
The Extracellular Vesicle Citrullinome and Signature in a Piglet Model of Neonatal Seizures
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
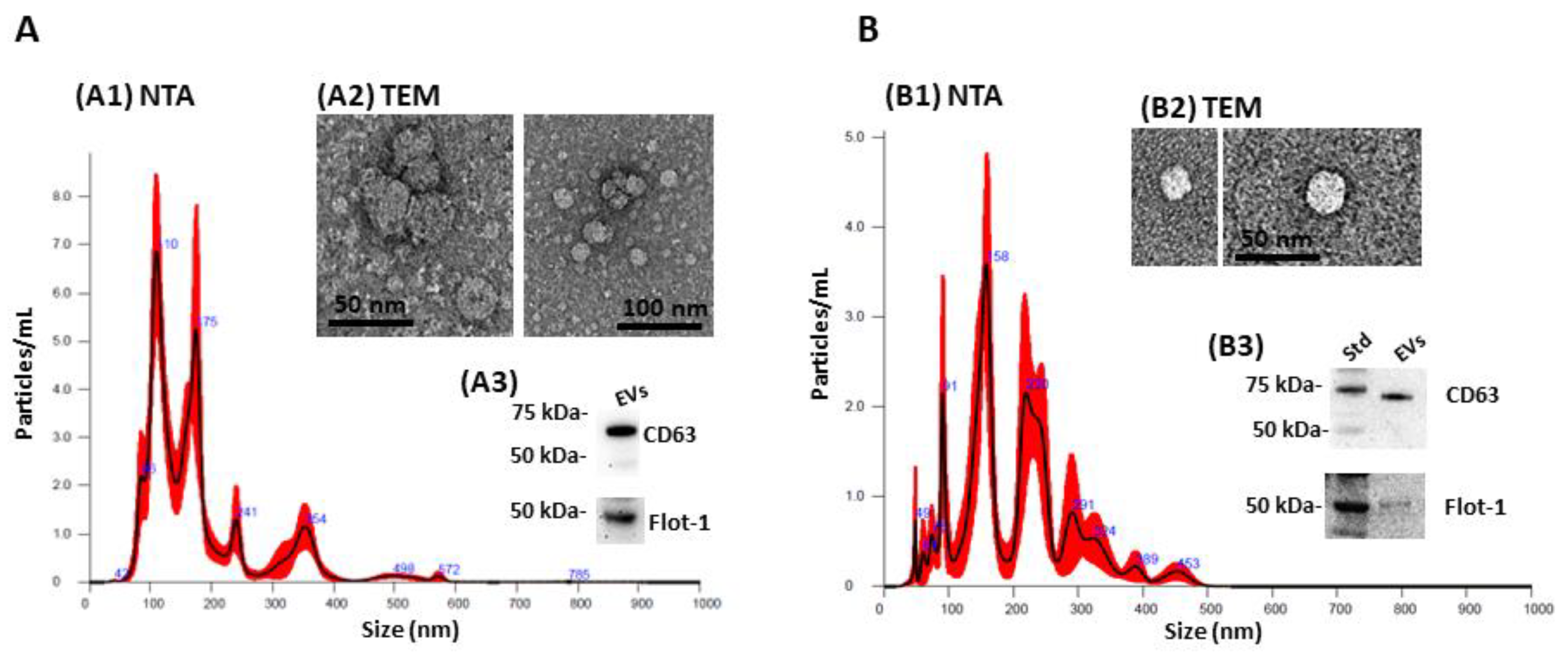
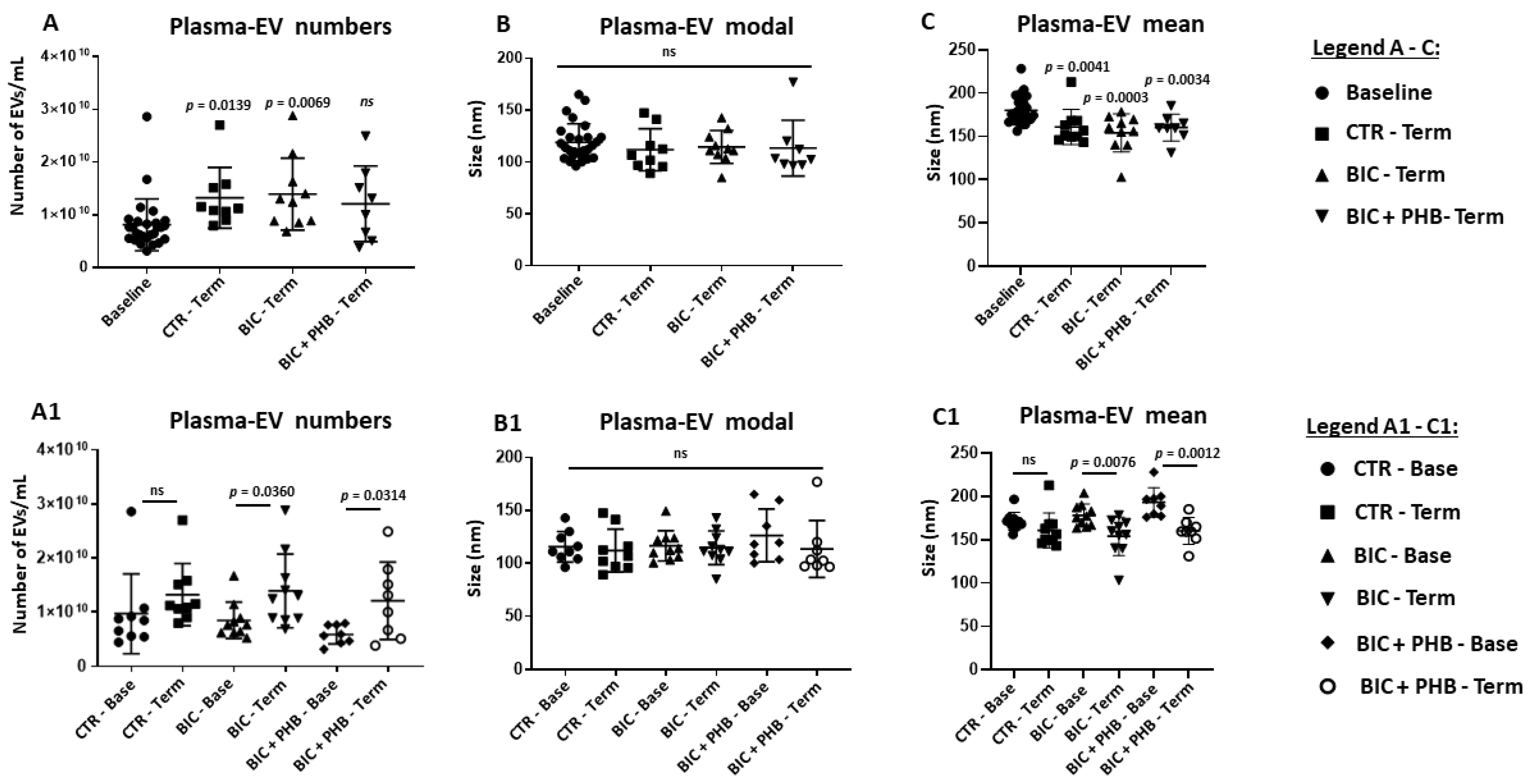
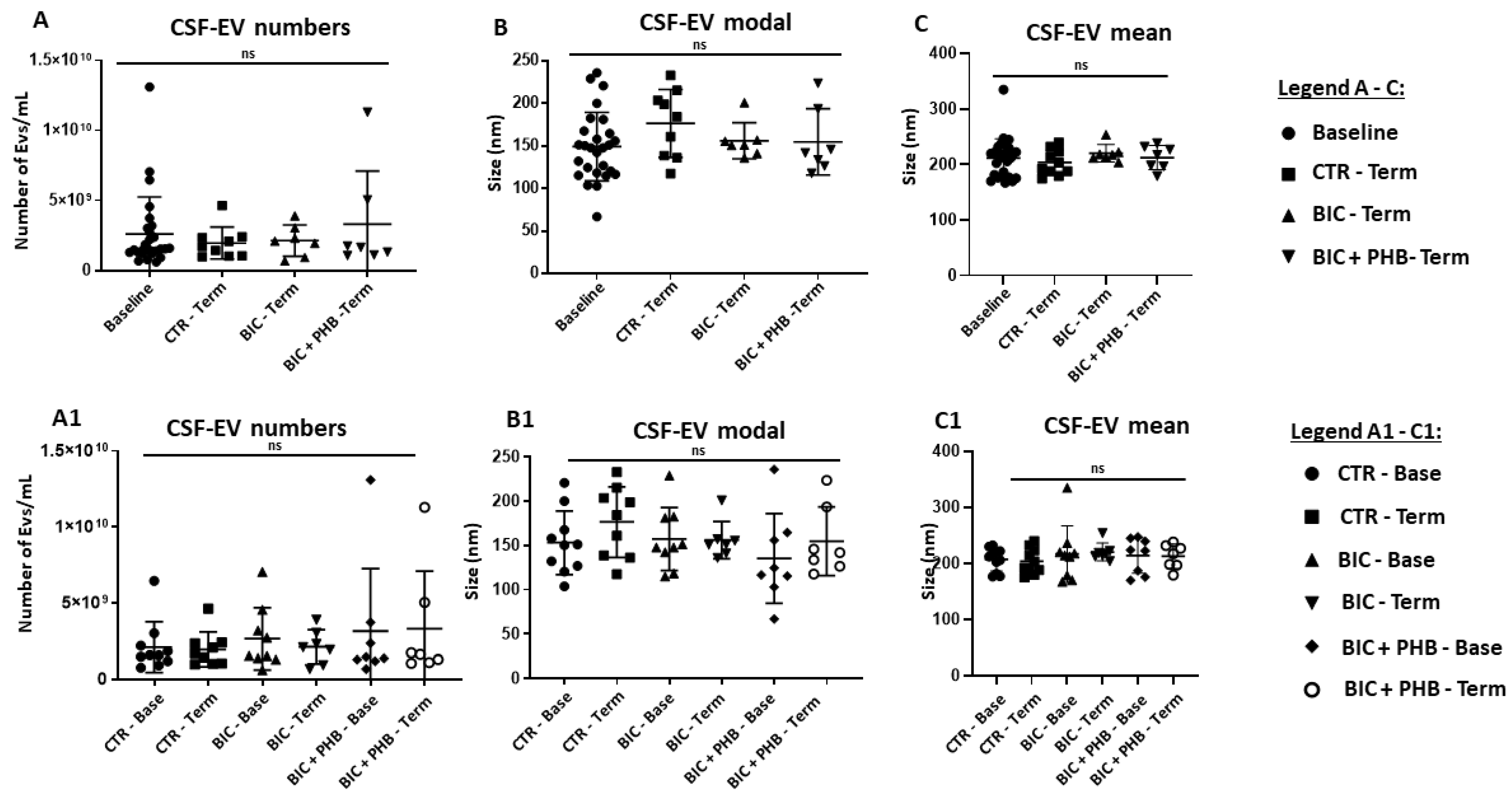
2.1. Extracellular Vesicle (EV) Characterisation and EV Profiling from Piglet Plasma and CSF
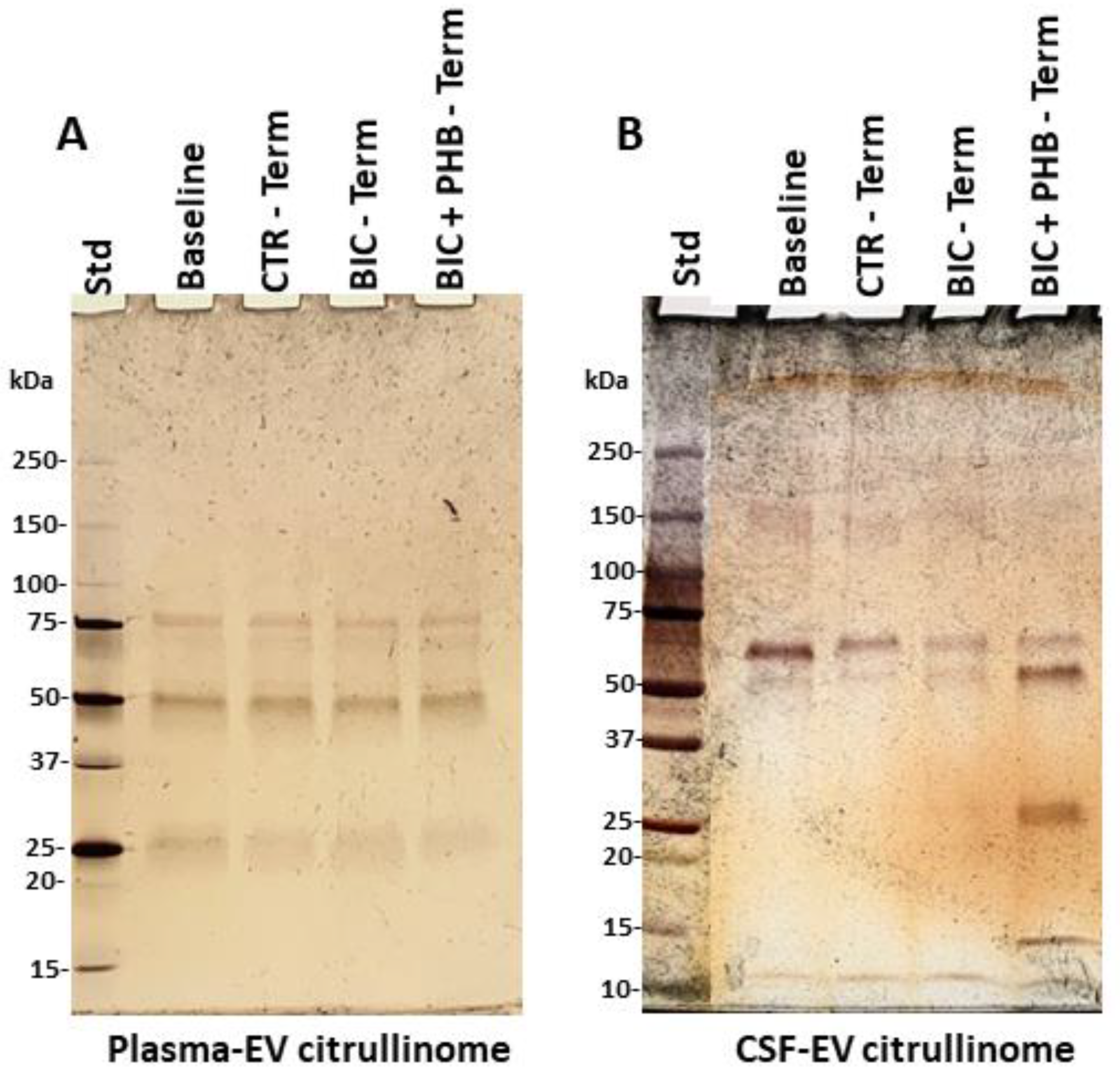
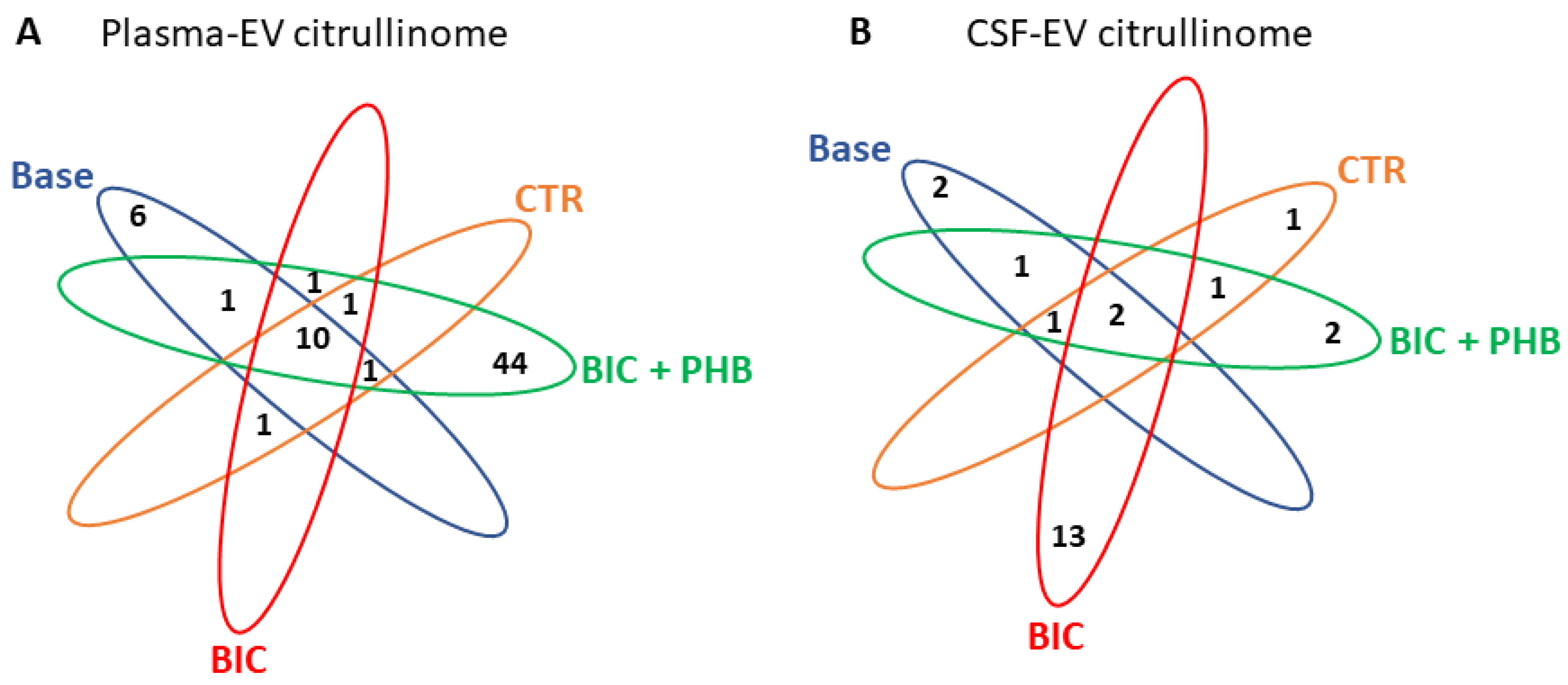
2.2. The Citrullinome of Plasma-EVs and CSF-EVs in a Piglet Model of Neonatal Seizures
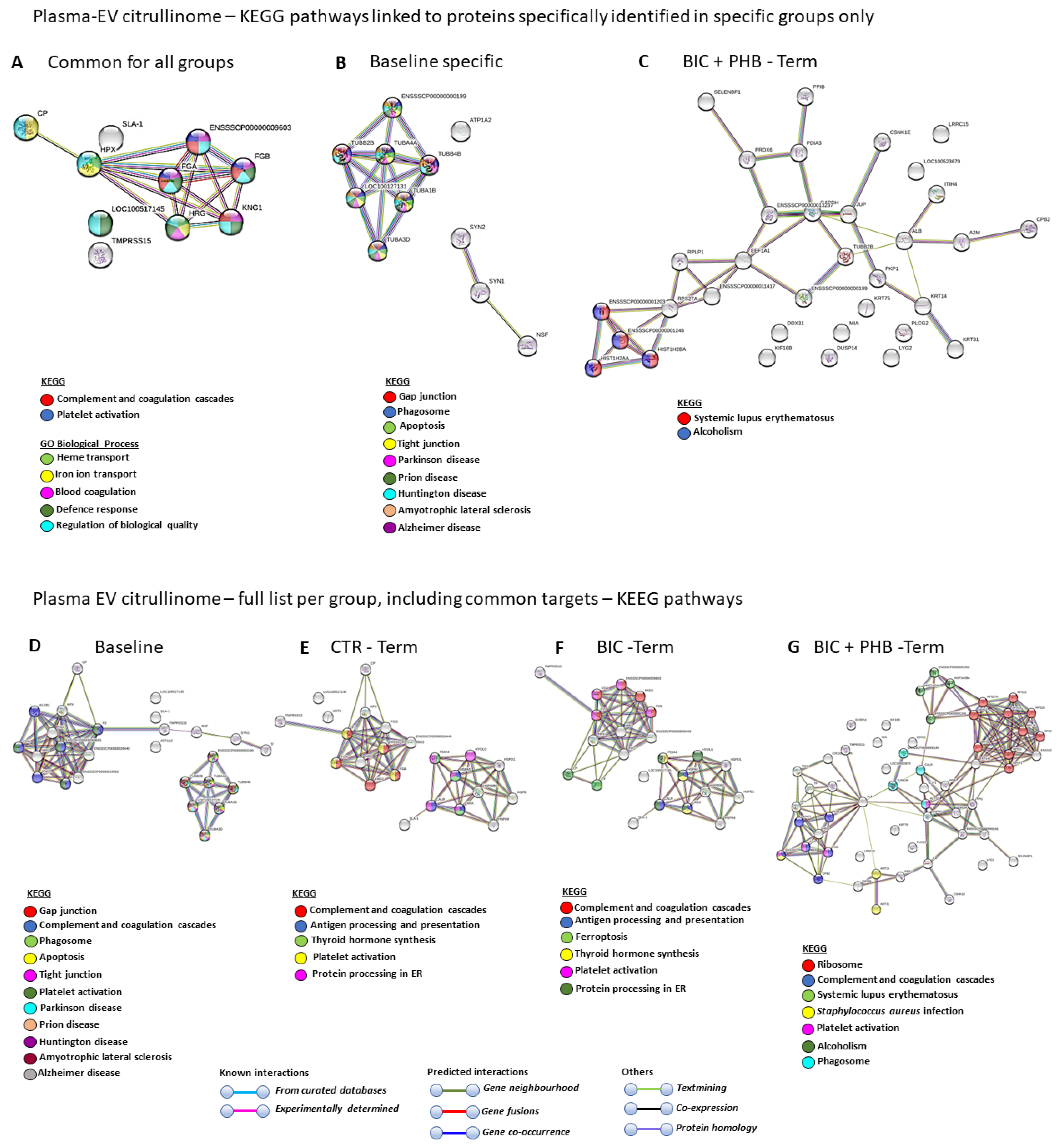
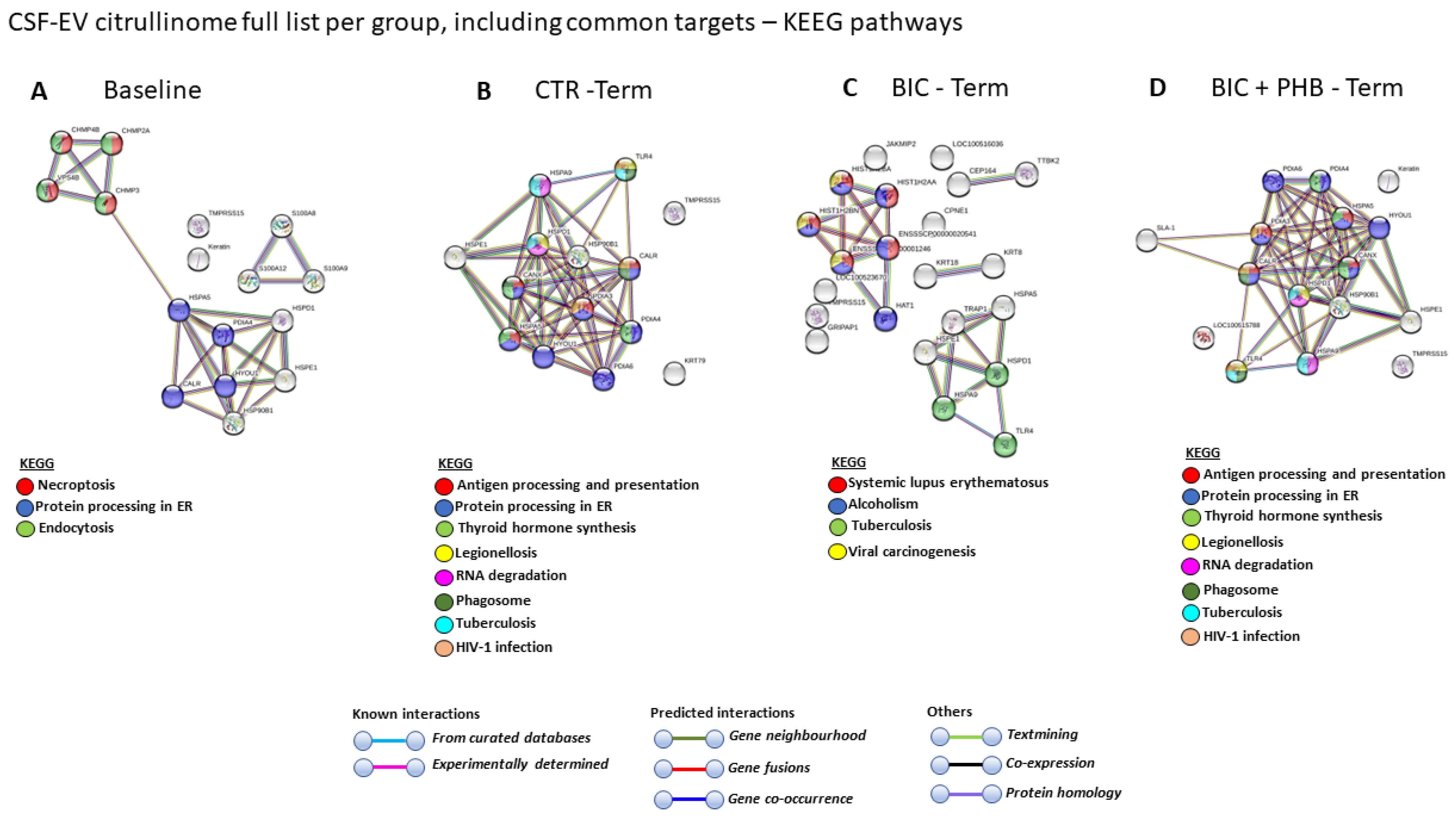
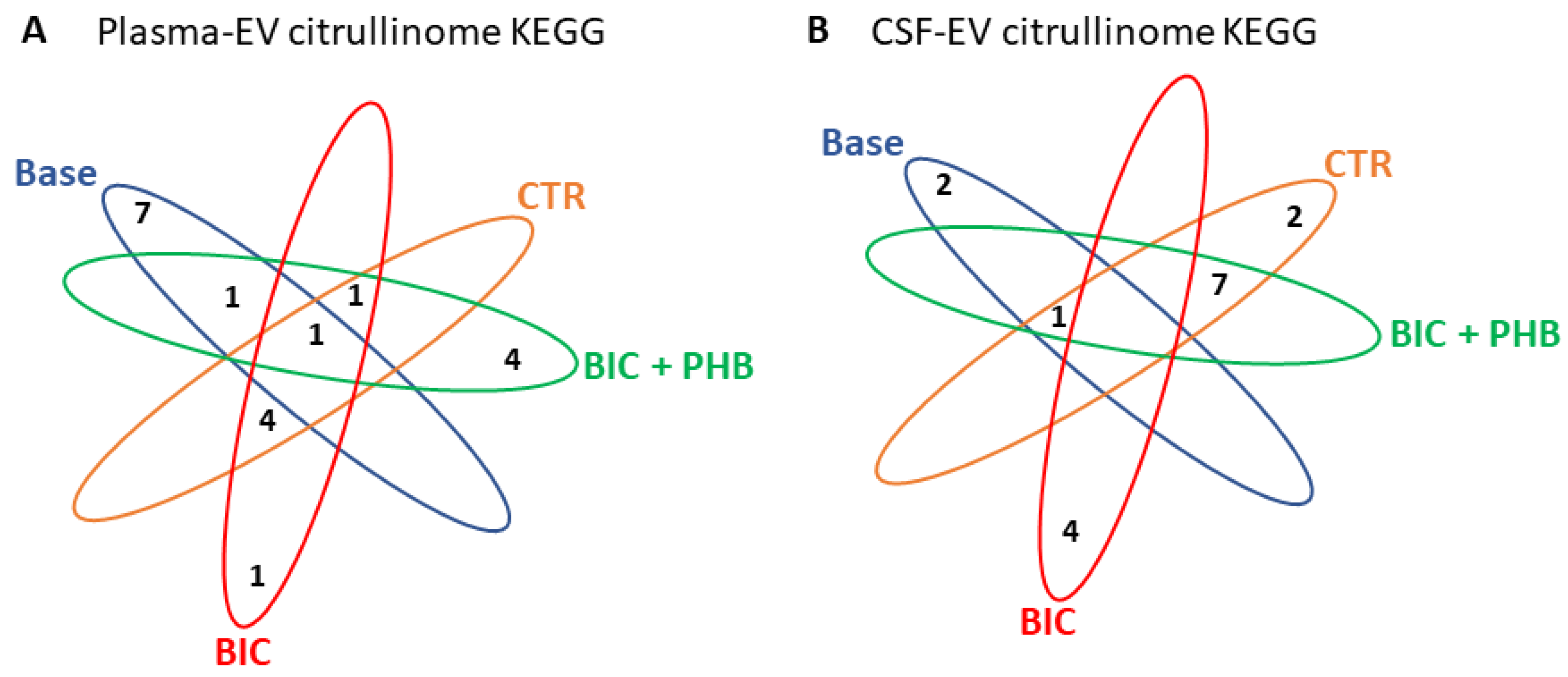
2.3. Protein Interaction Network Analysis of the Plasma-EV and CSF-EV Citrullinomes
2.3.1. Protein Interaction Networks for the Plasma-EV Citrullinome
2.3.2. Protein Interaction Networks of the CSF-EV Citrullinome
3. Discussion
4. Materials and Methods
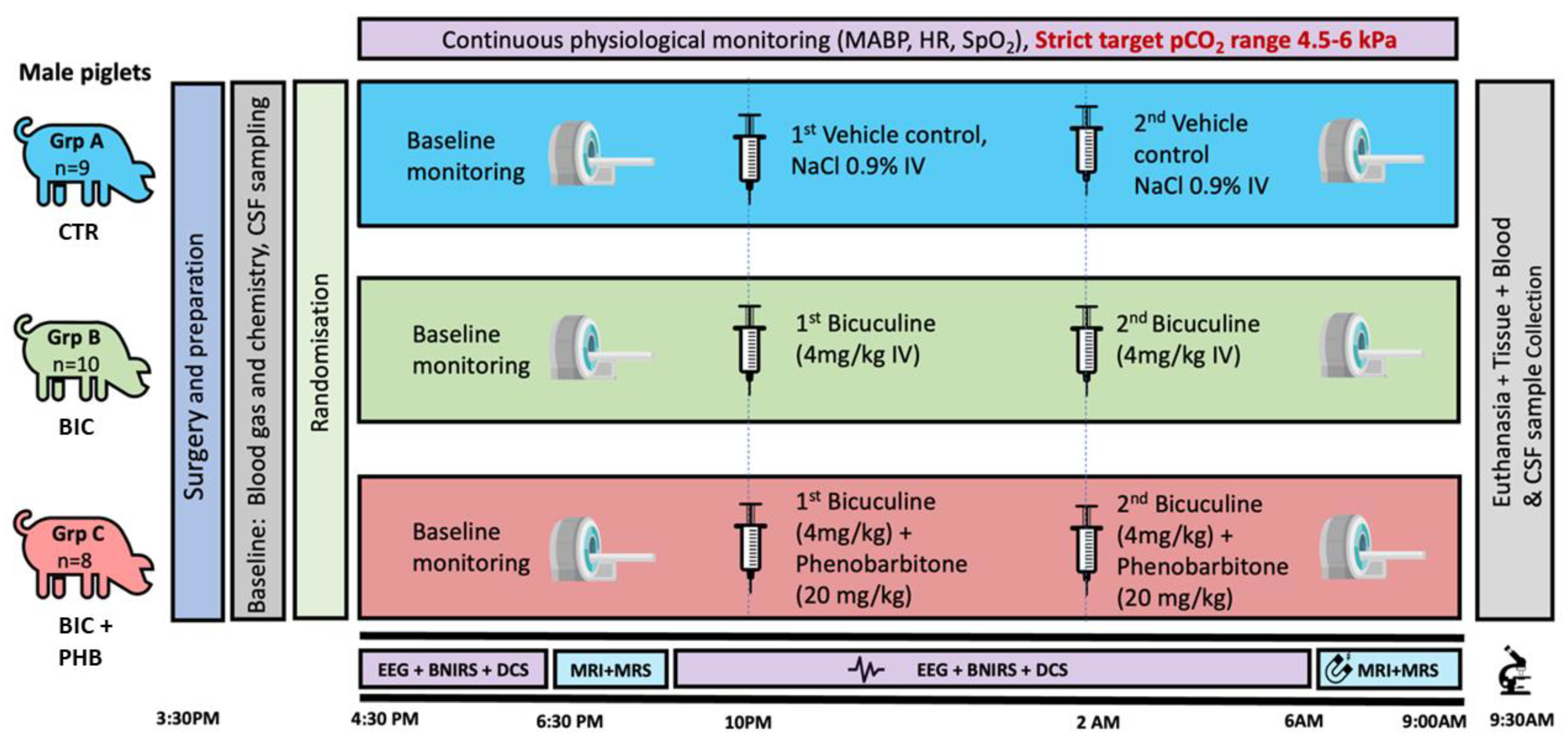
4.1. Piglet Model
4.2. Plasma and CSF Isolation
4.3. Extracellular Vesicle Isolation and Characterisation
4.4. Nanoparticle Tracking Analysis (NTA)
4.5. Western Blotting
4.6. Transmission Electron Microscopy
4.7. Proteomic Analysis of Citrullinated EV Protein-Cargo
4.8. Protein Interaction Network Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clozel, M.; Daval, J.L.; Monin, P.; Dubruc, C.; Morselli, P.L.; Vert, P. Regional cerebral blood flow during bicuculline-induced seizures in the newborn piglet: Effect of phenobarbital. Dev. Pharmacol. Ther. 1985, 8, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Johnston, G.A. Advantages of an antagonist: Bicuculline and other GABA antagonists. Br. J. Pharmacol. 2013, 169, 328–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harsono, M.; Pourcyrous, M.; Jolly, E.J.; de Jongh Curry, A.; Fedinec, A.L.; Liu, J.; Basuroy, S.; Zhuang, D.; Leffler, C.W.; Parfenova, H. Selective head cooling during neonatal seizures prevents postictal cerebral vascular dysfunction without reducing epileptiform activity. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H1202–H1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parfenova, H.; Daley, M.L.; Carratu, P.; Leffler, C.W. Heme oxygenase inhibition reduces neuronal activation evoked by bicuculline in newborn pigs. Brain Res. 2004, 1014, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Avanzini, G. Epilepsy, Experimental Models. In Encyclopedia of the Neurological Sciences; Aminoff, M.J., Daroff, R.B., Eds.; Academic Press: Cambridge, MA, USA, 2003; pp. 205–209. ISBN 9780122268700. [Google Scholar] [CrossRef]
- Pressler, R.M.; Abend, N.S.; Auvin, S.; Boylan, G.; Brigo, F.; Cilio, M.R.; De Vries, L.S.; Elia, M.; Espache, A.; Hahn, C.D.; et al. Treatment of Seizures in the Neonate: Guidelines and Consensus-Based Recommendations—Special Report from the ILAE Task Force on Neonatal Seizures. International League against Epilepsy. 2022. Available online: https://www.ilae.org/guidelines/guidelines-and-reports/treatment-of-seizures-in-the-neonate-guidelines-and-consensus-based-recommendations (accessed on 5 June 2023).
- Ben-Ari, Y.; Holmes, G.L. Effects of seizures on developmental processes in the immature brain. Lancet Neurol. 2006, 5, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Witalison, E.E.; Thompson, P.R.; Hofseth, L.J. Protein arginine deiminases and associated citrullination: Physiological functions and diseases associated with dysregulation. Curr. Drug Targets 2015, 16, 700–710. [Google Scholar] [CrossRef]
- Bicker, K.L.; Thompson, P.R. The protein arginine deiminases: Structure, function, inhibition, and disease. Biopolymers 2013, 99, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Briot, J.; Simon, M.; Méchin, M.C. Deimination, Intermediate Filaments and Associated Proteins. Int. J. Mol. Sci. 2020, 21, 8746. [Google Scholar] [CrossRef]
- Ciesielski, O.; Biesiekierska, M.; Panthu, B.; Soszyński, M.; Pirola, L.; Balcerczyk, A. Citrullination in the pathology of inflammatory and autoimmune disorders: Recent advances and future perspectives. Cell. Mol. Life Sci. 2022, 79, 94. [Google Scholar] [CrossRef]
- Yu, K.; Proost, P. Insights into peptidylarginine deiminase expression and citrullination pathways. Trends Cell Biol. 2022, 32, 746–761. [Google Scholar] [CrossRef]
- Lange, S.; Rocha-Ferreira, E.; Thei, L.; Mawjee, P.; Bennett, K.; Thompson, P.R.; Subramanian, V.; Nicholas, A.P.; Peebles, D.; Hristova, M.; et al. Peptidylarginine deiminases: Novel drug targets for prevention of neuronal damage following hypoxic ischemic insult (HI) in neonates. J. Neurochem. 2014, 130, 555–562. [Google Scholar] [CrossRef] [Green Version]
- Lange, S. Peptidylarginine Deiminases as Drug Targets in Neonatal Hypoxic-Ischemic Encephalopathy. Front. Neurol. 2016, 7, 22. [Google Scholar] [CrossRef] [Green Version]
- Lange, S.; Gögel, S.; Leung, K.Y.; Vernay, B.; Nicholas, A.P.; Causey, C.P.; Thompson, P.R.; Greene, N.D.; Ferretti, P. Protein deiminases: New players in the developmentally regulated loss of neural regenerative ability. Dev. Biol. 2011, 355, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Lazarus, R.C.; Buonora, J.E.; Flora, M.N.; Freedy, J.G.; Holstein, G.R.; Martinelli, G.P.; Jacobowitz, D.M.; Mueller, G.P. Protein Citrullination: A Proposed Mechanism for Pathology in Traumatic Brain Injury. Front. Neurol. 2015, 6, 204. [Google Scholar] [CrossRef] [Green Version]
- Attilio, P.J.; Flora, M.; Kamnaksh, A.; Bradshaw, D.J.; Agoston, D.; Mueller, G.P. The Effects of Blast Exposure on Protein Deimination in the Brain. Oxid. Med. Cell. Longev. 2017, 2017, 8398072. [Google Scholar] [CrossRef] [Green Version]
- Kosgodage, U.S.; Uysal-Onganer, P.; MacLatchy, A.; Kraev, I.; Chatterton, N.P.; Nicholas, A.P.; Inal, J.M.; Lange, S. Peptidylarginine Deiminases Post-Translationally Deiminate Prohibitin and Modulate Extracellular Vesicle Release and MicroRNAs in Glioblastoma Multiforme. Int. J. Mol. Sci. 2018, 20, 103. [Google Scholar] [CrossRef] [Green Version]
- Uysal-Onganer, P.; MacLatchy, A.; Mahmoud, R.; Kraev, I.; Thompson, P.R.; Inal, J.M.; Lange, S. Peptidylarginine Deiminase Isozyme-Specific PAD2, PAD3 and PAD4 Inhibitors Differentially Modulate Extracellular Vesicle Signatures and Cell Invasion in Two Glioblastoma Multiforme Cell Lines. Int. J. Mol. Sci. 2020, 21, 1495. [Google Scholar] [CrossRef] [Green Version]
- Nicholas, A.P. Dual immunofluorescence study of citrullinated proteins in Parkinson diseased substantia nigra. Neurosci. Lett. 2011, 495, 26–29. [Google Scholar] [CrossRef]
- Sancandi, M.; Uysal-Onganer, P.; Kraev, I.; Mercer, A.; Lange, S. Protein Deimination Signatures in Plasma and Plasma-EVs and Protein Deimination in the Brain Vasculature in a Rat Model of Pre-Motor Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 2743. [Google Scholar] [CrossRef] [Green Version]
- Petrozziello, T.; Mills, A.N.; Vaine, C.A.; Penney, E.B.; Fernandez-Cerado, C.; Legarda, G.; Velasco-Andrada, M.S.; Acuña, P.J.; Ang, M.A.; Muñoz, E.L.; et al. Neuroinflammation and histone H3 citrullination are increased in X-linked Dystonia Parkinsonism post-mortem prefrontal cortex. Neurobiol. Dis. 2020, 144, 105032. [Google Scholar] [CrossRef]
- Mercer, A.; Jaunmuktane, Z.; Hristova, M.; Lange, S. Differential, Stage Dependent Detection of Peptidylarginine Deiminases and Protein Deimination in Lewy Body Diseases-Findings from a Pilot Study. Int. J. Mol. Sci. 2022, 23, 13117. [Google Scholar] [CrossRef] [PubMed]
- Ishigami, A.; Ohsawa, T.; Hiratsuka, M.; Taguchi, H.; Kobayashi, S.; Saito, Y.; Murayama, S.; Asaga, H.; Toda, T.; Kimura, N.; et al. Abnormal accumulation of citrullinated proteins catalyzed by peptidylarginine deiminase in hippocampal extracts from patients with Alzheimer’s disease. J. Neurosci. Res. 2005, 80, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, A.P. Dual immunofluorescence study of citrullinated proteins in Alzheimer diseased frontal cortex. Neurosci. Lett. 2013, 545, 107–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishigami, A.; Masutomi, H.; Handa, S.; Nakamura, M.; Nakaya, S.; Uchida, Y.; Saito, Y.; Murayama, S.; Jang, B.; Jeon, Y.C.; et al. Mass spectrometric identification of citrullination sites and immunohistochemical detection of citrullinated glial fibrillary acidic protein in Alzheimer’s disease brains. J. Neurosci. Res. 2015, 93, 1664–1674. [Google Scholar] [CrossRef]
- Wang, L.; Chen, H.; Tang, J.; Guo, Z.; Wang, Y. Peptidylarginine Deiminase and Alzheimer’s Disease. J. Alzheimer’s Dis. 2022, 85, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Jang, B.; Ishigami, A.; Maruyama, N.; Carp, R.I.; Kim, Y.S.; Choi, E.K. Peptidylarginine deiminase and protein citrullination in prion diseases: Strong evidence of neurodegeneration. Prion 2013, 7, 42–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yusuf, I.O.; Qiao, T.; Parsi, S.; Tilvawala, R.; Thompson, P.R.; Xu, Z. Protein citrullination marks myelin protein aggregation and disease progression in mouse ALS models. Acta Neuropathol. Commun. 2022, 10, 135. [Google Scholar] [CrossRef]
- Raijmakers, R.; Vogelzangs, J.; Croxford, J.L.; Wesseling, P.; van Venrooij, W.J.; Pruijn, G.J. Citrullination of central nervous system proteins during the development of experimental autoimmune encephalomyelitis. J. Comp. Neurol. 2005, 486, 243–253. [Google Scholar] [CrossRef]
- Mastronardi, F.G.; Wood, D.D.; Mei, J.; Raijmakers, R.; Tseveleki, V.; Dosch, H.M.; Probert, L.; Casaccia-Bonnefil, P.; Moscarello, M.A. Increased citrullination of histone H3 in multiple sclerosis brain and animal models of demyelination: A role for tumor necrosis factor-induced peptidylarginine deiminase 4 translocation. J. Neurosci. 2006, 26, 11387–11396. [Google Scholar] [CrossRef] [Green Version]
- Faigle, W.; Cruciani, C.; Wolski, W.; Roschitzki, B.; Puthenparampil, M.; Tomas-Ojer, P.; Sellés-Moreno, C.; Zeis, T.; Jelcic, I.; Schaeren-Wiemers, N.; et al. Brain Citrullination Patterns and T Cell Reactivity of Cerebrospinal Fluid-Derived CD4+ T Cells in Multiple Sclerosis. Front. Immunol. 2019, 10, 540. [Google Scholar] [CrossRef] [Green Version]
- Kholia, S.; Jorfi, S.; Thompson, P.R.; Causey, C.P.; Nicholas, A.P.; Inal, J.M.; Lange, S. A novel role for peptidylarginine deiminases in microvesicle release reveals therapeutic potential of PAD inhibition in sensitizing prostate cancer cells to chemotherapy. J. Extracell. Vesicles 2015, 4, 26192. [Google Scholar] [CrossRef] [Green Version]
- Kosgodage, U.S.; Trindade, R.P.; Thompson, P.R.; Inal, J.M.; Lange, S. Chloramidine/Bisindolylmaleimide-I-Mediated Inhibition of Exosome and Microvesicle Release and Enhanced Efficacy of Cancer Chemotherapy. Int. J. Mol. Sci. 2017, 18, 1007. [Google Scholar] [CrossRef] [Green Version]
- Lange, S. Peptidylarginine deiminases and extracellular vesicles: Prospective drug targets and biomarkers in central nervous system diseases and repair. Neural Regen. Res. 2021, 16, 934–938. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Lim, Y.J.; Lee, S.J. Are exosomes the vehicle for protein aggregate propagation in neurodegenerative diseases? Acta Neuropathol. Commun. 2017, 5, 64. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Bai, X.; Zhang, A.; Huang, J.; Xu, S.; Zhang, J. Role of Exosomes in Central Nervous System Diseases. Front. Mol. Neurosci. 2019, 12, 240. [Google Scholar] [CrossRef]
- Hill, A.F. Extracellular Vesicles and Neurodegenerative Diseases. J. Neurosci. 2019, 39, 9269–9273. [Google Scholar] [CrossRef] [PubMed]
- Costine-Bartell, B.A.; McGuone, D.; Price, G.; Crawford, E.; Keeley, K.L.; Munoz-Pareja, J.; Dodge, C.P.; Staley, K.; Duhaime, A.C. Development of a Model of Hemispheric Hypodensity (“Big Black Brain”). J. Neurotrauma 2019, 36, 815–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terndrup, T.E.; Paskanik, A.M.; Fordyce, W.E.; Kanter, R.K. Development of a piglet model of status epilepticus: Preliminary results. Ann. Emerg. Med. 1993, 22, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Picca, A.; Guerra, F.; Calvani, R.; Marini, F.; Biancolillo, A.; Landi, G.; Beli, R.; Landi, F.; Bernabei, R.; Bentivoglio, A.R.; et al. Mitochondrial Signatures in Circulating Extracellular Vesicles of Older Adults with Parkinson’s Disease: Results from the EXosomes in PArkiNson’s Disease (EXPAND) Study. J. Clin. Med. 2020, 9, 504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, S.; Gallagher, M.; Kholia, S.; Kosgodage, U.S.; Hristova, M.; Hardy, J.; Inal, J.M. Peptidylarginine Deiminases-Roles in Cancer and Neurodegeneration and Possible Avenues for Therapeutic Intervention via Modulation of Exosome and Microvesicle (EMV) Release? Int. J. Mol. Sci. 2017, 18, 1196. [Google Scholar] [CrossRef] [Green Version]
- Lange, S.; Wray, S.; Devine, M.; Matarin, M.; Hardy, J. Protein deimination in protein misfolding disorders–modelled in human induced pluripotent stem cells (iPSCs). In Protein Deimination in Human Health and Disease; Nicholas, A.P., Bhattacharya, S.K., Eds.; Springer Science and Business Media: New York, NY, USA, 2017. [Google Scholar]
- Vaibhav, K.; Braun, M.; Alverson, K.; Khodadadi, H.; Kutiyanawalla, A.; Ward, A.; Banerjee, C.; Sparks, T.; Malik, A.; Rashid, M.H.; et al. Neutrophil extracellular traps exacerbate neurological deficits after traumatic brain injury. Sci. Adv. 2020, 6, eaax8847. [Google Scholar] [CrossRef] [PubMed]
- Fando, J.L.; Conn, M.; Wasterlain, C.G. Brain protein synthesis during neonatal seizures: An experimental study. Exp. Neurol. 1979, 63, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Shimada, N.; Handa, S.; Uchida, Y.; Fukuda, M.; Maruyama, N.; Asaga, H.; Choi, E.K.; Lee, J.; Ishigami, A. Developmental and age-related changes of peptidylarginine deiminase 2 in the mouse brain. J. Neurosci. Res. 2010, 88, 798–806. [Google Scholar] [CrossRef] [PubMed]
- DeBruin, L.S.; Haines, J.D.; Wellhauser, L.A.; Radeva, G.; Schonmann, V.; Bienzle, D.; Harauz, G. Developmental partitioning of myelin basic protein into membrane microdomains. J. Neurosci. Res. 2005, 80, 211–225. [Google Scholar] [CrossRef]
- Yurttas, P.; Vitale, A.M.; Fitzhenry, R.J.; Cohen-Gould, L.; Wu, W.; Gossen, J.A.; Coonrod, S.A. Role for PADI6 and the cytoplasmic lattices in ribosomal storage in oocytes and translational control in the early mouse embryo. Development 2008, 135, 2627–2636. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.C.; Shyur, S.D.; Huang, L.H.; Wu, J.Y.; Ma, Y.C. Focal seizures as an unusual presentation of neonatal lupus erythematosus. Asian Pac. J. Allergy Immunol. 2005, 23, 61–64. [Google Scholar]
- Chan, P.C.; Yu, C.H.; Yeh, K.W.; Horng, J.T.; Huang, J.L. Comorbidities of pediatric systemic lupus erythematosus: A 6-year nationwide population-based study. J. Microbiol. Immunol. Infect. 2016, 49, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Moudgil, A.; Kishore, K.; Srivastava, R.N. Neonatal lupus erythematosus, late onset hypocalcaemia, and recurrent seizures. Arch. Dis. Child. 1987, 62, 736–739. [Google Scholar] [CrossRef] [Green Version]
- Döring, M.; Rohrer, K.M.; Tsiflikas, I.; Buchenau, W.; Wilke, M.; Handgretinger, R.; Poets, C.F.; Goelz, R. A newborn with grouped facial skin lesions and subsequent seizures. BMC Pediatr. 2014, 14, 126. [Google Scholar] [CrossRef]
- Liu, Y.; Lightfoot, Y.L.; Seto, N.; Carmona-Rivera, C.; Moore, E.; Goel, R.; O’neil, L.; Mistry, P.; Hoffmann, V.; Mondal, S.; et al. Peptidylarginine deiminases 2 and 4 modulate innate and adaptive immune responses in TLR-7-dependent lupus. JCI Insight 2018, 3, e124729. [Google Scholar] [CrossRef] [Green Version]
- Colasanti, T.; Spinelli, F.R.; Barbati, C.; Ceccarelli, F.; Scarpa, S.; Vomero, M.; Alessandri, C.; Valesini, G.; Conti, F. Belimumab Decreases Autophagy and Citrullination in Peripheral Blood Mononuclear Cells from Patients with Systemic Lupus Erythematosus. Cells 2022, 11, 262. [Google Scholar] [CrossRef]
- Kosgodage, U.S.; Matewele, P.; Mastroianni, G.; Kraev, I.; Brotherton, D.; Awamaria, B.; Nicholas, A.P.; Lange, S.; Inal, J.M. Peptidylarginine Deiminase Inhibitors Reduce Bacterial Membrane Vesicle Release and Sensitize Bacteria to Antibiotic Treatment. Front. Cell. Infect. Microbiol. 2019, 9, 227. [Google Scholar] [CrossRef] [Green Version]
- Al Adwani, S.; Padhi, A.; Karadottir, H.; Mörman, C.; Gräslund, A.; Végvári, Á.; Johansson, J.; Rising, A.; Agerberth, B.; Bergman, P. Citrullination Alters the Antibacterial and Anti-Inflammatory Functions of the Host Defense Peptide Canine Cathelicidin K9CATH In Vitro. J. Immunol. 2021, 207, 974–984. [Google Scholar] [CrossRef]
- Lima, I.; Santiago, M. Antibodies against cyclic citrullinated peptides in infectious diseases—A systematic review. Clin. Rheumatol. 2010, 29, 1345–1351. [Google Scholar] [CrossRef]
- D’Alessio, S.; Cheng, H.; Eaton, L.; Kraev, I.; Pamenter, M.E.; Lange, S. Acute Hypoxia Alters Extracellular Vesicle Signatures and the Brain Citrullinome of Naked Mole-Rats (Heterocephalus glaber). Int. J. Mol. Sci. 2022, 23, 4683. [Google Scholar] [CrossRef]
- Asindi, A.A.; Antia-Obong, O.E.; Ibia, E.O.; Udo, J.J. Neonatal seizures in Nigerian infants. Afr. J. Med. Med. Sci. 1995, 24, 243–248. [Google Scholar]
- Airede, K.I.; Adeyemi, O.; Ibrahim, T. Neonatal bacterial meningitis and dexamethasone adjunctive usage in Nigeria. Niger. J. Clin. Pract. 2008, 11, 235–245. [Google Scholar]
- Sabouni, F.; Ranjbari, R.; Pourakbari, B.; Mahmoudi, S.; Teymuri, M.; Ashtiani, M.T.; Movahedi, Z.; Mamishi, S. Staphylococcus aureus infections in children in an Iranian referral pediatric hospital. J. Prev. Med. Hyg. 2013, 54, 205–207. [Google Scholar]
- Almuneef, M.; Memish, Z.; Khan, Y.; Kagallwala, A.; Alshaalan, M. Childhood bacterial meningitis in Saudi Arabia. J. Infect. 1998, 36, 157–160. [Google Scholar] [CrossRef]
- Gupta, S.; Vachhrajani, S.; Kulkarni, A.V.; Taylor, M.D.; Dirks, P.; Drake, J.M.; Rutka, J.T. Neurosurgical management of extraaxial central nervous system infections in children. J. Neurosurg. Pediatr. 2011, 7, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Akpede, G.O.; Abiodun, P.O.; Sykes, R.M. Pattern of infections in children under-six years old presenting with convulsions associated with fever of acute onset in a children’s emergency room in Benin City, Nigeria. J. Trop. Pediatr. 1993, 39, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Mattson, R.H. Selection of drugs for the treatment of epilepsy. Semin. Neurol. 1990, 10, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Mattson, R.H. Efficacy and adverse effects of established and new antiepileptic drugs. Epilepsia 1995, 36 (Suppl. S2), S13–S26. [Google Scholar] [CrossRef]
- Tychkivska, O.; Go, C.; Korzhynskyy, Y.; Ostalska, O. Asymmetric neonatal spasms as an early sign of brain malformation potentially caused by regular light alcohol consumption during the first 22 weeks of pregnancy, a clinical case report. Dev. Period Med. 2019, 23, 15–20. [Google Scholar]
- Cho, S.J.; Newton, J.; Li, T.; Khandai, P.; Luta, G.; Lovinger, D.M.; N’Gouemo, P. Prenatal alcohol exposure in the second trimester-equivalent increases the seizure susceptibility in developing rats. Alcohol 2020, 85, 153–164. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, P.; Zhai, B.; Zhang, M.; Xiang, Y.; Fang, J.; Xu, S.; Gao, Y.; Chen, X.; Sui, X.; et al. The emerging role of ferroptosis in inflammation. Biomed. Pharmacother. 2020, 127, 110108. [Google Scholar] [CrossRef]
- Stockwell, B.R.; Friedmann Angeli, J.P.; Bayir, H.; Bush, A.I.; Conrad, M.; Dixon, S.J.; Fulda, S.; Gascón, S.; Hatzios, S.K.; Kagan, V.E.; et al. Ferroptosis: A Regulated Cell Death Nexus Linking Metabolism, Redox Biology, and Disease. Cell 2017, 171, 273–285. [Google Scholar] [CrossRef] [Green Version]
- Jakaria, M.; Belaidi, A.A.; Bush, A.I.; Ayton, S. Ferroptosis as a mechanism of neurodegeneration in Alzheimer’s disease. J. Neurochem. 2021, 159, 804–825. [Google Scholar] [CrossRef]
- Luoqian, J.; Yang, W.; Ding, X.; Tuo, Q.Z.; Xiang, Z.; Zheng, Z.; Guo, Y.J.; Li, L.; Guan, P.; Ayton, S.; et al. Ferroptosis promotes T-cell activation-induced neurodegeneration in multiple sclerosis. Cell. Mol. Immunol. 2022, 19, 913–924. [Google Scholar] [CrossRef]
- He, Y.; Ying, J.; Tang, J.; Zhou, R.; Qu, H.; Qu, Y.; Mu, D. Neonatal Arterial Ischaemic Stroke: Advances in Pathologic Neural Death, Diagnosis, Treatment, and Prognosis. Curr. Neuropharmacol. 2022, 20, 2248–2266. [Google Scholar]
- Cai, Y.; Yang, Z. Ferroptosis and Its Role in Epilepsy. Front. Cell. Neurosci. 2021, 15, 696889. [Google Scholar] [CrossRef]
- Chen, X.; Kang, R.; Kroemer, G.; Tang, D. Ferroptosis in infection, inflammation, and immunity. J. Exp. Med. 2021, 218, e20210518. [Google Scholar] [CrossRef]
- Lassiter, H.A. The role of complement in neonatal hypoxic-ischemic cerebral injury. Clin. Perinatol. 2004, 31, 117–127. [Google Scholar] [CrossRef]
- Rocha-Ferreira, E.; Hristova, M. Antimicrobial peptides and complement in neonatal hypoxia-ischemia induced brain damage. Front. Immunol. 2015, 6, 56. [Google Scholar] [CrossRef] [Green Version]
- Shah, T.A.; Pallera, H.K.; Kaszowski, C.L.; Bass, W.T.; Lattanzio, F.A. Therapeutic Hypothermia Inhibits the Classical Complement Pathway in a Rat Model of Neonatal Hypoxic-Ischemic Encephalopathy. Front. Neurosci. 2021, 15, 616734. [Google Scholar] [CrossRef]
- Kopczynska, M.; Zelek, W.M.; Vespa, S.; Touchard, S.; Wardle, M.; Loveless, S.; Thomas, R.H.; Hamandi, K.; Morgan, B.P. Complement system biomarkers in epilepsy. Seizure 2018, 60, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Tan, T.H.; Perucca, P.; O’Brien, T.J.; Kwan, P.; Monif, M. Inflammation, ictogenesis, and epileptogenesis: An exploration through human disease. Epilepsia 2021, 62, 303–324. [Google Scholar] [CrossRef]
- Li, G.; Zhang, S.; Cheng, Y.; Lu, Y.; Jia, Z.; Yang, X.; Zhang, S.; Guo, W.; Pei, L. Baicalin suppresses neuron autophagy and apoptosis by regulating astrocyte polarization in pentylenetetrazol-induced epileptic rats and PC12 cells. Brain Res. 2022, 1774, 147723. [Google Scholar] [CrossRef]
- Chen, M.; Edwards, S.R.; Reutens, D.C. Complement in the Development of Post-Traumatic Epilepsy: Prospects for Drug Repurposing. J. Neurotrauma 2020, 37, 692–705. [Google Scholar] [CrossRef]
- Theron, S.; Andronikou, S.; Grobbelaar, M.; Steyn, F.; Mapukata, A.; du Plessis, J. Localized basal meningeal enhancement in tuberculous meningitis. Pediatr. Radiol. 2006, 36, 1182–1185. [Google Scholar] [CrossRef] [PubMed]
- Scher, M.S. Neonatal seizure classification: A fetal perspective concerning childhood epilepsy. Epilepsy Res. 2006, 70 (Suppl. S1), S41–S57. [Google Scholar] [CrossRef] [PubMed]
- Scher, M.S. “The First Thousand Days” Define a Fetal/Neonatal Neurology Program. Front. Pediatr. 2021, 9, 683138. [Google Scholar] [CrossRef] [PubMed]
- Netto, C.A.; Sanches, E.; Odorcyk, F.K.; Duran-Carabali, L.E.; Weis, S.N. Sex-dependent consequences of neonatal brain hypoxia-ischemia in the rat. J. Neurosci. Res. 2017, 95, 409–421. [Google Scholar] [CrossRef] [Green Version]
- Penny, T.R.; Pham, Y.; Sutherland, A.E.; Lee, J.; Jenkin, G.; Fahey, M.C.; Miller, S.L.; McDonald, C.A. Umbilical cord blood therapy modulates neonatal hypoxic ischemic brain injury in both females and males. Sci. Rep. 2021, 11, 15788. [Google Scholar] [CrossRef]
- Lecuyer, M.; Pathipati, P.; Faustino, J.; Vexler, Z.S. Neonatal stroke enhances interaction of microglia-derived extracellular vesicles with microglial cells. Neurobiol. Dis. 2021, 157, 105431. [Google Scholar] [CrossRef]
- Goetzl, L.; Merabova, N.; Darbinian, N.; Martirosyan, D.; Poletto, E.; Fugarolas, K.; Menkiti, O. Diagnostic Potential of Neural Exosome Cargo as Biomarkers for Acute Brain Injury. Ann. Clin. Transl. Neurol. 2017, 5, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Pineles, B.; Mani, A.; Sura, L.; Rossignol, C.; Albayram, M.; Weiss, M.D.; Goetzl, L. Neuronal exosome proteins: Novel biomarkers for predicting neonatal response to therapeutic hypothermia. Arch. Dis. Child. Fetal Neonatal Ed. 2022, 107, 60–64. [Google Scholar] [CrossRef]
- Kyriakidou, Y.; Cooper, I.; Kraev, I.; Lange, S.; Elliott, B.T. Preliminary Investigations Into the Effect of Exercise-Induced Muscle Damage on Systemic Extracellular Vesicle Release in Trained Younger and Older Men. Front. Physiol. 2021, 12, 723931. [Google Scholar] [CrossRef]
- Nicholas, A.P.; Whitaker, J.N. Preparation of a monoclonal antibody to citrullinated epitopes: Its characterization and some applications to immunohistochemistry in human brain. Glia 2002, 37, 328–336. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Hits | Baseline | CTR Terminal | BIC Terminal | BIC + PHB Terminal |
|---|---|---|---|---|
| Fibrinogen alpha chain | V | V | V | V |
| Fibrinogen beta chain | V | V | V | V |
| Fibrinogen gamma chain | V | V | V | V |
| Trypsinogen isoform X1 | V | V | V | V |
| Ig-like domain-containing protein | V | V | V | V |
| Histidine-rich glycoprotein | V | V | V | V |
| Hemopexin | V | V | V | V |
| Kininogen 1 | V | V | V | V |
| Complement C3 | V | V | V | V |
| Ceruloplasmin | V | V | V | V |
| Synapsin-1 | V | |||
| Sodium/potassium-transporting ATPase subunit alpha | V | |||
| Tubulin, beta 2B class Iib | V | |||
| Tubulin beta 4B class Ivb | V | |||
| Vesicle-fusing ATPase | V | |||
| Synapsin II | V | |||
| Keratin, type II cuticular Hb5 | V | |||
| IF rod domain-containing protein | V | |||
| Keratin 14 | V | |||
| Keratin 31 | V | |||
| Keratin 33A | V | |||
| Keratin 34 | V | |||
| Keratin 40 | V | |||
| Keratin 75 | V | |||
| SH3 domain-containing protein | V | |||
| Selenium binding protein 1 | V | |||
| Histone domain-containing protein | V | |||
| Histone H2A | V | |||
| Histone H2B | V | |||
| Histone H4 | V | |||
| Tubulin beta chain | V | |||
| 14_3_3 domain-containing protein | V | |||
| Leucine rich repeat containing 15 | V | |||
| Peptidyl-prolyl cis-trans isomerase | V | |||
| ADP/ATP translocase | V | |||
| Albumin | V | |||
| Plakophilin 1 | V | |||
| Tubulin alpha chain | V | |||
| Inter-alpha-trypsin inhibitor heavy chain H4 | V | |||
| Alpha-2-macroglobulin isoform X1 | V | |||
| Junction plakoglobin | V | |||
| Glyceraldehyde-3-phosphate dehydrogenase | V | |||
| Protein kinase domain-containing protein | V | |||
| Elongation factor 1-alpha | V | |||
| 40S ribosomal protein S16 | V | |||
| Phosphoinositide phospholipase C | V | |||
| Urb2 domain-containing protein | V | |||
| 60S acidic ribosomal protein P1 | V | |||
| Kinesin family member 16B | V | |||
| Phosphoglycerate kinase | V | |||
| Carboxypeptidase B2 | V | |||
| Peroxiredoxin-6 | V | |||
| Heat shock protein family A (Hsp70) member 8 | V | |||
| Lysozyme g-like protein | V | |||
| Protein disulfide-isomerase | V | |||
| Dual specificity phosphatase 14 | V | |||
| RNA helicase | V | |||
| Ubiquitin-40S ribosomal protein S27a | V | |||
| Keratin 5 | V | V | V | |
| 60 kDa heat shock protein, mitochondrial | V | V | V | |
| Actin, gamma 1 | V | V | ||
| Endoplasmin | V | V | ||
| Serotransferrin | V | V |
| Protein Hits | Baseline | CTR Terminal | BIC Terminal | BIC + PHB Terminal |
|---|---|---|---|---|
| Trypsinogen isoform X1 | V | V | V | V |
| 60 kDa heat shock protein, mitochondrial | V | V | V | V |
| EF-hand domain-containing protein | V | |||
| Charged multivesicular body protein 2a | V | |||
| Keratin 79 | V | |||
| Keratin, type II microfibrillar, component 7C | V | |||
| IF rod domain-containing protein | V | |||
| Keratin, type I cuticular Ha3-II | V | |||
| IF rod domain-containing protein | V | |||
| Keratin 18 | V | |||
| Histone H2A | V | |||
| Histone H2B | V | |||
| Histone H4 | V | |||
| Kinesin-like protein KIF16B isoform 2 | V | |||
| GRIP1 associated protein 1 | V | |||
| Janus kinase and microtubule interacting protein 2 | V | |||
| Tau tubulin kinase 2 | V | |||
| C2 domain-containing protein | V | |||
| GLOBIN domain-containing protein | V | |||
| Ig-like domain-containing protein | V | |||
| Keratin 5 | V | V | ||
| Endoplasmin | V | V | V | |
| IgG heavy chain | V | V |
| KEGG PATHWAYS | Baseline | CTR Terminal | BIC Terminal | BIC + PHB Terminal |
|---|---|---|---|---|
| Apoptosis | V | |||
| Tight junction | V | |||
| Parkinson’s disease | V | |||
| Prion disease | V | |||
| Huntington’s disease | V | |||
| Amyotrophic lateral sclerosis | V | |||
| Alzheimer disease | V | |||
| Complement and coagulation cascades | V | V | V | V |
| Gap junction | V | V | ||
| Phagosome | V | V | ||
| Platelet activation | V | V | ||
| Antigen processing and presentation | V | V | ||
| Thyroid hormone synthesis | V | V | ||
| Platelet activation | V | V | V | |
| Protein processing in ER | V | V | ||
| Ferroptosis | V | |||
| Ribosome | V | |||
| Systemic lupus erythematosus | V | |||
| Staphylococcus aureus infection | V | |||
| Alcoholism | V |
| KEGG PATHWAYS | Baseline | CTR Terminal | BIC Terminal | BIC + PHB Terminal |
|---|---|---|---|---|
| Necroptosis | V | |||
| Endocytosis | V | |||
| Protein processing in ER | V | V | V | |
| Antigen processing and presentation | V | V | ||
| Thyroid hormone synthesis | V | V | ||
| Legionellosis | V | V | ||
| RNA degradation | V | V | ||
| Phagosome | V | V | ||
| Tuberculosis | V | V | ||
| HIV-1 infection | V | V | ||
| Systemic lupus erythematosus | V | |||
| Alcoholism | V | |||
| Tuberculosis | V | |||
| Viral carcinogenesis | V |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mitra, S.; Harvey-Jones, K.; Kraev, I.; Verma, V.; Meehan, C.; Mintoft, A.; Norris, G.; Campbell, E.; Tucker, K.; Robertson, N.J.; et al. The Extracellular Vesicle Citrullinome and Signature in a Piglet Model of Neonatal Seizures. Int. J. Mol. Sci. 2023, 24, 11529. https://doi.org/10.3390/ijms241411529
Mitra S, Harvey-Jones K, Kraev I, Verma V, Meehan C, Mintoft A, Norris G, Campbell E, Tucker K, Robertson NJ, et al. The Extracellular Vesicle Citrullinome and Signature in a Piglet Model of Neonatal Seizures. International Journal of Molecular Sciences. 2023; 24(14):11529. https://doi.org/10.3390/ijms241411529
Chicago/Turabian StyleMitra, Subhabrata, Kelly Harvey-Jones, Igor Kraev, Vinita Verma, Christopher Meehan, Alison Mintoft, Georgina Norris, Ellie Campbell, Katie Tucker, Nicola J. Robertson, and et al. 2023. "The Extracellular Vesicle Citrullinome and Signature in a Piglet Model of Neonatal Seizures" International Journal of Molecular Sciences 24, no. 14: 11529. https://doi.org/10.3390/ijms241411529