
Transcriptional Readthrough Interrupts Boundary Function in Drosophila

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
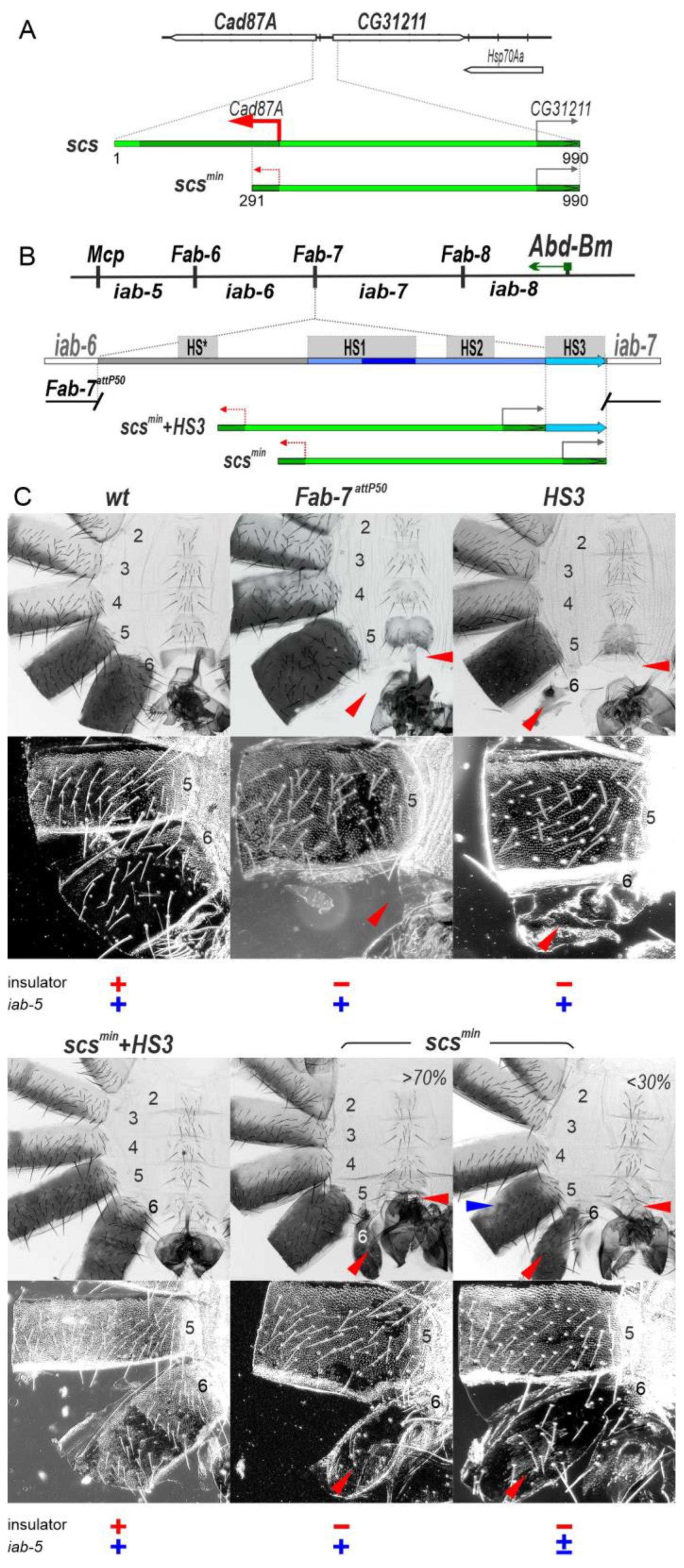
2.1. The scsmin Insulator Can Block Crosstalk between the iab-5 and iab-6 Domains Only in Cooperation with the iab-7 PRE
2.2. Transcription Induced by the Cad87A Promoter in scs Can Affect the Activity of the iab-7 Domain
2.3. Transcription Disrupts Boundary Function
2.4. Transcription Disrupts the Functioning of the iab-6 and iab-5 Regulatory Domains
2.5. Readthrough Transcription Disrupts the Functioning of a Minimal Fub (pHS2) Replacement Boundary
2.6. Readthrough Transcription Disrupts Fub Function in Its Endogenous Context
3. Discussion
4. Materials and Methods
4.1. Generation of the Replacement Lines
4.2. Cuticle Preparations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cavalheiro, G.R.; Pollex, T.; Furlong, E.E. To Loop or Not to Loop: What Is the Role of TADs in Enhancer Function and Gene Regulation? Curr. Opin. Genet. Dev. 2021, 67, 119–129. [Google Scholar] [CrossRef]
- Hafner, A.; Boettiger, A. The Spatial Organization of Transcriptional Control. Nat. Rev. Genet. 2022, 24, 53–68. [Google Scholar] [CrossRef]
- Jerkovic, I.; Cavalli, G. Understanding 3D Genome Organization by Multidisciplinary Methods. Nat. Rev. Mol. Cell Biol. 2021, 22, 511–528. [Google Scholar] [CrossRef]
- Mir, M.; Bickmore, W.; Furlong, E.E.M.; Narlikar, G. Chromatin Topology, Condensates and Gene Regulation: Shifting Paradigms or Just a Phase? Development 2019, 146, dev182766. [Google Scholar] [CrossRef] [Green Version]
- Batut, P.J.; Bing, X.Y.; Sisco, Z.; Raimundo, J.; Levo, M.; Levine, M.S. Genome Organization Controls Transcriptional Dynamics during Development. Science 2022, 375, 566–570. [Google Scholar] [CrossRef]
- Kyrchanova, O.; Georgiev, P. Mechanisms of Enhancer-Promoter Interactions in Higher Eukaryotes. Int. J. Mol. Sci. 2021, 22, 671. [Google Scholar] [CrossRef]
- Matthews, N.E.; White, R. Chromatin Architecture in the Fly: Living without CTCF/Cohesin Loop Extrusion? Alternating Chromatin States Provide a Basis for Domain Architecture in Drosophila. Bioessays 2019, 41, e1900048. [Google Scholar] [CrossRef] [Green Version]
- Melnikova, L.S.; Georgiev, P.G.; Golovnin, A.K. The Functions and Mechanisms of Action of Insulators in the Genomes of Higher Eukaryotes. Acta Naturae 2020, 12, 15–33. [Google Scholar] [CrossRef]
- Chen, D.; Lei, E.P. Function and Regulation of Chromatin Insulators in Dynamic Genome Organization. Curr. Opin. Cell Biol. 2019, 58, 61–68. [Google Scholar] [CrossRef]
- Kyrchanova, O.V.; Bylino, O.V.; Georgiev, P.G. Mechanisms of Enhancer-Promoter Communication and Chromosomal Architecture in Mammals and Drosophila. Front. Genet. 2022, 13, 1081088. [Google Scholar] [CrossRef]
- Maksimenko, O.G.; Fursenko, D.V.; Belova, E.V.; Georgiev, P.G. CTCF As an Example of DNA-Binding Transcription Factors Containing Clusters of C2H2-Type Zinc Fingers. Acta Naturae 2021, 13, 31–46. [Google Scholar] [CrossRef]
- Chetverina, D.; Fujioka, M.; Erokhin, M.; Georgiev, P.; Jaynes, J.B.; Schedl, P. Boundaries of Loop Domains (Insulators): Determinants of Chromosome Form and Function in Multicellular Eukaryotes. Bioessays 2017, 39, 1600233. [Google Scholar] [CrossRef] [Green Version]
- Matzat, L.H.; Lei, E.P. Surviving an Identity Crisis: A Revised View of Chromatin Insulators in the Genomics Era. Biochim. Biophys. Acta 2014, 1839, 203–214. [Google Scholar] [CrossRef] [Green Version]
- Kyrchanova, O.; Maksimenko, O.; Ibragimov, A.; Sokolov, V.; Postika, N.; Lukyanova, M.; Schedl, P.; Georgiev, P. The Insulator Functions of the Drosophila Polydactyl C2H2 Zinc Finger Protein CTCF: Necessity versus Sufficiency. Sci. Adv. 2020, 6, eaaz3152. [Google Scholar] [CrossRef] [Green Version]
- Kyrchanova, O.; Mogila, V.; Wolle, D.; Magbanua, J.P.; White, R.; Georgiev, P.; Schedl, P. The Boundary Paradox in the Bithorax Complex. Mech. Dev. 2015, 138 Pt 2, 122–132. [Google Scholar] [CrossRef]
- Lewis, E.B. A Gene Complex Controlling Segmentation in Drosophila. Nature 1978, 276, 565–570. [Google Scholar] [CrossRef]
- Maeda, R.K.; Karch, F. The ABC of the BX-C: The Bithorax Complex Explained. Development 2006, 133, 1413–1422. [Google Scholar] [CrossRef] [Green Version]
- Maeda, R.K.; Karch, F. The Open for Business Model of the Bithorax Complex in Drosophila. Chromosoma 2015, 124, 293–307. [Google Scholar] [CrossRef] [Green Version]
- Celniker, S.E.; Sharma, S.; Keelan, D.J.; Lewis, E.B. The Molecular Genetics of the Bithorax Complex of Drosophila: Cis-Regulation in the Abdominal-B Domain. EMBO J. 1990, 9, 4277–4286. [Google Scholar] [CrossRef]
- Duncan, I. The Bithorax Complex. Annu. Rev. Genet. 1987, 21, 285–319. [Google Scholar] [CrossRef]
- Garaulet, D.L.; Lai, E.C. Hox MiRNA Regulation within the Drosophila Bithorax Complex: Patterning Behavior. Mech. Dev. 2015, 138 Pt 2, 151–159. [Google Scholar] [CrossRef]
- Hagstrom, K.; Muller, M.; Schedl, P. Fab-7 Functions as a Chromatin Domain Boundary to Ensure Proper Segment Specification by the Drosophila Bithorax Complex. Genes Dev. 1996, 10, 3202–3215. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Herrero, E. Control of the Expression of the Bithorax Complex Genes Abdominal-A and Abdominal-B by Cis-Regulatory Regions in Drosophila Embryos. Development 1991, 111, 437–449. [Google Scholar] [CrossRef]
- Casares, F.; Sánchez-Herrero, E. Regulation of the Infraabdominal Regions of the Bithorax Complex of Drosophila by Gap Genes. Development 1995, 121, 1855–1866. [Google Scholar] [CrossRef]
- Drewell, R.A.; Nevarez, M.J.; Kurata, J.S.; Winkler, L.N.; Li, L.; Dresch, J.M. Deciphering the Combinatorial Architecture of a Drosophila Homeotic Gene Enhancer. Mech. Dev. 2014, 131, 68–77. [Google Scholar] [CrossRef]
- Iampietro, C.; Gummalla, M.; Mutero, A.; Karch, F.; Maeda, R.K. Initiator Elements Function to Determine the Activity State of BX-C Enhancers. PLoS Genet. 2010, 6, e1001260. [Google Scholar] [CrossRef] [Green Version]
- Bowman, S.K.; Deaton, A.M.; Domingues, H.; Wang, P.I.; Sadreyev, R.I.; Kingston, R.E.; Bender, W. H3K27 Modifications Define Segmental Regulatory Domains in the Drosophila Bithorax Complex. eLife 2014, 3, e02833. [Google Scholar] [CrossRef]
- Cheutin, T.; Cavalli, G. The Multiscale Effects of Polycomb Mechanisms on 3D Chromatin Folding. Crit. Rev. Biochem. Mol. Biol. 2019, 54, 399–417. [Google Scholar] [CrossRef]
- Chiang, A.; O’Connor, M.B.; Paro, R.; Simon, J.; Bender, W. Discrete Polycomb-Binding Sites in Each Parasegmental Domain of the Bithorax Complex. Development 1995, 121, 1681–1689. [Google Scholar] [CrossRef]
- Kassis, J.A.; Kennison, J.A.; Tamkun, J.W. Polycomb and Trithorax Group Genes in Drosophila. Genetics 2017, 206, 1699–1725. [Google Scholar] [CrossRef] [Green Version]
- Barges, S.; Mihaly, J.; Galloni, M.; Hagstrom, K.; Müller, M.; Shanower, G.; Schedl, P.; Gyurkovics, H.; Karch, F. The Fab-8 Boundary Defines the Distal Limit of the Bithorax Complex Iab-7 Domain and Insulates Iab-7 from Initiation Elements and a PRE in the Adjacent Iab-8 Domain. Development 2000, 127, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Bender, W.; Lucas, M. The Border between the Ultrabithorax and Abdominal-A Regulatory Domains in the Drosophila Bithorax Complex. Genetics 2013, 193, 1135–1147. [Google Scholar] [CrossRef] [Green Version]
- Galloni, M.; Gyurkovics, H.; Schedl, P.; Karch, F. The Bluetail Transposon: Evidence for Independent Cis-Regulatory Domains and Domain Boundaries in the Bithorax Complex. EMBO J. 1993, 12, 1087–1097. [Google Scholar] [CrossRef]
- Gyurkovics, H.; Gausz, J.; Kummer, J.; Karch, F. A New Homeotic Mutation in the Drosophila Bithorax Complex Removes a Boundary Separating Two Domains of Regulation. EMBO J. 1990, 9, 2579–2585. [Google Scholar] [CrossRef]
- Karch, F.; Galloni, M.; Sipos, L.; Gausz, J.; Gyurkovics, H.; Schedl, P. Mcp and Fab-7: Molecular Analysis of Putative Boundaries of Cis-Regulatory Domains in the Bithorax Complex of Drosophila Melanogaster. Nucleic Acids Res. 1994, 22, 3138–3146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihaly, J.; Hogga, I.; Gausz, J.; Gyurkovics, H.; Karch, F. In Situ Dissection of the Fab-7 Region of the Bithorax Complex into a Chromatin Domain Boundary and a Polycomb-Response Element. Development 1997, 124, 1809–1820. [Google Scholar] [CrossRef]
- Kyrchanova, O.; Sokolov, V.; Georgiev, P. Mechanisms of Interaction between Enhancers and Promoters in Three Drosophila Model Systems. Int. J. Mol. Sci. 2023, 24, 2855. [Google Scholar] [CrossRef]
- Kyrchanova, O.; Sabirov, M.; Mogila, V.; Kurbidaeva, A.; Postika, N.; Maksimenko, O.; Schedl, P.; Georgiev, P. Complete Reconstitution of Bypass and Blocking Functions in a Minimal Artificial Fab-7 Insulator from Drosophila Bithorax Complex. Proc. Natl. Acad. Sci. USA 2019, 116, 13462–13467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyrchanova, O.; Wolle, D.; Sabirov, M.; Kurbidaeva, A.; Aoki, T.; Maksimenko, O.; Kyrchanova, M.; Georgiev, P.; Schedl, P. Distinct Elements Confer the Blocking and Bypass Functions of the Bithorax Fab-8 Boundary. Genetics 2019, 213, 865–876. [Google Scholar] [CrossRef]
- Postika, N.; Metzler, M.; Affolter, M.; Müller, M.; Schedl, P.; Georgiev, P.; Kyrchanova, O. Boundaries Mediate Long-Distance Interactions between Enhancers and Promoters in the Drosophila Bithorax Complex. PLoS Genet. 2018, 14, e1007702. [Google Scholar] [CrossRef] [Green Version]
- Hogga, I.; Karch, F. Transcription through the Iab-7 Cis-Regulatory Domain of the Bithorax Complex Interferes with Maintenance of Polycomb-Mediated Silencing. Development 2002, 129, 4915–4922. [Google Scholar] [CrossRef]
- Hogga, I.; Mihaly, J.; Barges, S.; Karch, F. Replacement of Fab-7 by the Gypsy or Scs Insulator Disrupts Long-Distance Regulatory Interactions in the Abd-B Gene of the Bithorax Complex. Mol. Cell 2001, 8, 1145–1151. [Google Scholar] [CrossRef]
- Kuhn, E.J.; Hart, C.M.; Geyer, P.K. Studies of the Role of the Drosophila Scs and Scs’ Insulators in Defining Boundaries of a Chromosome Puff. Mol. Cell Biol. 2004, 24, 1470–1480. [Google Scholar] [CrossRef]
- Kyrchanova, O.; Leman, D.; Parshikov, A.; Fedotova, A.; Studitsky, V.; Maksimenko, O.; Georgiev, P. New Properties of Drosophila Scs and Scs’ Insulators. PLoS ONE 2013, 8, e62690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez, J.; Schedl, P. Sequences Required for Enhancer Blocking Activity of Scs Are Located within Two Nuclease-Hypersensitive Regions. EMBO J. 1994, 13, 5984–5993. [Google Scholar] [CrossRef] [PubMed]
- Kyrchanova, O.; Kurbidaeva, A.; Sabirov, M.; Postika, N.; Wolle, D.; Aoki, T.; Maksimenko, O.; Mogila, V.; Schedl, P.; Georgiev, P. The Bithorax Complex Iab-7 Polycomb Response Element Has a Novel Role in the Functioning of the Fab-7 Chromatin Boundary. PLoS Genet. 2018, 14, e1007442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagstrom, K.; Muller, M.; Schedl, P. A Polycomb and GAGA Dependent Silencer Adjoins the Fab-7 Boundary in the Drosophila Bithorax Complex. Genetics 1997, 146, 1365–1380. [Google Scholar] [CrossRef]
- Mishra, R.K.; Mihaly, J.; Barges, S.; Spierer, A.; Karch, F.; Hagstrom, K.; Schweinsberg, S.E.; Schedl, P. The Iab-7 Polycomb Response Element Maps to a Nucleosome-Free Region of Chromatin and Requires Both GAGA and Pleiohomeotic for Silencing Activity. Mol. Cell Biol. 2001, 21, 1311–1318. [Google Scholar] [CrossRef] [Green Version]
- Wolle, D.; Cleard, F.; Aoki, T.; Deshpande, G.; Schedl, P.; Karch, F. Functional Requirements for Fab-7 Boundary Activity in the Bithorax Complex. Mol. Cell Biol. 2015, 35, 3739–3752. [Google Scholar] [CrossRef] [Green Version]
- Holohan, E.E.; Kwong, C.; Adryan, B.; Bartkuhn, M.; Herold, M.; Renkawitz, R.; Russell, S.; White, R. CTCF Genomic Binding Sites in Drosophila and the Organisation of the Bithorax Complex. PLoS Genet. 2007, 3, e112. [Google Scholar] [CrossRef] [Green Version]
- Sabirov, M.; Kyrchanova, O.; Pokholkova, G.V.; Bonchuk, A.; Klimenko, N.; Belova, E.; Zhimulev, I.F.; Maksimenko, O.; Georgiev, P. Mechanism and Functional Role of the Interaction between CP190 and the Architectural Protein Pita in Drosophila Melanogaster. Epigenet. Chromatin 2021, 14, 16. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, Y.B.; Linder-Basso, D.; Kharchenko, P.V.; Tolstorukov, M.Y.; Kim, M.; Li, H.-B.; Gorchakov, A.A.; Minoda, A.; Shanower, G.; Alekseyenko, A.A.; et al. Nature and Function of Insulator Protein Binding Sites in the Drosophila Genome. Genome Res. 2012, 22, 2188–2198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bender, W.; Fitzgerald, D.P. Transcription Activates Repressed Domains in the Drosophila Bithorax Complex. Development 2002, 129, 4923–4930. [Google Scholar] [CrossRef] [PubMed]
- Majumder, P.; Cai, H.N. The Functional Analysis of Insulator Interactions in the Drosophila Embryo. Proc. Natl. Acad. Sci. USA 2003, 100, 5223–5228. [Google Scholar] [CrossRef]
- Kuhn, E.J.; Viering, M.M.; Rhodes, K.M.; Geyer, P.K. A Test of Insulator Interactions in Drosophila. EMBO J. 2003, 22, 2463–2471. [Google Scholar] [CrossRef] [Green Version]
- Gohl, D.; Aoki, T.; Blanton, J.; Shanower, G.; Kappes, G.; Schedl, P. Mechanism of Chromosomal Boundary Action: Roadblock, Sink, or Loop? Genetics 2011, 187, 731–748. [Google Scholar] [CrossRef]
- Kyrchanova, O.; Chetverina, D.; Maksimenko, O.; Kullyev, A.; Georgiev, P. Orientation-Dependent Interaction between Drosophila Insulators Is a Property of This Class of Regulatory Elements. Nucleic Acids Res. 2008, 36, 7019–7028. [Google Scholar] [CrossRef] [Green Version]
- Gaszner, M.; Vazquez, J.; Schedl, P. The Zw5 Protein, a Component of the Scs Chromatin Domain Boundary, Is Able to Block Enhancer-Promoter Interaction. Genes Dev. 1999, 13, 2098–2107. [Google Scholar] [CrossRef] [Green Version]
- Castro Alvarez, J.J.; Revel, M.; Carrasco, J.; Cléard, F.; Pauli, D.; Hilgers, V.; Karch, F.; Maeda, R.K. Repression of the Hox Gene Abd-A by ELAV-Mediated Transcriptional Interference. PLoS Genet. 2021, 17, e1009843. [Google Scholar] [CrossRef]
- Erokhin, M.; Davydova, A.; Parshikov, A.; Studitsky, V.M.; Georgiev, P.; Chetverina, D. Transcription through Enhancers Suppresses Their Activity in Drosophila. Epigenet. Chromatin 2013, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Fujioka, M.; Nezdyur, A.; Jaynes, J.B. An Insulator Blocks Access to Enhancers by an Illegitimate Promoter, Preventing Repression by Transcriptional Interference. PLoS Genet. 2021, 17, e1009536. [Google Scholar] [CrossRef] [PubMed]
- Pande, A.; Brosius, J.; Makalowska, I.; Makalowski, W.; Raabe, C.A. Transcriptional Interference by Small Transcripts in Proximal Promoter Regions. Nucleic Acids Res. 2018, 46, 1069–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pande, A.; Makalowski, W.; Brosius, J.; Raabe, C.A. Enhancer Occlusion Transcripts Regulate the Activity of Human Enhancer Domains via Transcriptional Interference: A Computational Perspective. Nucleic Acids Res. 2020, 48, 3435–3454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibragimov, A.; Bing, X.; Shidlovskii, Y.; Levine, M.; Georgiev, P.; Schedl, P. The Insulating Activity of the Drosophila BX-C Chromatin Boundary Fub-1 Is Parasegmentally Regulated by LncRNA Read-Through. bioRxiv 2023. [Google Scholar] [CrossRef]
- Herman, A.B.; Tsitsipatis, D.; Gorospe, M. Integrated LncRNA Function upon Genomic and Epigenomic Regulation. Mol. Cell 2022, 82, 2252–2266. [Google Scholar] [CrossRef]
- Núñez-Martínez, H.N.; Recillas-Targa, F. Emerging Functions of LncRNA Loci beyond the Transcript Itself. Int. J. Mol. Sci. 2022, 23, 6258. [Google Scholar] [CrossRef]
- Sartorelli, V.; Lauberth, S.M. Enhancer RNAs Are an Important Regulatory Layer of the Epigenome. Nat. Struct. Mol. Biol. 2020, 27, 521–528. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene Regulation by Long Non-Coding RNAs and Its Biological Functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyrchanova, O.; Sokolov, V.; Tikhonov, M.; Manukyan, G.; Schedl, P.; Georgiev, P. Transcriptional Readthrough Interrupts Boundary Function in Drosophila. Int. J. Mol. Sci. 2023, 24, 11368. https://doi.org/10.3390/ijms241411368
Kyrchanova O, Sokolov V, Tikhonov M, Manukyan G, Schedl P, Georgiev P. Transcriptional Readthrough Interrupts Boundary Function in Drosophila. International Journal of Molecular Sciences. 2023; 24(14):11368. https://doi.org/10.3390/ijms241411368
Chicago/Turabian StyleKyrchanova, Olga, Vladimir Sokolov, Maxim Tikhonov, Galya Manukyan, Paul Schedl, and Pavel Georgiev. 2023. "Transcriptional Readthrough Interrupts Boundary Function in Drosophila" International Journal of Molecular Sciences 24, no. 14: 11368. https://doi.org/10.3390/ijms241411368