

Impact of Truncated Oxidized Phosphatidylcholines on Phospholipase A2 Activity in Mono- and Polyunsaturated Biomimetic Vesicles
 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
2.1. PLA2 Activity Assay
2.1.1. Effect of the Unsaturated Fatty Acid at sn-2 Position on sPLA2 Activity in PC Vesicles
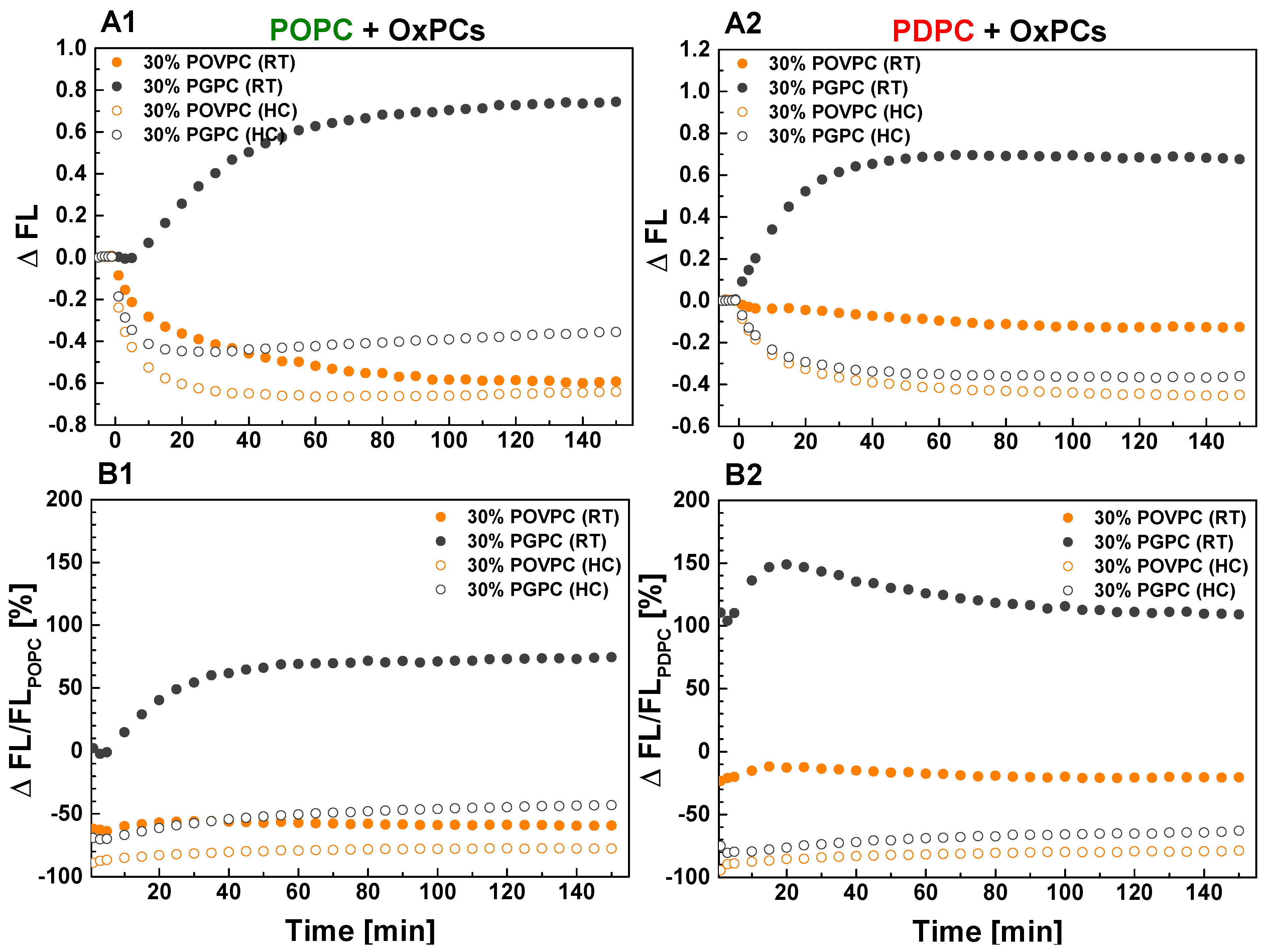
2.1.2. Effect on sPLA2 Activity on POPC and PDPC Vesicles after POVPC or PGPC Addition
2.2. Effect of POVPC and PGPC on Membrane Lipid Order in POPC or PDPC Lipid Matrix
2.3. Effect of POVPC and PGPC on Vesicle Size in POPC and PDPC Lipid Matrix
3. Materials and Methods
3.1. Materials
3.2. Liposome Preparation
3.3. Fluorescence Spectroscopy
3.4. Dynamic Light Scattering (DLS) Measurements
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schaloske, R.H.; Dennis, E.A. The phospholipase A2 superfamily and its group numbering system. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2006, 1761, 1246–1259. [Google Scholar] [CrossRef] [PubMed]
- Brueseke, T.J.; Bell, J.D. A new hat for an old enzyme: Waste management. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2006, 1761, 1270–1279. [Google Scholar] [CrossRef] [PubMed]
- Geddis, M.S.; Rehder, V. Initial stages of neural regeneration in Helisoma trivolvis are dependent upon PLA2 activity. J. Neurobiol. 2003, 54, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Dennis, E.A. Phospholipase A2 in eicosanoid generation. Am. J. Respir. Crit. Care Med. 2000, 161, S32–S35. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Kudo, I. Phospholipase A2. J. Biochem. 2002, 131, 285–292. [Google Scholar] [CrossRef]
- Yedgar, S.; Cohen, Y.; Shoseyov, D. Control of phospholipase A2 activities for the treatment of inflammatory conditions. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2006, 1761, 1373–1382. [Google Scholar] [CrossRef]
- Karakas, M.; Koenig, W. Phospholipase A2 as a therapeutic target for atherosclerosis. Clin. Lipidol. 2010, 5, 43–56. [Google Scholar] [CrossRef]
- Burke, J.E.; Dennis, E.A. Phospholipase A2 structure/function, mechanism, and signaling1. J. Lipid Res. 2009, 50, S237–S242. [Google Scholar] [CrossRef] [Green Version]
- Aloulou, A.; Ali, Y.; Sofiane, B.; Gargouri, Y.; Gelb, M. Phospholipases: An Overview. Methods Mol. Biol. 2012, 861, 63–85. [Google Scholar] [CrossRef]
- Dennis, E.A.; Cao, J.; Hsu, Y.-H.; Magrioti, V.; Kokotos, G. Phospholipase A2 Enzymes: Physical Structure, Biological Function, Disease Implication, Chemical Inhibition, and Therapeutic Intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef] [Green Version]
- Hønger, T.; Jørgensen, K.; Stokes, D.; Biltonen, R.L.; Mouritsen, O.G. Phospholipase A2 activity and physical properties of lipid-bilayer substrates. In Methods in Enzymology; Academic Press: New York, NY, USA, 1997; Volume 286, pp. 168–190. [Google Scholar]
- Alekseeva, A.S.; Volynsky, P.E.; Krylov, N.A.; Chernikov, V.P.; Vodovozova, E.L.; Boldyrev, I.A. Phospholipase A2 way to hydrolysis: Dint formation, hydrophobic mismatch, and lipid exclusion. Biochim. Biophys. Acta (BBA)-Biomembr. 2021, 1863, 183481. [Google Scholar] [CrossRef]
- Høyrup, P.; Jørgensen, K.; Mouritsen, O.G. Phospholipase A2—An enzyme that is sensitive to the physics of its substrate. Europhys. Lett. (EPL) 2002, 57, 464–470. [Google Scholar] [CrossRef]
- Bell, J.D.; Biltonen, R.L. Activation of phospholipase A2 on lipid bilayers. In Methods in Enzymology; Academic Press: New York, NY, USA, 1991; Volume 197, pp. 249–258. [Google Scholar]
- Zumbuehl, A.; Dobner, B.; Brezesinski, G. Phase behavior of selected artificial lipids. Curr. Opin. Colloid Interface Sci. 2014, 19, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Hjort Ipsen, J.; Karlström, G.; Mourtisen, O.G.; Wennerström, H.; Zuckermann, M.J. Phase equilibria in the phosphatidylcholine-cholesterol system. Biochim. Biophys. Acta (BBA)-Biomembr. 1987, 905, 162–172. [Google Scholar] [CrossRef]
- Feigenson, G.W. Phase behavior of lipid mixtures. Nat. Chem. Biol. 2006, 2, 560–563. [Google Scholar] [CrossRef]
- Veatch, S.L.; Keller, S.L. Separation of Liquid Phases in Giant Vesicles of Ternary Mixtures of Phospholipids and Cholesterol. Biophys. J. 2003, 85, 3074–3083. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, H.-J.; Lingwood, D.; Levental, I.; Sampaio, J.L.; Kalvodova, L.; Rajendran, L.; Simons, K. Order of lipid phases in model and plasma membranes. Proc. Natl. Acad. Sci. USA 2009, 106, 16645–16650. [Google Scholar] [CrossRef]
- Hashimoto, M.; Hossain, S.; Al Mamun, A.; Matsuzaki, K.; Arai, H. Docosahexaenoic acid: One molecule diverse functions. Crit. Rev. Biotechnol. 2017, 37, 579–597. [Google Scholar] [CrossRef] [Green Version]
- Layé, S. Polyunsaturated fatty acids, neuroinflammation and well being. Prostaglandins Leukot. Essent. Fat. Acids (PLEFA) 2010, 82, 295–303. [Google Scholar] [CrossRef]
- Saito, M.; Nakatsugawa, K. Increased susceptibility of liver to lipid-peroxidation after ingestion of a high fish-oil diet. Int. J. Vitam. Nutr. Res. 1994, 64, 144–151. [Google Scholar]
- Bochkov, V.; Oskolkova, O.; Birukov, K.; Levonen, A.-L.; Binder, C.; Stöckl, J. Generation and Biological Activities of Oxidized Phospholipids. Antioxid. Redox Signal. 2009, 12, 1009–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashraf, M.Z.; Srivastava, S. Oxidized Phospholipids: Introduction and Biological Significance. In Lipoproteins—Role in Health and Diseases; Frank, S., Kostner, G., Eds.; BoD—Books on Demand; InTech: Rijeka, Croatia, 2012; pp. 409–430. [Google Scholar] [CrossRef] [Green Version]
- Nasri, Z.; Memari, S.; Wenske, S.; Clemen, R.; Martens, U.; Delcea, M.; Bekeschus, S.; Weltmann, K.-D.; von Woedtke, T.; Wende, K. Singlet-Oxygen-Induced Phospholipase A2 Inhibition: A Major Role for Interfacial Tryptophan Dioxidation. Chem.–A Eur. J. 2021, 27, 14702–14710. [Google Scholar] [CrossRef] [PubMed]
- Deigner, H.-P.; Hermetter, A. Oxidized phospholipids: Emerging lipid mediators in pathophysiology. Curr. Opin. Lipidol. 2008, 19, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Fruhwirth, G.O.; Loidl, A.; Hermetter, A. Oxidized phospholipids: From molecular properties to disease. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2007, 1772, 718–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, A.D.; Leitinger, N.; Navab, M.; Faull, K.F.; Hörkkö, S.; Witztum, J.L.; Palinski, W.; Schwenke, D.; Salomon, R.G.; Sha, W.; et al. Structural Identification by Mass Spectrometry of Oxidized Phospholipids in Minimally Oxidized Low Density Lipoprotein That Induce Monocyte/Endothelial Interactions and Evidence for Their Presence in Vivo. J. Biol. Chem. 1997, 272, 13597–13607. [Google Scholar] [CrossRef] [Green Version]
- Shancy Petsel, J.; Chikkamenahalli Lakshminarayana, L.; Thomas, M.M.; Gopal Kedihitlu, M. Platelet-activating factor and oxidized phosphatidylcholines do not suppress endotoxin-induced pro-inflammatory signaling among human myeloid and endothelial cells. AIMS Allergy Immunol. 2017, 1, 108–123. [Google Scholar] [CrossRef]
- Reddy, S.; Hama, S.; Grijalva, V.; Hassan, K.; Mottahedeh, R.; Hough, G.; Wadleigh, D.J.; Navab, M.; Fogelman, A.M. Mitogen-activated Protein Kinase Phosphatase 1 Activity Is Necessary for Oxidized Phospholipids to Induce Monocyte Chemotactic Activity in Human Aortic Endothelial Cells*. J. Biol. Chem. 2001, 276, 17030–17035. [Google Scholar] [CrossRef] [Green Version]
- Bochkov, V.N.; Kadl, A.; Huber, J.; Gruber, F.; Binder, B.R.; Leitinger, N. Protective role of phospholipid oxidation products in endotoxin-induced tissue damage. Nature 2002, 419, 77–81. [Google Scholar] [CrossRef]
- Hartwich, J.; Dembinska-Kieć, A.; Gruca, A.; Motyka, M.; Partyka, L.; Skrzeczyńska, J.; Bzowska, M.; Pryjma, J.; Huber, J.; Leitinger, N.; et al. Regulation of platelet adhesion by oxidized lipoproteins and oxidized phospholipids. Platelets 2002, 13, 141–151. [Google Scholar] [CrossRef]
- Loidl, A.; Sevcsik, E.; Riesenhuber, G.; Deigner, H.-P.; Hermetter, A. Oxidized Phospholipids in Minimally Modified Low Density Lipoprotein Induce Apoptotic Signaling via Activation of Acid Sphingomyelinase in Arterial Smooth Muscle Cells*. J. Biol. Chem. 2003, 278, 32921–32928. [Google Scholar] [CrossRef] [Green Version]
- Walton, K.A.; Cole, A.L.; Yeh, M.; Subbanagounder, G.; Krutzik, S.R.; Modlin, R.L.; Lucas, R.M.; Nakai, J.; Smart, E.J.; Vora, D.K.; et al. Specific Phospholipid Oxidation Products Inhibit Ligand Activation of Toll-Like Receptors 4 and 2. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1197–1203. [Google Scholar] [CrossRef] [Green Version]
- Stemmer, U.; Hermetter, A. Protein modification by aldehydophospholipids and its functional consequences. Biochim. Biophys. Acta (BBA)-Biomembr. 2012, 1818, 2436–2445. [Google Scholar] [CrossRef] [Green Version]
- Stemmer, U.; Ramprecht, C.; Zenzmaier, E.; Stojčić, B.; Rechberger, G.; Kollroser, M.; Hermetter, A. Uptake and protein targeting of fluorescent oxidized phospholipids in cultured RAW 264.7 macrophages. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2012, 1821, 706–718. [Google Scholar] [CrossRef] [Green Version]
- Friedman, P.; Hörkkö, S.; Steinberg, D.; Witztum, J.L.; Dennis, E.A. Correlation of Antiphospholipid Antibody Recognition with the Structure of Synthetic Oxidized Phospholipids: Importance of schiff base formation and aldol condensation*. J. Biol. Chem. 2002, 277, 7010–7020. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Gutteridge, J.M.C. Role of free radicals and catalytic metal ions in human disease: An overview. In Methods in Enzymology; Academic Press: New York, NY, USA, 1990; Volume 186, pp. 1–85. [Google Scholar]
- Bhatia, S. Introduction to enzymes and their applications. In Introduction to Pharmaceutical Biotechnology; IOP Publishing: Bristol, England, 2018; Volume 2. [Google Scholar] [CrossRef]
- Bisswanger, H. Enzyme assays. Perspect. Sci. 2014, 1, 41–55. [Google Scholar] [CrossRef] [Green Version]
- Parasassi, T.; Krasnowska, E.K.; Bagatolli, L.; Gratton, E. Laurdan and Prodan as Polarity-Sensitive Fluorescent Membrane Probes. J. Fluoresc. 1998, 8, 365–373. [Google Scholar] [CrossRef]
- Parasassi, T.; Gilbert, P.; Ravagnan, G.; Rusch, R.; Gratton, E. Quantitation of lipid phases in phospholipid vesicles by the generalized polarization of Laurdan fluorescence. Biophys. J. 1991, 60, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Lasch, J.; Willhardt, I.; Kinder, D.; Sauer, H.; Smesny, S. Fluorometric Assays of Phospholipase A2 Activity with Three Different Substrates in Biological Samples of Patients with Schizophrenia. Clin. Chem. Lab. Med. (CCLM) 2003, 41, 908–914. [Google Scholar] [CrossRef]
- Cho, W. Seeing Is Believing: Real-Time Cellular Activity Assay for Phospholipase A2. ACS Chem. Biol. 2006, 1, 65–66. [Google Scholar] [CrossRef] [Green Version]
- Vitkova, V.; Staneva, G.; Hazarosova, R.; Georgieva, S.I.; Valkova, I.; Antonova, K.; Todorov, P. Valorphins alter physicochemical characteristics of phosphatidylcholine membranes: Datasets on lipid packing, bending rigidity, specific electrical capacitance, dipole potential, vesicle size. Data Brief 2022, 45, 108716. [Google Scholar] [CrossRef]
- Pande, A.H.; Kar, S.; Tripathy, R.K. Oxidatively modified fatty acyl chain determines physicochemical properties of aggregates of oxidized phospholipids. Biochim. Biophys. Acta (BBA)-Biomembr. 2010, 1798, 442–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattila, J.-P.; Sabatini, K.; Kinnunen, P.K.J. Oxidized Phospholipids as Potential Novel Drug Targets. Biophys. J. 2007, 93, 3105–3112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, P.; Birukov, K.G. Oxidized phospholipids in control of inflammation and endothelial barrier. Transl. Res. 2009, 153, 166–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Holub, B.J. Clinical nutrition: 4. Omega-3 fatty acids in cardiovascular care. Can. Med. Assoc. J. 2002, 166, 608–615. [Google Scholar]
- Stillwell, W.; Wassall, S.R. Docosahexaenoic acid: Membrane properties of a unique fatty acid. Chem. Phys. Lipids 2003, 126, 1–27. [Google Scholar] [CrossRef]
- De Santis, A.; Varela, Y.; Sot, J.; D’Errico, G.; Goñi, F.M.; Alonso, A. Omega-3 polyunsaturated fatty acids do not fluidify bilayers in the liquid-crystalline state. Sci. Rep. 2018, 8, 16240. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Davis, T.A.; Porter, N.A. Rate Constants for Peroxidation of Polyunsaturated Fatty Acids and Sterols in Solution and in Liposomes. J. Am. Chem. Soc. 2009, 131, 13037–13044. [Google Scholar] [CrossRef] [Green Version]
- Reis, A.; Spickett, C.M. Chemistry of phospholipid oxidation. Biochim. Et Biophys. Acta (BBA) - Biomembr. 2012, 1818, 2374–2387. [Google Scholar] [CrossRef] [Green Version]
- Beranova, L.; Cwiklik, L.; Jurkiewicz, P.; Hof, M.; Jungwirth, P. Oxidation Changes Physical Properties of Phospholipid Bilayers: Fluorescence Spectroscopy and Molecular Simulations. Langmuir 2010, 26, 6140–6144. [Google Scholar] [CrossRef]
- Singer, A.G.; Ghomashchi, F.; Le Calvez, C.; Bollinger, J.; Bezzine, S.; Rouault, M.; Sadilek, M.; Nguyen, E.; Lazdunski, M.; Lambeau, G.; et al. Interfacial Kinetic and Binding Properties of the Complete Set of Human and Mouse Groups I, II, V, X, and XII Secreted Phospholipases A2. J. Biol. Chem. 2002, 277, 48535–48549. [Google Scholar] [CrossRef] [Green Version]
- Hossen, M.S.; Shapla, U.M.; Gan, S.H.; Khalil, M.I. Impact of Bee Venom Enzymes on Diseases and Immune Responses. Molecules 2017, 22, 25. [Google Scholar] [CrossRef] [Green Version]
- Bollinger, J.G.; Diraviyam, K.; Ghomashchi, F.; Murray, D.; Gelb, M.H. Interfacial Binding of Bee Venom Secreted Phospholipase A2 to Membranes Occurs Predominantly by a Nonelectrostatic Mechanism. Biochemistry 2004, 43, 13293–13304. [Google Scholar] [CrossRef]
- Ghomashchi, F.; Lin, Y.; Hixon, M.S.; Yu, B.-Z.; Annand, R.; Jain, M.K.; Gelb, M.H. Interfacial Recognition by Bee Venom Phospholipase A2: Insights into Nonelectrostatic Molecular Determinants by Charge Reversal Mutagenesis. Biochemistry 1998, 37, 6697–6710. [Google Scholar] [CrossRef]
- Pande, A.H.; Qin, S.; Nemec, K.N.; He, X.; Tatulian, S.A. Isoform-Specific Membrane Insertion of Secretory Phospholipase A2 and Functional Implications. Biochemistry 2006, 45, 12436–12447. [Google Scholar] [CrossRef]
- Mouchlis, V.D.; Dennis, E.A. Phospholipase A2 catalysis and lipid mediator lipidomics. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2019, 1864, 766–771. [Google Scholar] [CrossRef]
- Abe, A.; Hiraoka, M.; Ohguro, H.; Tesmer, J.J.; Shayman, J.A. Preferential hydrolysis of truncated oxidized glycerophospholipids by lysosomal phospholipase A2. J. Lipid Res. 2017, 58, 339–349. [Google Scholar] [CrossRef] [Green Version]
- Koumanov, K.; Wolf, C.; Béreziat, G. Modulation of human type II secretory phospholipase A2 by sphingomyelin and annexin VI. Biochem. J. 1997, 326, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Gesquiere, L.; Cho, W.; Subbaiah, P.V. Role of Group IIa and Group V Secretory Phospholipases A2 in the Metabolism of Lipoproteins. Substrate Specificities of the Enzymes and the Regulation of Their Activities by Sphingomyelin. Biochemistry 2002, 41, 4911–4920. [Google Scholar] [CrossRef]
- Singh, D.K.; Subbaiah, P.V. Modulation of the activity and arachidonic acid selectivity of group X secretory phospholipase A2 by sphingolipids. J. Lipid Res. 2007, 48, 683–692. [Google Scholar] [CrossRef] [Green Version]
- Subbaiah, P.V.; Sargis, R.M. Sphingomyelin: A natural modulator of membrane homeostasis and inflammation. Med. Hypotheses 2001, 57, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.N.; Agarwal, S. Liposomes as membrane model for study of lipid peroxidation. Free Radic. Biol. Med. 1988, 4, 51–72. [Google Scholar] [CrossRef] [PubMed]
- Feller, S.E.; Gawrisch, K.; MacKerell, A.D. Polyunsaturated Fatty Acids in Lipid Bilayers: Intrinsic and Environmental Contributions to Their Unique Physical Properties. J. Am. Chem. Soc. 2002, 124, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Eldho, N.V.; Feller, S.E.; Tristram-Nagle, S.; Polozov, I.V.; Gawrisch, K. Polyunsaturated Docosahexaenoic vs Docosapentaenoic AcidDifferences in Lipid Matrix Properties from the Loss of One Double Bond. J. Am. Chem. Soc. 2003, 125, 6409–6421. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Ranganathan, R. Mixing of oxidized and bilayer phospholipids. Biochim. Biophys. Acta (BBA)-Biomembr. 2015, 1848, 1472–1480. [Google Scholar] [CrossRef] [Green Version]
- Ramprecht, C.; Jaritz, H.; Streith, I.; Zenzmaier, E.; Köfeler, H.; Hofmann-Wellenhof, R.; Schaider, H.; Hermetter, A. Toxicity of oxidized phosphatidylcholines in cultured human melanoma cells. Chem. Phys. Lipids 2015, 189, 39–47. [Google Scholar] [CrossRef]
- Imai, Y.; Kuba, K.; Neely, G.G.; Yaghubian-Malhami, R.; Perkmann, T.; van Loo, G.; Ermolaeva, M.; Veldhuizen, R.; Leung, Y.H.C.; Wang, H.; et al. Identification of Oxidative Stress and Toll-like Receptor 4 Signaling as a Key Pathway of Acute Lung Injury. Cell 2008, 133, 235–249. [Google Scholar] [CrossRef]
- Stewart, C.R.; Stuart, L.M.; Wilkinson, K.; van Gils, J.M.; Deng, J.; Halle, A.; Rayner, K.J.; Boyer, L.; Zhong, R.; Frazier, W.A.; et al. CD36 ligands promote sterile inflammation through assembly of a Toll-like receptor 4 and 6 heterodimer. Nat. Immunol. 2010, 11, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Podrez, E.A.; Poliakov, E.; Shen, Z.; Zhang, R.; Deng, Y.; Sun, M.; Finton, P.J.; Shan, L.; Febbraio, M.; Hajjar, D.P.; et al. A Novel Family of Atherogenic Oxidized Phospholipids Promotes Macrophage Foam Cell Formation via the Scavenger Receptor CD36 and Is Enriched in Atherosclerotic Lesions*. J. Biol. Chem. 2002, 277, 38517–38523. [Google Scholar] [CrossRef] [Green Version]
- Binder, C.J.; Hörkkö, S.; Dewan, A.; Chang, M.-K.; Kieu, E.P.; Goodyear, C.S.; Shaw, P.X.; Palinski, W.; Witztum, J.L.; Silverman, G.J. Pneumococcal vaccination decreases atherosclerotic lesion formation: Molecular mimicry between Streptococcus pneumoniae and oxidized LDL. Nat. Med. 2003, 9, 736–743. [Google Scholar] [CrossRef]
- Bretscher, P.; Egger, J.; Shamshiev, A.; Trötzmüller, M.; Köfeler, H.; Carreira, E.M.; Kopf, M.; Freigang, S. Phospholipid oxidation generates potent anti-inflammatory lipid mediators that mimic structurally related pro-resolving eicosanoids by activating Nrf2. EMBO Mol. Med. 2015, 7, 593–607. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ΔFL/FLPC Values of sPLA2 Hydrolysis (%) at Steady State | |||
|---|---|---|---|
| POPC+ | ΔFL/FL | PDPC+ | ΔFL/FL |
| 30% PGPC (RT) | +74.4 | 30% PGPC (RT) | +109.1 |
| 30% PGPC (HC) | −43.1 | 30% PGPC (HC) | −63.0 |
| 30% POVPC (RT) | −59.3 | 30% POVPC (RT) | −20.3 |
| 30% POVPC (HC) | −77.7 | 30% POVPC (HC) | −78.8 |
| Composition | FL = F530/F530, initial − 1 at t = 0 min; a | PLA2 Reaction Rate, b, min−1 |
|---|---|---|
| POPC (RT) | 0.073 ± 0.007 | 0.060 ± 0.003 |
| POPC + 30% POVPC (RT) | 0.026 ± 0.003 | 0.023 ± 0.002 |
| POPC + 30% PGPC (RT) | 0.085 ± 0.003 | 0.053 ± 0.002 |
| POPC (HC) | 0.179 ± 0.012 | 0.080 ± 0.007 |
| POPC + 30% POVPC (HC) | 0.015 ± 0.001 | 0.012 ± 7.79 × 10−4 |
| POPC + 30% PGPC (HC) | 0.059 ± 0.003 | 0.021 ± 0.002 |
| PDPC (RT) | 0.046 ± 0.004 | 0.033 ± 0.002 |
| PDPC + 30% POVPC (RT) | 0.036 ± 0.001 | 0.026 ± 8.55 × 10−4 |
| PDPC + 30% PGPC (RT) | 0.106 ± 0.004 | 0.063 ± 0.003 |
| PDPC (HC) | 0.049 ± 0.003 | 0.039 ± 0.002 |
| PDPC + 30% POVPC (HC) | 0.000 ± 0.001 | 0.007 ± 8.61 × 10−4 |
| PDPC + 30% PGPC (HC) | 0.017 ± 0.001 | 0.005 ± 6.63 × 10−4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yordanova, V.; Hazarosova, R.; Vitkova, V.; Momchilova, A.; Robev, B.; Nikolova, B.; Krastev, P.; Nuss, P.; Angelova, M.I.; Staneva, G. Impact of Truncated Oxidized Phosphatidylcholines on Phospholipase A2 Activity in Mono- and Polyunsaturated Biomimetic Vesicles. Int. J. Mol. Sci. 2023, 24, 11166. https://doi.org/10.3390/ijms241311166
Yordanova V, Hazarosova R, Vitkova V, Momchilova A, Robev B, Nikolova B, Krastev P, Nuss P, Angelova MI, Staneva G. Impact of Truncated Oxidized Phosphatidylcholines on Phospholipase A2 Activity in Mono- and Polyunsaturated Biomimetic Vesicles. International Journal of Molecular Sciences. 2023; 24(13):11166. https://doi.org/10.3390/ijms241311166
Chicago/Turabian StyleYordanova, Vesela, Rusina Hazarosova, Victoria Vitkova, Albena Momchilova, Bozhil Robev, Biliana Nikolova, Plamen Krastev, Philippe Nuss, Miglena I. Angelova, and Galya Staneva. 2023. "Impact of Truncated Oxidized Phosphatidylcholines on Phospholipase A2 Activity in Mono- and Polyunsaturated Biomimetic Vesicles" International Journal of Molecular Sciences 24, no. 13: 11166. https://doi.org/10.3390/ijms241311166