

Macrophages Provide Essential Support for Erythropoiesis, and Extracellular ATP Contributes to a Erythropoiesis-Supportive Microenvironment during Repeated Psychological Stress
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. The Effects of Macrophage Depletion on Hematological Parameters
2.2. Depletion of Macrophages Abolishes the Effects of Repeated Restraint Stress on Erythroid Progenitors in the Bone Marrow and Spleen
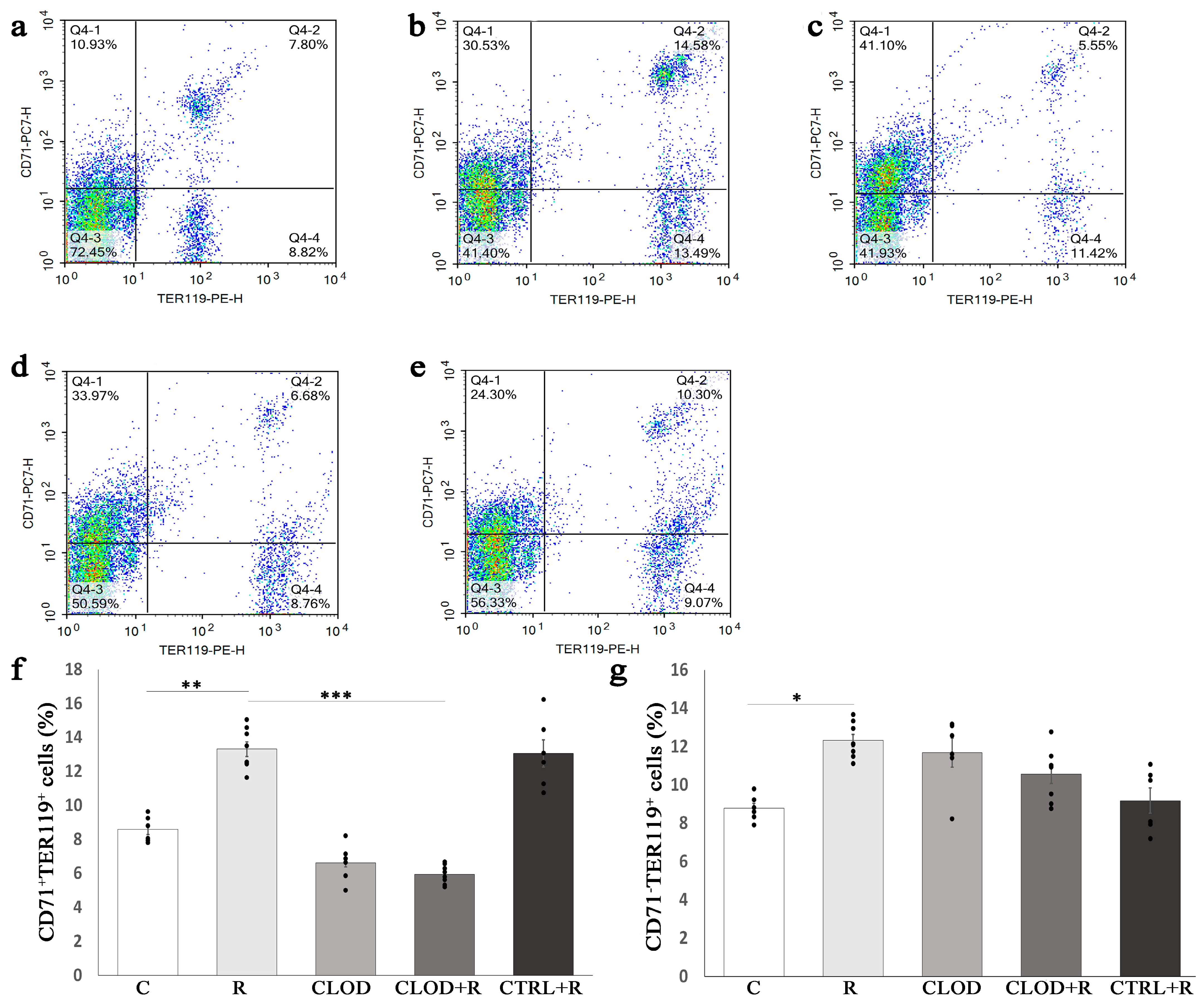
2.3. The Effects of Macrophage Depletion on Ter119-Positive Cells in the Bone Marrow and Spleen under Conditions of Repeated Stress
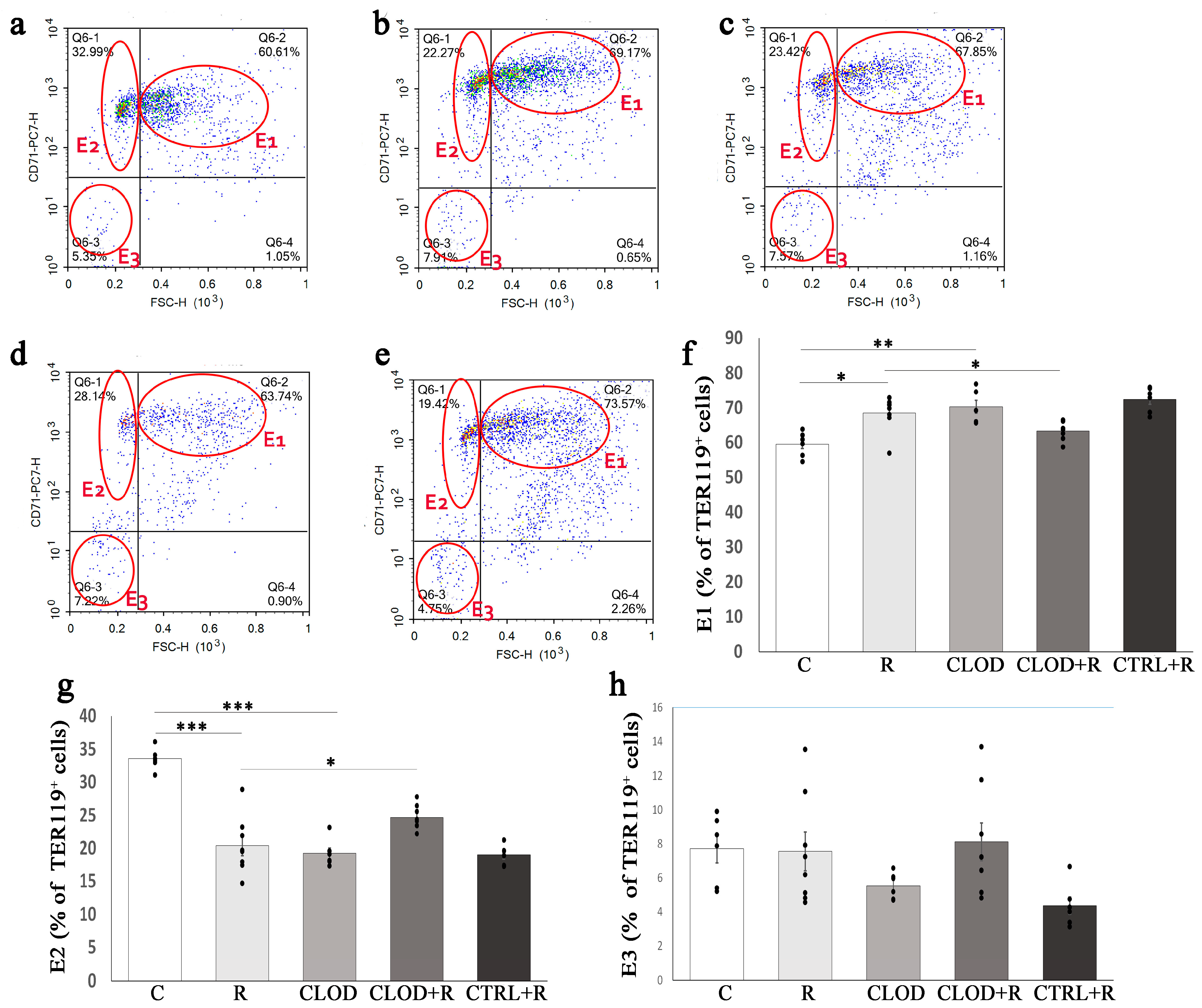
2.4. The Effects of Macrophage Depletion on Different Subpopulations of Ter119-Positive Cells in the Bone Marrow and Spleen upon Repeated Stress
2.5. Repeated Restraint Stress Increases ATP Levels and P2X7R Expression in the Bone Marrow and Spleen
2.6. Enhanced Specific Activity and Expression Levels of the Ectonucleotidase CD39 in the Bone Marrow and Spleen during Repeated Stress
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Procedure
4.3. Hematologic Parameters
4.4. Colony Assays
4.5. Flow Cytometry
4.6. RNA Extraction and Real-Time PCR
4.7. Extracellular ATP Measurement
4.8. CD39 Activity
4.9. Western Blot
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Paulson, R.F.; Shi, L.; Wu, D.C. Stress Erythropoiesis: New Signals and New Stress Progenitor Cells. Curr. Opin. Hematol. 2011, 18, 139–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alamo, I.G.; Kannan, K.B.; Loftus, T.J.; Ramos, H.; Efron, P.A.; Mohr, A.M. Severe Trauma and Chronic Stress Activates Extramedullary Erythropoiesis. J. Trauma Acute Care Surg. 2017, 83, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Paulson, R.F.; Ruan, B.; Hao, S.; Chen, Y. Stress Erythropoiesis Is a Key Inflammatory Response. Cells 2020, 9, 634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Liu, D.; Song, P.; Jiang, F.; Chi, X.; Zhang, T. Exposure to Hypoxia Causes Stress Erythropoiesis and Downregulates Immune Response Genes in Spleen of Mice. BMC Genom. 2021, 22, 413. [Google Scholar] [CrossRef]
- Millot, S.; Andrieu, V.; Letteron, P.; Lyoumi, S.; Hurtado-Nedelec, M.; Karim, Z.; Thibaudeau, O.; Bennada, S.; Charrier, J.L.; Lasocki, S.; et al. Erythropoietin Stimulates Spleen Bmp4-Dependent Stress Erythropoiesis and Partially Corrects Anemia in a Mouse Model of Generalized Inflammation. Blood 2010, 116, 6072–6081. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.C.; Paulson, R.F. Hypoxia Regulates Bmp4 Expression in the Murine Spleen During the Recovery from Acute Anemia. PLoS ONE 2010, 5, e11303. [Google Scholar] [CrossRef]
- Vignjevic, S.; Budec, M.; Markovic, D.; Dikic, D.; Mitrovic, O.; Mojsilovic, S.; Duric, S.V.; Koko, V.; Cokic, B.B.; Cokic, V.; et al. Chronic Psychological Stress Activates Bmp4-Dependent Extramedullary Erythropoiesis. J. Cell Mol. Med. 2014, 18, 91–103. [Google Scholar] [CrossRef]
- Paulson, R.F.; Hariharan, S.; Little, J.A. Stress Erythropoiesis: Definitions and Models for Its Study. Exp. Hematol. 2020, 89, 43–54.e2. [Google Scholar] [CrossRef]
- Klei, T.R.; Meinderts, S.M.; van den Berg, T.K.; van Bruggen, R. From the Cradle to the Grave: The Role of Macrophages in Erythropoiesis and Erythrophagocytosis. Front. Immunol. 2017, 8, 73. [Google Scholar] [CrossRef] [Green Version]
- Seyfried, A.N.; Maloney, J.M.; MacNamara, K.C. Macrophages Orchestrate Hematopoietic Programs and Regulate Hsc Function During Inflammatory Stress. Front. Immunol. 2020, 11, 1499. [Google Scholar] [CrossRef]
- Levesque, J.P.; Summers, K.M.; Millard, S.M.; Bisht, K.; Winkler, I.G.; Pettit, A.R. Role of Macrophages and Phagocytes in Orchestrating Normal and Pathologic Hematopoietic Niches. Exp. Hematol. 2021, 100, 12–31 e11. [Google Scholar] [CrossRef] [PubMed]
- McCabe, A.; MacNamara, K.C. Macrophages: Key Regulators of Steady-State and Demand-Adapted Hematopoiesis. Exp. Hematol. 2016, 44, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Guo, R.; Song, Y.; Jiang, Z. Erythroblastic Island Macrophages Shape Normal Erythropoiesis and Drive Associated Disorders in Erythroid Hematopoietic Diseases. Front. Cell Dev. Biol. 2020, 8, 613885. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, H.C. Niches for Extramedullary Hematopoiesis in the Spleen. Niche 2012, 1, 12–16. [Google Scholar] [CrossRef]
- Short, C.; Lim, H.K.; Tan, J.; O’Neill, H.C. Targeting the Spleen as an Alternative Site for Hematopoiesis. Bioessays 2019, 41, e1800234. [Google Scholar] [CrossRef] [Green Version]
- Liao, C.; Prabhu, K.S.; Paulson, R.F. Monocyte-Derived Macrophages Expand the Murine Stress Erythropoietic Niche During the Recovery from Anemia. Blood 2018, 132, 2580–2593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pixley, F.J. Macrophage Migration and Its Regulation by Csf-1. Int. J. Cell Biol. 2012, 2012, 501962. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Takemura, N.; Saitoh, T. Macrophage Response Driven by Extracellular Atp. Biol. Pharm. Bull. 2021, 44, 599–604. [Google Scholar] [CrossRef]
- Zhang, X.; Mosser, D.M. Macrophage Activation by Endogenous Danger Signals. J. Pathol. 2008, 214, 161–178. [Google Scholar] [CrossRef]
- Savio, L.E.B.; de Andrade Mello, P.; Figliuolo, V.R.; de Avelar Almeida, T.F.; Santana, P.T.; Oliveira, S.D.S.; Silva, C.L.M.; Feldbrugge, L.; Csizmadia, E.; Minshall, R.D.; et al. Cd39 Limits P2x7 Receptor Inflammatory Signaling and Attenuates Sepsis-Induced Liver Injury. J. Hepatol. 2017, 67, 716–726. [Google Scholar] [CrossRef] [Green Version]
- Vignjevic, S.; Budec, M.; Markovic, D.; Dikic, D.; Mitrovic, O.; Diklic, M.; Suboticki, T.; Cokic, V.; Jovcic, G. Glucocorticoid Receptor Mediates the Expansion of Splenic Late Erythroid Progenitors During Chronic Psychological Stress. J. Physiol. Pharmacol. 2015, 66, 91–100. [Google Scholar] [PubMed]
- Vignjevic Petrinovic, S.; Budec, M.; Markovic, D.; Gotic, M.; Mitrovic Ajtic, O.; Mojsilovic, S.; Stosic-Grujicic, S.; Ivanov, M.; Jovcic, G.; Cokic, V. Macrophage Migration Inhibitory Factor Is an Endogenous Regulator of Stress-Induced Extramedullary Erythropoiesis. Histochem. Cell Biol. 2016, 146, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Vignjevic Petrinovic, S.; Budec, M.; Markovic, D.; Mitrovic Ajtic, O.; Jovcic, G.; Milosevic, M.; Momcilovic, S.; Cokic, V. Nitric Oxide-Dependent Expansion of Erythroid Progenitors in a Murine Model of Chronic Psychological Stress. Histochem. Cell Biol. 2020, 153, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Lagraauw, H.M.; Kuiper, J.; Bot, I. Acute and Chronic Psychological Stress as Risk Factors for Cardiovascular Disease: Insights Gained from Epidemiological, Clinical and Experimental Studies. Brain Behav. Immun. 2015, 50, 18–30. [Google Scholar] [CrossRef]
- Merabet, N.; Lucassen, P.J.; Crielaard, L.; Stronks, K.; Quax, R.; Sloot, P.M.A.; la Fleur, S.E.; Nicolaou, M. How Exposure to Chronic Stress Contributes to the Development of Type 2 Diabetes: A Complexity Science Approach. Front. Neuroendocrinol. 2022, 65, 100972. [Google Scholar] [CrossRef] [PubMed]
- Vignjevic Petrinovic, S.; Jaukovic, A.; Milosevic, M.; Bugarski, D.; Budec, M. Targeting Stress Erythropoiesis Pathways in Cancer. Front. Physiol. 2022, 13, 844042. [Google Scholar] [CrossRef] [PubMed]
- Dhabhar, F.S. Effects of Stress on Immune Function: The Good, the Bad, and the Beautiful. Immunol. Res. 2014, 58, 193–210. [Google Scholar] [CrossRef]
- Munshi, S.; Loh, M.K.; Ferrara, N.; DeJoseph, M.R.; Ritger, A.; Padival, M.; Record, M.J.; Urban, J.H.; Rosenkranz, J.A. Repeated Stress Induces a Pro-Inflammatory State, Increases Amygdala Neuronal and Microglial Activation, and Causes Anxiety in Adult Male Rats. Brain Behav. Immun. 2020, 84, 180–199. [Google Scholar] [CrossRef]
- Voorhees, J.L.; Powell, N.D.; Moldovan, L.; Mo, X.; Eubank, T.D.; Marsh, C.B. Chronic Restraint Stress Upregulates Erythropoiesis through Glucocorticoid Stimulation. PLoS ONE 2013, 8, e77935. [Google Scholar] [CrossRef]
- McLaughlin, K.J.; Gomez, J.L.; Baran, S.E.; Conrad, C.D. The Effects of Chronic Stress on Hippocampal Morphology and Function: An Evaluation of Chronic Restraint Paradigms. Brain Res. 2007, 1161, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Buynitsky, T.; Mostofsky, D.I. Restraint Stress in Biobehavioral Research: Recent Developments. Neurosci. Biobehav. Rev. 2009, 33, 1089–1098. [Google Scholar] [CrossRef]
- Bader, J.E.; Enos, R.T.; Velázquez, K.T.; Carson, M.S.; Sougiannis, A.T.; McGuinness, O.P.; Robinson, C.M.; Murphy, E.A. Repeated clodronate-liposome treatment results in neutrophilia and is not effective in limiting obesity-linked metabolic impairments. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E358–E372. [Google Scholar] [CrossRef]
- Ramos, P.; Casu, C.; Gardenghi, S.; Breda, L.; Crielaard, B.J.; Guy, E.; Marongiu, M.F.; Gupta, R.; Levine, R.L.; Abdel-Wahab, O.; et al. Macrophages Support Pathological Erythropoiesis in Polycythemia Vera and Beta-Thalassemia. Nat. Med. 2013, 19, 437–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Chen, J.; Wang, W.; Wang, L.; Ma, L.; Shen, H.; Li, M. Psychological stress induces hypoferremia through the IL-6–hepcidin axis in rats. Biochem. Biophys. Res. Commun. 2008, 373, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Theurl, M.; Theurl, I.; Hochegger, K.; Obrist, P.; Subramaniam, N.; van Rooijen, N.; Schuemann, K.; Weiss, G. Kupffer cells modulate iron homeostasis in mice via regulation of hepcidin expression. J. Mol. Med. 2008, 86, 825–835. [Google Scholar] [CrossRef] [PubMed]
- Chasis, J.A.; Mohandas, N. Erythroblastic Islands: Niches for Erythropoiesis. Blood 2008, 112, 470–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dulmovits, B.M.; Hom, J.; Narla, A.; Mohandas, N.; Blanc, L. Characterization, Regulation, and Targeting of Erythroid Progenitors in Normal and Disordered Human Erythropoiesis. Curr. Opin. Hematol. 2017, 24, 159–166. [Google Scholar] [CrossRef]
- Munugalavadla, V.; Kapur, R. Role of C-Kit and Erythropoietin Receptor in Erythropoiesis. Crit. Rev. Oncol. Hematol. 2005, 54, 63–75. [Google Scholar] [CrossRef]
- Hao, S.; Xiang, J.; Wu, D.C.; Fraser, J.W.; Ruan, B.; Cai, J.; Patterson, A.D.; Lai, Z.C.; Paulson, R.F. Gdf15 Regulates Murine Stress Erythroid Progenitor Proliferation and the Development of the Stress Erythropoiesis Niche. Blood Adv. 2019, 3, 2205–2217. [Google Scholar] [CrossRef]
- Trivedi, G.; Inoue, D.; Chen, C.; Bitner, L.; Chung, Y.R.; Taylor, J.; Gönen, M.; Wess, J.; Wahab, O.A.; Zhang, L. Muscarinic acetylcholine receptor regulates self-renewal of early erythroid progenitors. Sci. Transl. Med. 2019, 11, eaaw3781. [Google Scholar] [CrossRef]
- Myslivecek, J.; Rícný, J.; Palkovits, M.; Kvetnanský, R. The effects of short-term immobilization stress on muscarinic receptors, beta-adrenoceptors, and adenylyl cyclase in different heart regions. Ann. N. Y. Acad. Sci. 2004, 1018, 315–322. [Google Scholar] [CrossRef]
- Myslivecek, J.; Tillinger, A.; Novakova, M.; Kvetnanský, R. Regulation of adrenoceptor and muscarinic receptor gene expression after single and repeated stress. Ann. N. Y. Acad. Sci. 2008, 1148, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Chow, A.; Huggins, M.; Ahmed, J.; Hashimoto, D.; Lucas, D.; Kunisaki, Y.; Pinho, S.; Leboeuf, M.; Noizat, C.; van Rooijen, N.; et al. Cd169(+) Macrophages Provide a Niche Promoting Erythropoiesis under Homeostasis and Stress. Nat. Med. 2013, 19, 429–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Jin, X.; He, X.; Pan, L.; Zhang, X.; Zhao, Y. Macrophages Support Splenic Erythropoiesis in 4t1 Tumor-Bearing Mice. PLoS ONE 2015, 10, e0121921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kina, T.; Ikuta, K.; Takayama, E.; Wada, K.; Majumdar, A.S.; Weissman, I.L.; Katsura, Y. The Monoclonal Antibody Ter-119 Recognizes a Molecule Associated with Glycophorin a and Specifically Marks the Late Stages of Murine Erythroid Lineage. Br. J. Haematol. 2000, 109, 280–287. [Google Scholar] [CrossRef]
- Koury, M.J.; Bondurant, M.C.; Atkinson, J.B. Erythropoietin Control of Terminal Erythroid Differentiation: Maintenance of Cell Viability, Production of Hemoglobin, and Development of the Erythrocyte Membrane. Blood Cells 1987, 13, 217–226. [Google Scholar]
- Perron-Deshaies, G.; St-Louis, P.; Romero, H.; Scorza, T. Impact of Erythropoietin Production by Erythroblastic Island Macrophages on Homeostatic Murine Erythropoiesis. Int. J. Mol. Sci. 2020, 21, 8930. [Google Scholar] [CrossRef]
- Yanai, N.; Satoh, T.; Obinata, M. Endothelial Cells Create a Hematopoietic Inductive Microenvironment Preferential to Erythropoiesis in the Mouse Spleen. Cell Struct. Funct. 1991, 16, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Greenwald, A.C.; Licht, T.; Kumar, S.; Oladipupo, S.S.; Iyer, S.; Grunewald, M.; Keshet, E. Vegf Expands Erythropoiesis Via Hypoxia-Independent Induction of Erythropoietin in Noncanonical Perivascular Stromal Cells. J. Exp. Med. 2019, 216, 215–230. [Google Scholar] [CrossRef] [Green Version]
- Stellacci, E.; Di Noia, A.; Di Baldassarre, A.; Migliaccio, G.; Battistini, A.; Migliaccio, A.R. Interaction between the Glucocorticoid and Erythropoietin Receptors in Human Erythroid Cells. Exp. Hematol. 2009, 37, 559–572. [Google Scholar] [CrossRef] [Green Version]
- Haas, N.; Riedt, T.; Labbaf, Z.; Bassler, K.; Gergis, D.; Frohlich, H.; Gutgemann, I.; Janzen, V.; Schorle, H. Kit Transduced Signals Counteract Erythroid Maturation by Mapk-Dependent Modulation of Erythropoietin Signaling and Apoptosis Induction in Mouse Fetal Liver. Cell Death Differ. 2015, 22, 790–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierce, H.; Zhang, D.; Magnon, C.; Lucas, D.; Christin, J.R.; Huggins, M.; Schwartz, G.J.; Frenette, P.S. Cholinergic Signals from the CNS Regulate G-CSF-Mediated HSC Mobilization from Bone Marrow via a Glucocorticoid Signaling Relay. Cell Stem. Cell 2017, 20, 648–658.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mende, N.; Laurenti, E. Hematopoietic Stem and Progenitor Cells Outside the Bone Marrow: Where, When, and Why. Exp. Hematol. 2021, 104, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, M.Z.; Adamiak, M.; Plonka, M.; Abdel-Latif, A.; Ratajczak, J. Mobilization of Hematopoietic Stem Cells as a Result of Innate Immunity-Mediated Sterile Inflammation in the Bone Marrow Microenvironment-the Involvement of Extracellular Nucleotides and Purinergic Signaling. Leukemia 2018, 32, 1116–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tung, L.T.; Wang, H.; Belle, J.I.; Petrov, J.C.; Langlais, D.; Nijnik, A. P53-Dependent Induction of P2x7 on Hematopoietic Stem and Progenitor Cells Regulates Hematopoietic Response to Genotoxic Stress. Cell Death Dis. 2021, 12, 923. [Google Scholar] [CrossRef]
- Lee, G.R.; Shaefi, S.; Otterbein, L.E. HO-1 and CD39: It Takes Two to Protect the Realm. Front. Immunol. 2019, 10, 1765. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Liu, R.; Nemkov, T.; Couturier, J.; Liang, L.; Song, A.; Zhao, S.; Sun, K.; Adebiyi, M.; Wen, Y.E.; et al. Adenosine A2b Receptor Controls Erythroid Lineage Commitment in Stress Erythropoiesis by Promoting Metabolic Reprogramming. Blood 2018, 132, 845. [Google Scholar] [CrossRef]
- Baykov, A.A.; Evtushenko, O.A.; Avaeva, S.M. A Malachite Green Procedure for Orthophosphate Determination and Its Use in Alkaline Phosphatase-Based Enzyme Immunoassay. Anal. Biochem. 1988, 171, 266–270. [Google Scholar] [CrossRef]
- Markwell, M.A.; Haas, S.M.; Bieber, L.L.; Tolbert, N.E. A Modification of the Lowry Procedure to Simplify Protein Determination in Membrane and Lipoprotein Samples. Anal. Biochem. 1978, 87, 206–210. [Google Scholar] [CrossRef]
- Haidl, I.D.; Jefferies, W.A. The macrophage cell surface glycoprotein F4/80 is a highly glycosylated proteoglycan. Eur. J. Immunol. 1996, 5, 1139–1146. [Google Scholar] [CrossRef]
- Austyn, J.M.; Gordon, S. F4/80, a monoclonal antibody directed specifically against the mouse macrophage. Eur. J. Immunol. 1981, 10, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative Expression Software Tool (Rest) for Group-Wise Comparison and Statistical Analysis of Relative Expression Results in Real-Time Pcr. Nucleic. Acids. Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | R | CLOD | CLOD + R | CTRL + R | |
|---|---|---|---|---|---|
| RBC (1012/L) | 5.52 ± 0.16 | 5.35 ± 0.18 | 4.54 ± 0.12 | 3.95 ± 0.06 *(a)***(b) | 5.18 ± 0.19 |
| Hb (g/L) | 165.34 ± 1.95 | 130.57 ± 2.68 ***(a) | 140.26 ± 3.14 ***(a) | 100.68 ± 1.68 ***(a)***(b) | 132.94 ± 2.48 |
| Hct (%) | 45.67 ± 1.17 | 43.75 ± 1.71 | 40.67 ± 0.56 **(a) | 39.12 ± 0.64 **(a) | 42.83 ± 1.3 |
| MCV (fl) | 83.78 ± 3.84 | 86.26 ± 1.79 | 84.66 ± 2.49 | 92.51 ± 4.45 | 85.65 ± 2.26 |
| MCH (pg) | 30.06 ± 0.88 | 24.41 ± 0.72 **(a) | 27.26 ± 0.9 | 24.72 ± 0.73 **(a) | 25.84 ± 1.14 *(a) |
| MCHC (g/L) | 363.18 ± 9.54 | 301.56 ± 13.03 **(a) | 344.94 ± 6.6 | 278.20 ± 5.42 ***(a) | 311.58 ± 9.87 *(a) |
| RDW (%) | 11.36 ± 0.54 | 14.01 ± 0.28 *(a) | 11.97 ± 0.38 | 13.46 ± 0.87 | 13.78 ± 0.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Momčilović, S.; Bogdanović, A.; Milošević, M.S.; Mojsilović, S.; Marković, D.C.; Kočović, D.M.; Vignjević Petrinović, S. Macrophages Provide Essential Support for Erythropoiesis, and Extracellular ATP Contributes to a Erythropoiesis-Supportive Microenvironment during Repeated Psychological Stress. Int. J. Mol. Sci. 2023, 24, 11373. https://doi.org/10.3390/ijms241411373
Momčilović S, Bogdanović A, Milošević MS, Mojsilović S, Marković DC, Kočović DM, Vignjević Petrinović S. Macrophages Provide Essential Support for Erythropoiesis, and Extracellular ATP Contributes to a Erythropoiesis-Supportive Microenvironment during Repeated Psychological Stress. International Journal of Molecular Sciences. 2023; 24(14):11373. https://doi.org/10.3390/ijms241411373
Chicago/Turabian StyleMomčilović, Sanja, Andrija Bogdanović, Maja S. Milošević, Slavko Mojsilović, Dragana C. Marković, Dušica M. Kočović, and Sanja Vignjević Petrinović. 2023. "Macrophages Provide Essential Support for Erythropoiesis, and Extracellular ATP Contributes to a Erythropoiesis-Supportive Microenvironment during Repeated Psychological Stress" International Journal of Molecular Sciences 24, no. 14: 11373. https://doi.org/10.3390/ijms241411373