to Drought, Cold, and Salt Stresses. In Mechanism of Plant Hormone Signaling under Stress; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 327–344. [Google Scholar]
- Sharma, E.; Sharma, R.; Borah, P.; Jain, M.; Khurana, P. Emerging Roles of Auxin in Abiotic Stress Responses. In Elucidation of Abiotic Stress Signaling in Plants: Functional Genomics Perspectives; Springer: Berlin/Heidelberg, Germany, 2015; Volume 1, pp. 299–328. [Google Scholar]
- Mir, M.A.; John, R.; Alyemeni, M.N.; Alam, P.; Ahmad, P. Jasmonic Acid Ameliorates Alkaline Stress by Improving Growth Performance, Ascorbate Glutathione Cycle and Glyoxylase System in Maize Seedlings. Sci. Rep. 2018, 8, 2831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.A.; Nazar, R.; Iqbal, N.; Anjum, N.A. Phytohormones and Abiotic Stress Tolerance in Plants; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2012; p. viii. 306p. [Google Scholar]
- Kang, J.M.; Turano, F.J. The Putative Glutamate Receptor 1.1 (AtGLR1.1) Functions as a Regulator of Carbon and Nitrogen Metabolism in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2003, 100, 6872–6877. [Google Scholar] [CrossRef] [PubMed]
- McMichael, B.L.; Jordan, W.R.; Powell, R.D. An Effect of Water Stress on Ethylene Production by Intact Cotton Petioles. Plant Physiol. 1972, 49, 658–660. [Google Scholar] [CrossRef] [Green Version]
- Joo, J.; Choi, H.J.; Lee, Y.H.; Kim, Y.K.; Song, S.I. A Transcriptional Repressor of the ERF Family Confers Drought Tolerance to Rice and Regulates Genes Preferentially Located on Chromosome 11. Planta 2013, 238, 155–170. [Google Scholar] [CrossRef]
- Zhao, X.C.; Schaller, G.E. Effect of Salt and Osmotic Stress Upon Expression of the Ethylene Receptor ETR1 in Arabidopsis thaliana. FEBS Lett. 2004, 562, 189–192. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.H.; Liu, J.; He, X.J.; Mu, R.L.; Zhou, H.L.; Chen, S.Y.; Zhang, J.S. Modulation of Ethylene Responses Affects Plant Salt-Stress Responses. Plant Physiol. 2007, 143, 707–719. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.X.; Li, Z.F.; Quan, R.D.; Li, G.J.; Wang, R.G.; Huang, R.F. An AP2 Domain-Containing Gene, ESE1, Targeted by the Ethylene Signaling Component Ein3 Is Important for the Salt Response in Arabidopsis. Plant Physiol. 2011, 157, 854–865. [Google Scholar] [CrossRef] [Green Version]
- Asensi-Fabado, M.A.; Cela, J.; Muller, M.; Arrom, L.; Chang, C.R.; Munne-Bosch, S. Enhanced Oxidative Stress in the Ethylene-Insensitive (ein3-1) Mutant of Arabidopsis thaliana Exposed to Salt Stress. J. Plant Physiol. 2012, 169, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Woodrow, P.; Pontecorvo, G.; Ciarmiello, L.F.; Annunziata, M.G.; Fuggi, A.; Carillo, P. Transcription Factors and Genes in Abiotic Stress. In Crop Stress and Its Management: Perspectives and Strategies; Springer: Berlin/Heidelberg, Germany, 2012; pp. 317–357. [Google Scholar]
- Li, C.N.; Ng, C.K.Y.; Fan, L.M. Myb Transcription Factors, Active Players in Abiotic Stress Signaling. Environ. Exp. Bot. 2015, 114, 80–91. [Google Scholar] [CrossRef]
- Cao, L.; Yu, Y.; Ding, X.D.; Zhu, D.; Yang, F.; Liu, B.D.; Sun, X.L.; Duan, X.B.; Yin, K.D.; Zhu, Y.M. The Glycine soja NAC Transcription Factor GsNAC019 Mediates the Regulation of Plant Alkaline Tolerance and ABA Sensitivity. Plant Mol. Biol. 2017, 95, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.; Singh, S.; Singh, M.; Srivastava, P.K.; Mishra, R.K.; Singh, V.P.; Prasad, S.M. Transcriptional Regulation of Salinity Stress in Plants: A Short Review. Plant Gene 2017, 11, 160–169. [Google Scholar] [CrossRef]
- Ju, Y.L.; Yue, X.F.; Min, Z.; Wang, X.H.; Fang, Y.L.; Zhang, J.X. VvNAC17, a Novel Stress-Responsive Grapevine (Vitis vinifera L.) NAC Transcription Factor, Increases Sensitivity to Abscisic Acid and Enhances Salinity, Freezing, and Drought Tolerance in Transgenic Arabidopsis. Plant Physiol. Biochem. 2020, 146, 98–111. [Google Scholar] [CrossRef]
- Jung, S.E.; Kim, T.H.; Shim, J.S.; Bang, S.W.; Bin Yoon, H.; Oh, S.H.; Kim, Y.S.; Oh, S.J.; Seo, J.S.; Kim, J.K. Rice NAC17 Transcription Factor Enhances Drought Tolerance by Modulating Lignin Accumulation. Plant Sci. 2022, 323, 111404. [Google Scholar] [CrossRef]
- Rathnayake, K.; Garcia, T.; Cushman, J.; Wone, B. A Novel NAC83 Transcription Factor from Kalanchoe Fedtschenkoi Enhances Drought and Salt Tolerance in Arabidopsis. In Proceedings of the Plant and Animal Genome XXVII Conference, San Diego, CA, USA, 12–16 January 2019. [Google Scholar]
- Xu, Z.Y.; Gongbuzhaxi; Wang, C.Y.; Xue, F.; Zhang, H.; Ji, W.Q. Wheat NAC Transcription Factor TaNAC29 Is Involved in Response to Salt Stress. Plant Physiol. Biochem. 2015, 96, 356–363. [Google Scholar] [CrossRef]
- Patil, M.; Ramu, S.V.; Jathish, P.; Sreevathsa, R.; Reddy, P.C.; Prasad, T.G.; Udayakumar, M. Overexpression of AtNAC2 (ANAC092) in Groundnut (Arachis hypogaea L.) Improves Abiotic Stress Tolerance. Plant Biotechnol. Rep. 2014, 8, 161–169. [Google Scholar] [CrossRef]
- Jin, H.X.; Huang, F.; Cheng, H.; Song, H.N.; Yu, D.Y. Overexpression of the GmNAC2 Gene, an NAC Transcription Factor, Reduces Abiotic Stress Tolerance in Tobacco. Plant Mol. Biol. Rep. 2013, 31, 435–442. [Google Scholar] [CrossRef]
- Shen, J.B.; Lv, B.; Luo, L.Q.; He, J.M.; Mao, C.J.; Xi, D.D.; Ming, F. The NAC-Type Transcription Factor OsNAC2 Regulates ABA-Dependent Genes and Abiotic Stress Tolerance in Rice. Sci. Rep. 2017, 7, 40641. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.T.; Cao, L.F.; Hu, K.X.; Yu, X.Y.; Qu, S.C. MiR164-NAC21/22 Module Regulates the Resistance of Malus hupehensis against Alternaria alternata by Controlling Jasmonic Acid Signaling. Plant Sci. 2023, 330, 111635. [Google Scholar] [CrossRef] [PubMed]
- Burian, A.; Raczynska-Szajgin, M.; Borowska-Wykret, D.; Piatek, A.; Aida, M.; Kwiatkowska, D. The Cup-Shaped Cotyledon 2 and 3 Genes Have a Post-Meristematic Effect on Arabidopsis thaliana Phyllotaxis. Ann. Bot. 2015, 115, 807–820. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.G.; Shi, J.L.; Cao, J.L.; He, M.Y.; Wang, Y.J. VpWRKY3, a Biotic and Abiotic Stress-Related Transcription Factor from the Chinese Wild Vitis pseudoreticulata. Plant Cell Rep. 2012, 31, 2109–2120. [Google Scholar] [CrossRef] [PubMed]
- Ning, W.F.; Zhai, H.; Yu, J.Q.; Liang, S.; Yang, X.; Xing, X.Y.; Huo, J.L.; Pang, T.; Yang, Y.L.; Bai, X. Overexpression of Glycine soja WRKY20 Enhances Drought Tolerance and Improves Plant Yields under Drought Stress in Transgenic Soybean. Mol. Breed. 2017, 37, 19. [Google Scholar] [CrossRef]
- Scarpeci, T.E.; Zanor, M.I.; Mueller-Roeber, B.; Valle, E.M. Overexpression of AtWRKY30 Enhances Abiotic Stress Tolerance During Early Growth Stages in Arabidopsis thaliana. Plant Mol. Biol. 2013, 83, 265–277. [Google Scholar] [CrossRef]
- Cai, R.H.; Zhao, Y.; Wang, Y.F.; Lin, Y.X.; Peng, X.J.; Li, Q.; Chang, Y.W.; Jiang, H.Y.; Xiang, Y.; Cheng, B.J. Overexpression of a Maize WRKY58 Gene Enhances Drought and Salt Tolerance in Transgenic Rice. Plant Cell Tissue Organ. 2014, 119, 565–577. [Google Scholar] [CrossRef]
- Lopez-Galiano, M.J.; Gonzalez-Hernandez, A.I.; Crespo-Salvador, O.; Rausell, C.; Real, M.D.; Escamilla, M.; Camanes, G.; Garcia-Agustin, P.; Gonzalez-Bosch, C.; Garcia-Robles, I. Epigenetic Regulation of the Expression of WRKY75 Transcription Factor in Response to Biotic and Abiotic Stresses in Solanaceae Plants. Plant Cell Rep. 2018, 37, 167–176. [Google Scholar] [CrossRef]
- Vannini, C.; Iriti, M.; Bracale, M.; Locatelli, F.; Faoro, F.; Croce, P.; Pirona, R.; Di Maro, A.; Coraggio, I.; Genga, A. The Ectopic Expression of the Rice OsMYB4 Gene in Arabidopsis Increases Tolerance to Abiotic, Environmental and Biotic Stresses. Physiol. Mol. Plant Pathol. 2006, 69, 26–42. [Google Scholar] [CrossRef]
- Cai, H.S.; Tian, S.; Liu, C.L.; Dong, H.S. Identification of a MYB3R Gene Involved in Drought, Salt and Cold Stress in Wheat (Triticum aestivum L.). Gene 2011, 485, 146–152. [Google Scholar] [CrossRef]
- Jaradat, M.R.; Feurtado, J.A.; Huang, D.Q.; Lu, Y.Q.; Cutler, A.J. Multiple Roles of the Transcription Factor AtMYBR1/AtMYB44 in ABA Signaling, Stress Responses, and Leaf Senescence. BMC Plant Biol. 2013, 13, 192. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Wang, Y.H.; Zheng, H.; Lu, W.; Wu, C.G.; Huang, J.G.; Yan, K.; Yang, G.D.; Zheng, C.C. Salt-Induced Transcription Factor MYB74 Is Regulated by the RNA-Directed DNA Methylation Pathway in Arabidopsis. J. Exp. Bot. 2015, 66, 5997–6008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.N.; Yang, X.Y.; Guo, S.; Wang, Z.H.; Zhang, Z.H.; Fang, Z.M. MiR319-Targeted OsTCP21 and OsgaMYB Regulate Tillering and Grain Yield in Rice. J. Integr. Plant Biol. 2021, 63, 1260–1272. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Mittler, R. ROS-Induced ROS Release in Plant and Animal Cells. Free Radic. Biol. Med. 2018, 122, 21–27. [Google Scholar] [CrossRef]
- Han, H.N.; Wang, Q.; Wei, L.; Liang, Y.; Dai, J.L.; Xia, G.M.; Liu, S.W. Small Rna and Degradome Sequencing Used to Elucidate the Basis of Tolerance to Salinity and Alkalinity in Wheat. BMC Plant Biol. 2018, 18, 195. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, L.J.; Huang, R.D. Cytoskeleton and Plant Salt Stress Tolerance. Plant Signal Behav. 2011, 6, 29–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasteneys, G.O. Progress in Understanding the Role of Microtubules in Plant Cells. Curr. Opin. Plant Biol. 2004, 7, 651–660. [Google Scholar] [CrossRef]
- Qu, Y.N.; Song, P.; Hu, Y.W.; Jin, X.; Jia, Q.R.; Zhang, X.D.; Chen, L.; Zhang, Q. Regulation of Stomatal Movement by Cortical Microtubule Organization in Response to Darkness and ABA Signaling in Arabidopsis. Plant Growth Regul. 2018, 84, 467–479. [Google Scholar] [CrossRef]
- Li, J.; Arieti, R.; Staiger, C.J. Actin Filament Dynamics and Their Role in Plant Cell Expansion. In Plant Cell Wall Patterning and Cell Shape; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; pp. 127–162. [Google Scholar]
- Li, J.; Yu, D.; Qanmber, G.; Lu, L.; Wang, L.; Zheng, L.; Liu, Z.; Wu, H.; Liu, X.; Chen, Q.; et al. GhKLCR1, a Kinesin Light Chain-Related Gene, Induces Drought-Stress Sensitivity in Arabidopsis. Sci. China Life Sci. 2019, 62, 63–75. [Google Scholar] [CrossRef]
- Liu, B.; Fan, J.; Zhang, Y.; Mu, P.; Wang, P.; Su, J.; Lai, H.; Li, S.; Feng, D.; Wang, J.; et al. Ospfa-Dsp1, a Rice Protein Tyrosine Phosphatase, Negatively Regulates Drought Stress Responses in Transgenic Tobacco and Rice Plants. Plant Cell Rep. 2012, 31, 1021–1032. [Google Scholar] [CrossRef]
- Le Gall, H.; Philippe, F.; Domon, J.M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell Wall Metabolism in Response to Abiotic Stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef]
- Liu, Y.B.; Lu, S.M.; Zhang, J.F.; Liu, S.; Lu, Y.T. A Xyloglucan Endotransglucosylase/Hydrolase Involves in Growth of Primary Root and Alters the Deposition of Cellulose in Arabidopsis. Planta 2007, 226, 1547–1560. [Google Scholar] [CrossRef]
- Cho, S.K.; Kim, J.E.; Park, J.A.; Eom, T.J.; Kim, W.T. Constitutive Expression of Abiotic Stress-Inducible Hot Pepper CaXTH3, Which Encodes a Xyloglucan Endotransglucosylase/Hydrolase Homolog, Improves Drought and Salt Tolerance in Transgenic Arabidopsis Plants. FEBS Lett. 2006, 580, 3136–3144. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.W.; Huang, Y.; He, H.; Han, T.; Di, P.C.; Sechet, J.; Fang, L.; Liang, Y.; Scheller, H.B.; Mortimer, J.C.; et al. Xyloglucan Endotransglucosylase-Hydrolase 30 Negatively Affects Salt Tolerance in Arabidopsis. J. Exp. Bot. 2019, 70, 5495–5506. [Google Scholar] [CrossRef] [PubMed]
- Evers, M.; Huttner, M.; Dueck, A.; Meister, G.; Engelmann, J.C. Mira: Adaptable Novel MiRNA Identification in Plants Using Small RNA Sequencing Data. BMC Bioinform. 2015, 16, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.J.; Ma, Y.K.; Chen, T.; Wang, M.; Wang, X.J. Psrobot: A Web-Based Plant Small RNA Meta-Analysis Toolbox. Nucleic Acids Res. 2012, 40, W22–W28. [Google Scholar] [CrossRef] [PubMed]
- Fahlgren, N.; Carrington, J.C. MiRNA Target Prediction in Plants. Methods Mol. Biol. 2010, 592, 51–57. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, H.; Qu, Y.; Wang, Y.; Zhang, Z.; Geng, Y.; Li, Y.; Shao, Q.; Zhang, H.; Ma, C. Transcriptome and Small RNA Sequencing Reveals the Basis of Response to Salinity, Alkalinity and Hypertonia in Quinoa (Chenopodium quinoa Willd.). Int. J. Mol. Sci. 2023, 24, 11789. https://doi.org/10.3390/ijms241411789
Han H, Qu Y, Wang Y, Zhang Z, Geng Y, Li Y, Shao Q, Zhang H, Ma C. Transcriptome and Small RNA Sequencing Reveals the Basis of Response to Salinity, Alkalinity and Hypertonia in Quinoa (Chenopodium quinoa Willd.). International Journal of Molecular Sciences. 2023; 24(14):11789. https://doi.org/10.3390/ijms241411789
Chicago/Turabian StyleHan, Huanan, Yusen Qu, Yingcan Wang, Zaijie Zhang, Yuhu Geng, Yuanyuan Li, Qun Shao, Hui Zhang, and Changle Ma. 2023. "Transcriptome and Small RNA Sequencing Reveals the Basis of Response to Salinity, Alkalinity and Hypertonia in Quinoa (Chenopodium quinoa Willd.)" International Journal of Molecular Sciences 24, no. 14: 11789. https://doi.org/10.3390/ijms241411789