
Perfect Complementarity Mechanism for Aphid Control: Oligonucleotide Insecticide Macsan-11 Selectively Causes High Mortality Rate for Macrosiphoniella sanborni Gillette
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin of Material
2.2. Sequence Development
2.3. DNA Synthesis
2.4. Sample Preparation and PCR
2.5. Nuclease Activity
2.6. Statistics
2.7. Photos
2.8. Sequencing
3. Results
3.1. Synthesis of Oligonucleotides
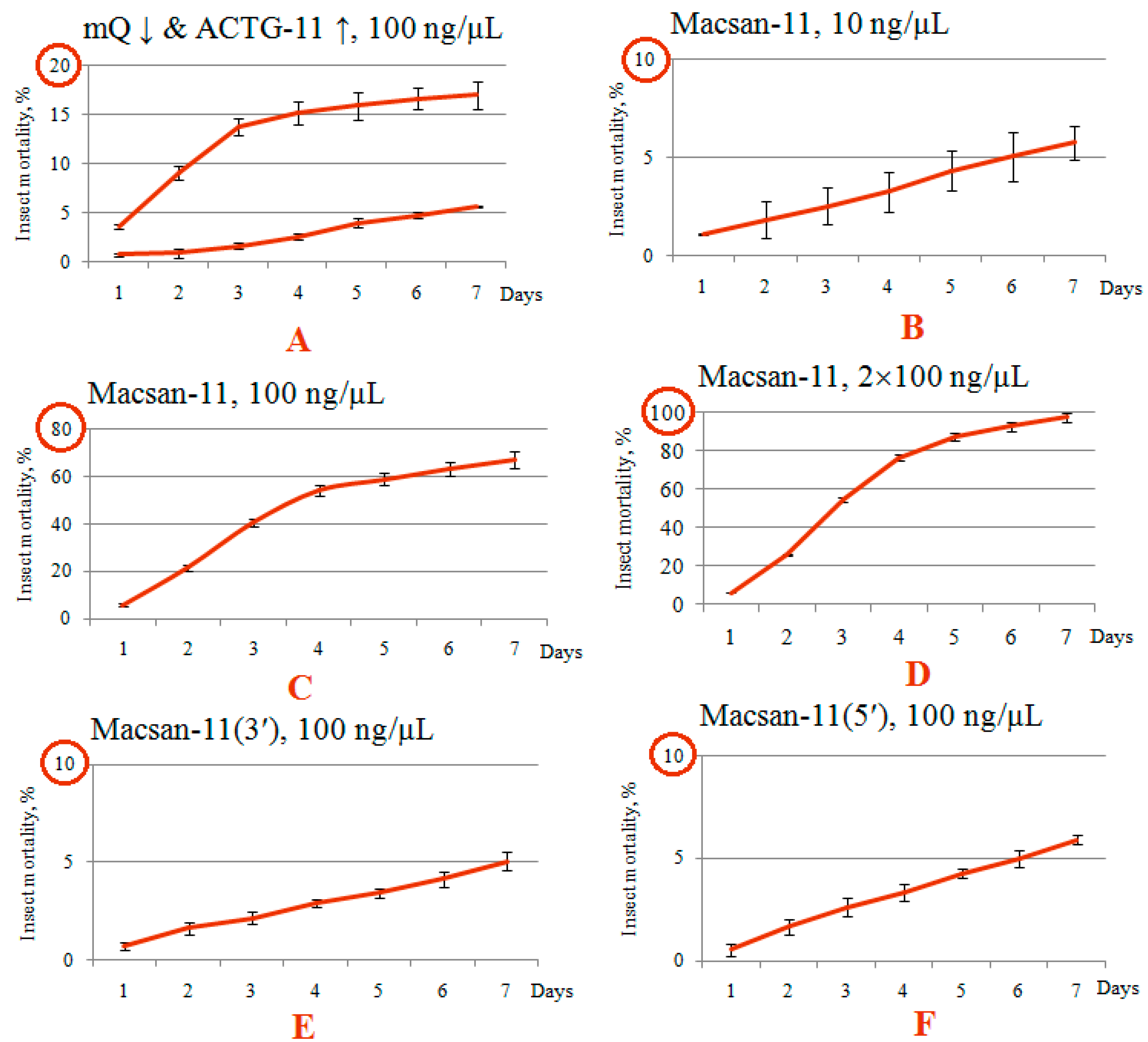
3.2. Evaluation of the Insecticidal Effect of Macsan-11 with Perfect Complementarity to the Target
3.3. Pronounced Decrease in Insecticidal Effect for Olinscides without Perfect Complementarity to the Target
3.4. Assessment of Nuclease Activity
3.5. Evaluation of Concentration of Target RNA
3.6. Sequencing of the Target Fragment of ITS2 in Chrysanthemum Aphids
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oberemok, V.; Laikova, K.; Useinov, R.; Gal’chinsky, N.; Novikov, I.; Gorlov, M. High Mortality of Sap-sucking Insects One Week After Topical Application of DNA Insecticides. Vitr. Cell. Dev. Biol.–Anim. 2020, 56, 31–39. [Google Scholar] [CrossRef]
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops: An Identification and Information Guide; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 1984. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.K.; Forbes, A.R.; Raworth, D.A. Aphid-transmitted viruses and their vectors of the world. Agric. Canada Res. Branch Tech. Bull. 1991, 3E, 216. [Google Scholar]
- Chatzivassiliou, E.K.; Papapanagiotou, A.P.; Mpenardis, P.D.; Perdikis, D.C.; Menexes, G. Transmission of Moroccan watermelon mosaic virus (MWMV) by aphids in Greece. Plant Dis. 2016, 100, 601–606. [Google Scholar] [CrossRef] [Green Version]
- Volesky, N.; Schrumm, Z.R. High Tunnel Pest Management-Aphids. Utah Pest Fact Sheet. Utta State Univ. 2021, N. ENT–225–21–PR, 1–7. [Google Scholar]
- Zhong, J.; Wang, Y.; Lu, Y.; Ma, X.; Zhang, Q.; Wang, X.; Zhang, Q.; Sun, M. Identification and Expression Analysis of Chemosensory Genes in the Antennal Transcriptome of Chrysanthemum Aphid Macrosiphoniella sanborni. Insects 2022, 13, 597. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Gao, T.; Li, P.; Tian, C.; Song, A.; Jiang, J.; Guan, Z.; Fang, W.; Chen, F.; Chen, S. Chrysanthemum CmWRKY53 negatively regulates the resistance of chrysanthemum to the aphid Macrosiphoniella sanborni. Hortic. Res. 2020, 7, 109. [Google Scholar] [CrossRef] [PubMed]
- Rene, E.R.; Bui, X.T.; Ngo, H.H.; Nghiem, L.D.; Guo, W. Green technologies for sustainable environment: An introduction. Environ. Sci. Pollut. Res. Int. 2021, 28, 63437–63439. [Google Scholar] [CrossRef]
- Dhakal, R.; Ghimire, R.; Sapkota, M.; Thapa, S.; Bhatta, A.K.; Regmi, R. Bioefficacy of different insecticides on cowpea aphid (Aphis craccivora Koch). Int. J. Entomol. Res. 2019, 7, 1–7. [Google Scholar] [CrossRef]
- Naveena, J.B. Sharanabasappa Effect of different insecticides against aphids, Aphis gossypii and whiteflies, Bemisia tabaci. J. Pharmacog. Phytochem. 2019, 8, 1572–1573. [Google Scholar]
- Rahardjo, I.B.; Hutapea, D.; Marwoto, B.; Budiarto, K. Effects of Several Botanical Insecticides Applied in Different Periods to Control Aphids (Macrosiphoniella sanborni Gillette) on Chrysanthemum. AGRIVITAJ Agricult. Sci. 2021, 43, 495–506. [Google Scholar] [CrossRef]
- Fan, J.; Zhang, X.Y.; Sun, X.Z.; Xu, B.Y. Effect of methyl jasmonate on aphid resistance of chrysanthemum. Ying Yong Sheng Tai XueBao 2020, 31, 4197–4205. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Shen, J.; Zhou, Y.; Wei, Z.P.; Gao, J.M. Insecticidal Constituents from Buddlej aalbiflora Hemsl. Nat. Prod. Res. 2017, 31, 1446–1449. [Google Scholar] [CrossRef] [PubMed]
- Ikbal, C.; Pavela, R. Essential oils as active ingredients of botanical insecticides against aphids. J. Pest. Sci. 2019, 92, 971–986. [Google Scholar] [CrossRef]
- Emam, A.K. Biological Control of the Chrysanthemum Aphid, Macrosiphoniellasanborni (Gillete) by Release Coccinella septempunctata L. on Chrysanthemum Plants. Pathol. 2016, 7, 291–295. [Google Scholar] [CrossRef]
- Helyer, N.; Gill, G.; Bywater, A. Elevated humidities for control of chrysanthemum pests with Verticillium lecanii. Pesticide Sci. 1992, 3, 373–378. [Google Scholar] [CrossRef]
- Sudan, Y.E.; Yuhui, D.; Feng, M.G. Time and concentration dependent interactions of Beauveria bassiana with sublethal rates of imidacloprid against the aphid pests Macrosiphoniella sanborni and Myzus persicae. Ann. Appl. Bio. 2005, 146, 459–468. [Google Scholar] [CrossRef]
- Schulz, R.; Bub, S.; Petschick, L.L.; Stehle, S.; Wolfram, J. Applied pesticide toxicity shifts toward plants and invertebrates, even in GM crops. Science 2021, 372, 81–84. [Google Scholar] [CrossRef]
- Kaleem Ullah, R.M.; Gao, F.; Sikandar, A.; Wu, H. Insights into the Effects of Insecticides on Aphids (Hemiptera: Aphididae): Resistance Mechanisms and Molecular Basis. Int. J. Mol. Sci. 2023, 24, 6750. [Google Scholar] [CrossRef] [PubMed]
- Sekamatte, M.B.; Ogenga-Latigo, M. Efficacy and Impact of Some Insecticides Used to Control Aphids, Aphis gossypii Glover (Homopters: Aphididae) on Cotton in Uganda, on Predators. In Proceedings of the World Cotton Research Conference-2, Athens, Greece, 6–12 September 1998; pp. 724–728. [Google Scholar]
- Sial, M.U.; Mehmood, K.; Saeed, S.; Husain, M.; Rasool, K.G.; Aldawood, A.S. Neonicotinoid’s resistance monitoring, diagnostic mechanisms and cytochrome P450 expression in green peach aphid [Myzus persicae (Sulzer) (Hemiptera: Aphididae)]. PLoS ONE 2022, 17, e0261090. [Google Scholar] [CrossRef]
- Umina, P.A.; Edwards, O.; Carson, P.; Van Rooyen, A.; Anderson, A. High levels of resistance to carbamate and pyrethroid chemicals widespread in Australian Myzus persicae (Hemiptera: Aphididae) populations. J. Econ. Entomol. 2014, 107, 1626–1638. [Google Scholar] [CrossRef]
- Oberemok, V.V. Ukrainian Patent Application. No. 36445, 27 October 2008. [Google Scholar]
- Hunter, W.B. Improving Suppression of Hemipteran Vectors and Bacterial Pathogens of Citrus and Solanaceous Plants: Advances in Antisense Oligonucleotides (FANA). Front. Agron. 2021, 3, 675247. [Google Scholar] [CrossRef]
- Sandoval-Mojica, A.F. Antibacterial FANA oligonucleotides as a novel approach for managing the Huanglongbing pathosystem. Sci. Rep. 2021, 11, 2760. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Oberemok, V.V.; Useinov, R.Z.; Skorokhod, O.A.; Gal’chinsky, N.V.; Novikov, I.A.; Makalish, T.P.; Yatskova, E.V.; Sharmagiy, A.K.; Golovkin, I.O.; Gninenko, Y.I. Oligonucleotide Insecticides for Green Agriculture: Regulatory Role of Contact DNA in Plant-Insect Interactions. Int. J. Mol. Sci. 2022, 23, 15681. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence | Target | |
|---|---|---|
| Macsan-11 | 5′-TGTGTTCGTTA-3′ | ITS2 1 of polycistronic rRNA transcript |
| Macsan-11(3′) | 5′-TGTGTTCGTTT-3′ | ITS2 (changed A to T at 3′ end of Macsan-11) |
| Macsan-11(5′) | 5′-AGTGTTCGTTA-3′ | ITS2 (changed T to A at 5′ end of Macsan-11) |
| ACTG-11 | 5′-ACTGACTGACT-3′ | Identification of the insecticidal effect of a random DNA oligonucleotide |
| Agex_F | 5′-TGCAAGTGCGCTTCCACTTA-3′ | Forward primer for sequencing and concentration evaluation/ITS2 |
| Agex_R | 5′-TAGCAGACGAAACGGCAGAT-3′ | Reverse primer for concentration evaluation/ITS2 |
| Agoss_R | 5′-ACGGGGACATCGTGATTTTG-3′ | Reverse primer for sequencing/ITS2 |
| Name | Theoretical m/z Ratio | Resulting m/z Ratio | |
|---|---|---|---|
| Olinscides | Macsan-11 | 3343.57 | 3345.93 |
| Macsan-11(3′) | 3343.57 | 3339.26 | |
| Macsan-11(5′) | 3361.60 | 3367.41 | |
| ACTG-11 | 3315.60 | 3321.04 | |
| Primers | Agex_F | 6067.05 | 6062.50 |
| Agex_R | 6183.10 | 6178.09 | |
| Agoss_R | 6187.06 | 6192.74 |
| Day | Control (mQ Water) | Macsan-11 |
|---|---|---|
| 1st | 5.20 ± 0.01 | 5.18 ± 0.03 |
| 7th | 5.21 ± 0.05 | 5.20 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puzanova, Y.V.; Novikov, I.A.; Bilyk, A.I.; Sharmagiy, A.K.; Plugatar, Y.V.; Oberemok, V.V. Perfect Complementarity Mechanism for Aphid Control: Oligonucleotide Insecticide Macsan-11 Selectively Causes High Mortality Rate for Macrosiphoniella sanborni Gillette. Int. J. Mol. Sci. 2023, 24, 11690. https://doi.org/10.3390/ijms241411690
Puzanova YV, Novikov IA, Bilyk AI, Sharmagiy AK, Plugatar YV, Oberemok VV. Perfect Complementarity Mechanism for Aphid Control: Oligonucleotide Insecticide Macsan-11 Selectively Causes High Mortality Rate for Macrosiphoniella sanborni Gillette. International Journal of Molecular Sciences. 2023; 24(14):11690. https://doi.org/10.3390/ijms241411690
Chicago/Turabian StylePuzanova, Yelizaveta V., Ilya A. Novikov, Anastasiya I. Bilyk, Alexander K. Sharmagiy, Yuri V. Plugatar, and Volodymyr V. Oberemok. 2023. "Perfect Complementarity Mechanism for Aphid Control: Oligonucleotide Insecticide Macsan-11 Selectively Causes High Mortality Rate for Macrosiphoniella sanborni Gillette" International Journal of Molecular Sciences 24, no. 14: 11690. https://doi.org/10.3390/ijms241411690