
Genome-Wide Identification and Expression Analysis of ACTIN Family Genes in the Sweet Potato and Its Two Diploid Relatives

Abstract
:1. Introduction
2. Results
2.1. Identification and Characteristic of ACTINs in the Sweet Potato and Its Two Diploid Relatives
2.2. Phylogenetic Relationships of ACTINs in the Sweet Potato and Its Two Diploid Relatives
2.3. Analysis of Conserved Motifs and Exon-Intron Structure of ACTINs in the Sweet Potato and Its Two Diploid Relatives
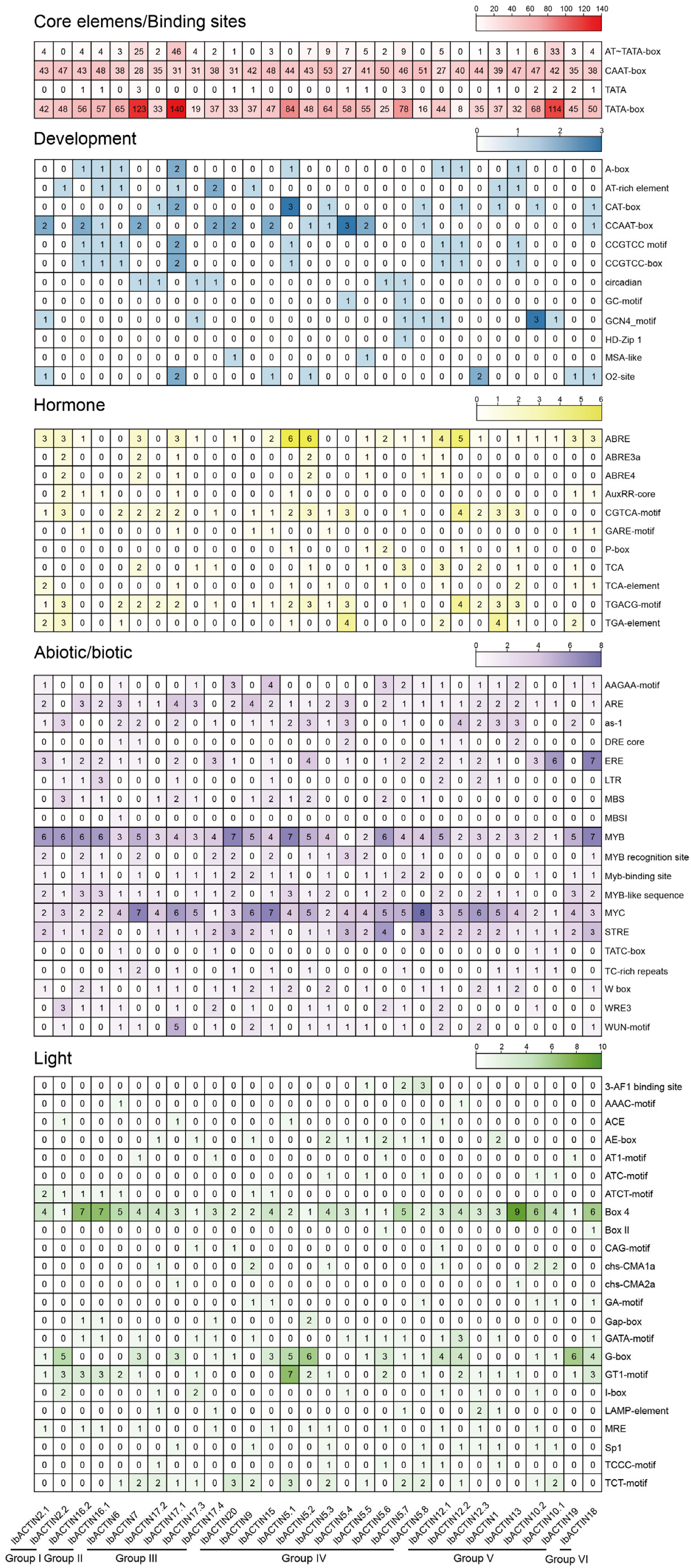
2.4. Analysis of cis-Elements in the Promoter of IbACTINs in the Sweet Potato
2.5. Protein Interaction Network of ACTINs in the Sweet Potato
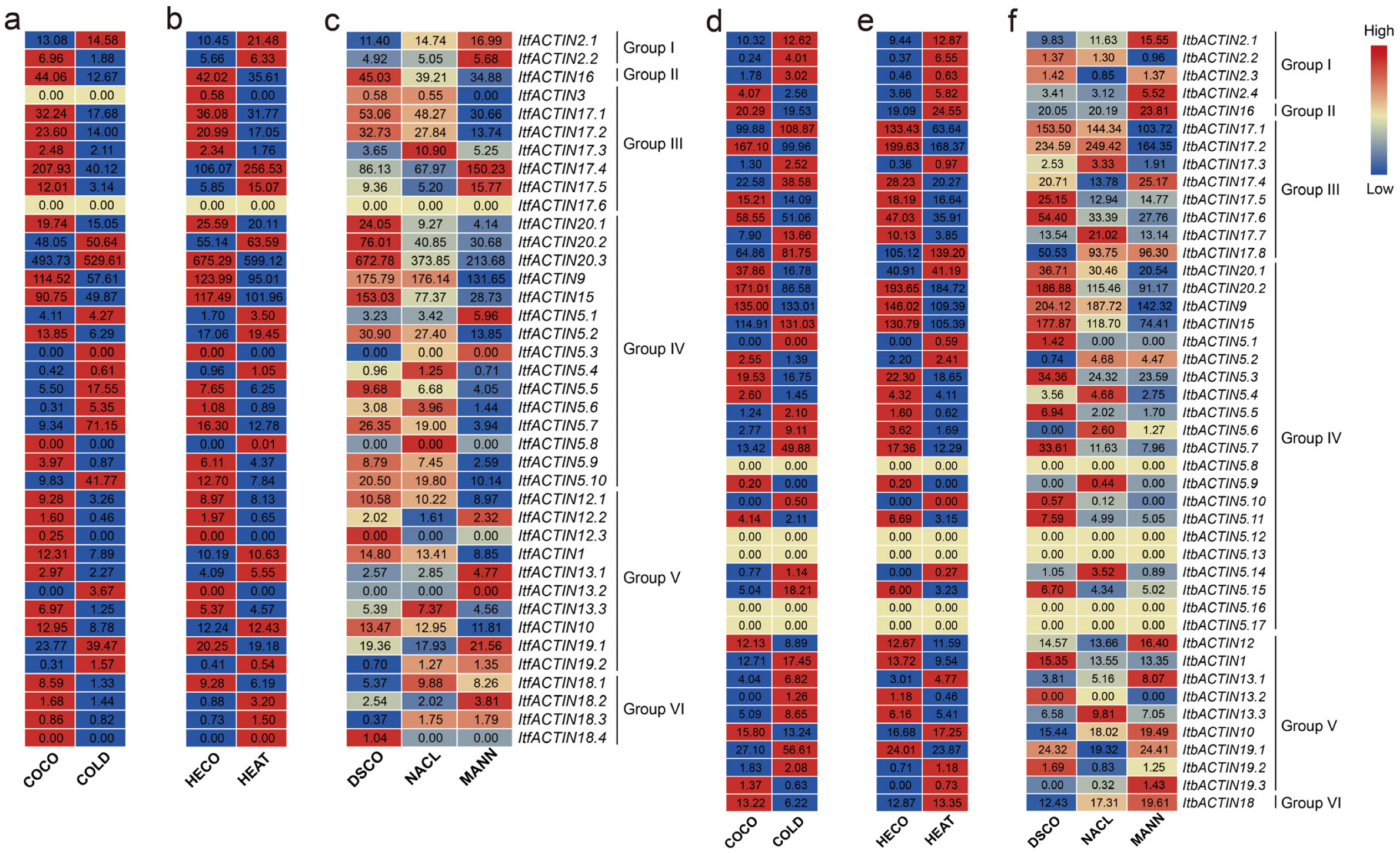
2.6. Expression Analysis of ACTINs in the Sweet Potato and Its Two Diploid Relatives
2.6.1. Analysis of Expression in Various Tissues
2.6.2. Analysis of Expression in Different Developmental Stages
2.6.3. Analysis of the Expression of Hormone Response
2.6.4. Analysis of Expression under Abiotic Stresses
3. Discussion
3.1. Evolution of the ACTIN Gene Family in the Sweet Potato and Its Two Diploid Relatives
3.2. Stable Expression of ACTIN in the Sweet Potato and Its Two Diploid Relatives
3.3. Differences in the Functions of ACTINs in Growth and Development between the Sweet Potato and Its Two Diploid Relatives
3.4. Different Functions of ACTINs in Hormone Crosstalk in the Sweet Potato and Its Two Diploid Relatives
3.5. Different Functions of ACTINs in Abiotic Stress Responses in the Sweet Potato and Its Two Diploid Relatives
4. Materials and Methods
4.1. Identification of ACTINs
4.2. Chromosomal Distribution of ACTINs
4.3. Protein Properties Prediction of ACTINs
4.4. Phylogenetic Analysis of ACTINs
4.5. Domain Identification and Conserved Motifs Analysis of ACTINs
4.6. Exon-Intron Structures and Promoter Analysis of ACTINs
4.7. Protein Interaction Network of ACTINs
4.8. Transcriptome Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jürgens, G. Cytokinesis in higher plants. Annu. Rev. Plant Biol. 2005, 56, 281–299. [Google Scholar] [CrossRef] [PubMed]
- Mascarenhas, J.P. Molecular mechanisms of pollen tube growth and differentiation. Plant Cell 1993, 5, 1303–1314. [Google Scholar] [CrossRef] [Green Version]
- Baluska, F.; Hlavacka, A.; Samaj, J.; Palme, K.; Robinson, D.G.; Matoh, T.; McCurdy, D.W.; Menzel, D.; Volkmann, D. F-actin-dependent endocytosis of cell wall pectins in meristematic root cells. Insights from brefeldin A-induced compartments. Plant Physiol. 2002, 130, 422–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrero, R.A.; Umeda, M.; Yamamura, S.; Uchimiya, H. Arabidopsis CAP regulates the actin cytoskeleton necessary for plant cell elongation and division. Plant Cell 2002, 14, 149–163. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Fu, Y.; Dowd, P.; Li, S.; Vernoud, V.; Gilroy, S.; Yang, Z. A Rho family GTPase controls actin dynamics and tip growth via two counteracting downstream pathways in pollen tubes. J. Cell Biol. 2005, 169, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Maisch, J.; Nick, P. Actin is involved in auxin-dependent patterning. Plant Physiol. 2007, 143, 1695–1704. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, R.; Holmes, K.C. Actin structure and function. Annu. Rev. Biophys. 2011, 40, 169–186. [Google Scholar] [CrossRef] [Green Version]
- Kabsch, W.; Mannherz, H.G.; Suck, D.; Pai, E.F.; Holmes, K.C. Atomic structure of the actin: DNase I complex. Nature 1990, 347, 37–44. [Google Scholar] [CrossRef]
- Meagher, R.B.; McKinney, E.C.; Kandasamy, M.K. Isovariant dynamics expand and buffer the responses of complex systems: The diverse plant actin gene family. Plant Cell 1999, 11, 995–1006. [Google Scholar] [CrossRef] [Green Version]
- Shah, D.M.; Hightower, R.C.; Meagher, R.B. Genes encoding actin in higher plants: Intron positions are highly conserved but the coding sequences are not. J. Mol. Appl. Genet. 1983, 2, 111–126. [Google Scholar]
- Hanson, J.; Lowy, J. The structure of F-actin and of actin filaments isolated from muscle. J. Mol. Biol. 1963, 6, 46–60. [Google Scholar] [CrossRef]
- Franklin-Tong, V.E. Signaling and the modulation of pollen tube growth. Plant Cell 1999, 11, 727–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramanian, R.; Karve, A.; Kandasamy, M.; Meagher, R.B.; Moore, B. A role for F-actin in hexokinase-mediated glucose signaling. Plant Physiol. 2007, 145, 1423–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmieri, M.; Kiss, J.Z. Disruption of the F-actin cytoskeleton limits statolith movement in Arabidopsis hypocotyls. J. Exp. Bot. 2005, 56, 2539–2550. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Chen, Q.; Ji, Q.; Zhang, L.; Wang, J. Genome-wide analysis of ACTIN families in Arabidopsis Thaliana, Oryza Sativa and Populus trichocarpa. J. Shanghai Univ. 2009, 15, 426–431. [Google Scholar]
- Cui, L.; Zheng, T.; Zhang, K.; Zhang, C.; Shang, G.; Fang, J. Identification, evolution and expression analyses of Actin gene family of Vitis vinifera. J. Plant Resour. Environ. 2017, 15, 1–10. [Google Scholar]
- McDowell, J.M.; Huang, S.; McKinney, E.C.; An, Y.Q.; Meagher, R.B. Structure and evolution of the actin gene family in Arabidopsis thaliana. Genetics 1996, 142, 587–602. [Google Scholar] [CrossRef]
- Huang, S.; An, Y.Q.; McDowell, J.M.; McKinney, E.C.; Meagher, R.B. The Arabidopsis ACT11 actin gene is strongly expressed in tissues of the emerging inflorescence, pollen, and developing ovules. Plant Mol. Biol. 1997, 33, 125–139. [Google Scholar] [CrossRef]
- Huang, S.; An, Y.Q.; McDowell, J.M.; McKinney, E.C.; Meagher, R.B. The Arabidopsis thaliana ACT4/ACT12 actin gene subclass is strongly expressed throughout pollen development. Plant J. Cell Mol. Biol. 1996, 10, 189–202. [Google Scholar] [CrossRef]
- An, Y.Q.; Huang, S.; McDowell, J.M.; McKinney, E.C.; Meagher, R.B. Conserved expression of the Arabidopsis ACT1 and ACT3 actin subclass in organ primordia and mature pollen. Plant Cell 1996, 8, 15–30. [Google Scholar] [CrossRef]
- Kandasamy, M.K.; McKinney, E.C.; Meagher, R.B. Functional nonequivalency of actin isovariants in Arabidopsis. Mol. Biol. Cell 2002, 13, 251–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inada, N.; Takahashi, N.; Umeda, M. Arabidopsis thaliana subclass I ACTIN DEPOLYMERIZING FACTORs and vegetative ACTIN2/8 are novel regulators of endoreplication. J. Plant Res. 2021, 134, 1291–1300. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, M.K.; McKinney, E.C.; Meagher, R.B. A single vegetative actin isovariant overexpressed under the control of multiple regulatory sequences is sufficient for normal Arabidopsis development. Plant Cell 2009, 21, 701–718. [Google Scholar] [CrossRef] [Green Version]
- McElroy, D.; Rothenberg, M.; Reece, K.S.; Wu, R. Characterization of the rice (Oryza sativa) actin gene family. Plant Mol. Biol. 1990, 15, 257–268. [Google Scholar] [CrossRef]
- Brunner, A.M.; Yakovlev, I.A.; Strauss, S.H. Validating internal controls for quantitative plant gene expression studies. BMC Plant Biol. 2004, 4, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.-R.; Nam, H.-Y.; Kim, S.-U.; Kim, S.-I.; Chang, Y.-J. Normalization of reverse transcription quantitative-PCR with housekeeping genes in rice. Biotechnol. Lett. 2003, 25, 1869–1872. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Nijhawan, A.; Tyagi, A.K.; Khurana, J.P. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2006, 345, 646–651. [Google Scholar] [CrossRef]
- Jian, B.; Liu, B.; Bi, Y.; Hou, W.; Wu, C.; Han, T. Validation of internal control for gene expression study in soybean by quantitative real-time PCR. BMC Mol. Biol. 2008, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q. Improvement for agronomically important traits by gene engineering in sweetpotato. Breed. Sci. 2017, 67, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Wang, Z.; Li, X.; He, S.; Liu, Q.; Zhai, H.; Zhao, N.; Gao, S.; Zhang, H. Genome-Wide Identification and Expression Analysis of JAZ Family Involved in Hormone and Abiotic Stress in Sweet Potato and Its Two Diploid Relatives. Int. J. Mol. Sci. 2021, 22, 9786. [Google Scholar] [CrossRef]
- Zang, N.; Zhai, H.; Gao, S.; Chen, W.; He, S.; Liu, Q. Efficient production of transgenic plants using the bar gene for herbicide resistance in sweetpotato. Sci. Hortic. 2009, 122, 649–653. [Google Scholar] [CrossRef]
- Liu, D.; He, S.; Song, X.; Zhai, H.; Liu, N.; Zhang, D.; Ren, Z.; Liu, Q. IbSIMT1, a novel salt-induced methyltransferase gene from Ipomoea batatas, is involved in salt tolerance. Plant Cell Tissue Organ Cult. 2015, 120, 701–715. [Google Scholar] [CrossRef]
- Wu, S.; Lau, K.H.; Cao, Q.; Hamilton, J.P.; Sun, H.; Zhou, C.; Eserman, L.; Gemenet, D.C.; Olukolu, B.A.; Wang, H.; et al. Genome sequences of two diploid wild relatives of cultivated sweetpotato reveal targets for genetic improvement. Nat. Commun. 2018, 9, 4580. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Lei, Z.; Song, Y.; Hai, J.; He, D. Identification and characterization of the expansin gene family in upland cotton (Gossypium hirsutum). Sci. Agric. Sin. 2019, 52, 3713–3732. [Google Scholar]
- Xu, K.; Zhang, Y.; Zhang, Y.; Wang, B.; Wang, F.; Li, H. Research advances on the F-box gene family in plants. Biol Bull. 2018, 34, 26–32. [Google Scholar]
- Szymanski, D.B. Breaking the WAVE complex: The point of Arabidopsis trichomes. Curr. Opin. Plant Biol. 2005, 8, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.H.; Kost, B.; Xia, G.; Chua, N.H. Molecular identification and characterization of the Arabidopsis AtADF1, AtADFS and AtADF6 genes. Plant Mol. Biol. 2001, 45, 517–527. [Google Scholar] [CrossRef]
- Mathur, J.; Spielhofer, P.; Kost, B.; Chua, N. The actin cytoskeleton is required to elaborate and maintain spatial patterning during trichome cell morphogenesis in Arabidopsis thaliana. Development 1999, 126, 5559–5568. [Google Scholar] [CrossRef]
- Noh, Y.S.; Amasino, R.M. PIE1, an ISWI family gene, is required for FLC activation and floral repression in Arabidopsis. Plant Cell 2003, 15, 1671–1682. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Wang, D.; Wu, B.; Yan, F.; Xue, H.; Wang, Q.; Quan, S.; Chen, Y. Taf14 recognizes a common motif in transcriptional machineries and facilitates their clustering by phase separation. Nat. Commun. 2020, 11, 4206. [Google Scholar] [CrossRef]
- Sacharowski, S.P.; Gratkowska, D.M.; Sarnowska, E.A.; Kondrak, P.; Jancewicz, I.; Porri, A.; Bucior, E.; Rolicka, A.T.; Franzen, R.; Kowalczyk, J.; et al. SWP73 subunits of arabidopsis SWI/SNF chromatin remodeling complexes play distinct roles in leaf and flower development. Plant Cell 2015, 27, 1889–1906. [Google Scholar] [CrossRef] [PubMed]
- Eamens, A.; Vaistij, F.E.; Jones, L. NRPD1a and NRPD1b are required to maintain post-transcriptional RNA silencing and RNA-directed DNA methylation in Arabidopsis. Plant J. Cell Mol. Biol. 2008, 55, 596–606. [Google Scholar] [CrossRef]
- Kolářová, K.; Nešpor Dadejová, M.; Loja, T.; Lochmanová, G.; Sýkorová, E.; Dvořáčková, M. Disruption of NAP1 genes in Arabidopsis thaliana suppresses the fas1 mutant phenotype, enhances genome stability and changes chromatin compaction. Plant J. Cell Mol. Biol. 2021, 106, 56–73. [Google Scholar] [CrossRef]
- Gómez-Zambrano, Á.; Crevillén, P.; Franco-Zorrilla, J.M.; López, J.A.; Moreno-Romero, J.; Roszak, P.; Santos-González, J.; Jurado, S.; Vázquez, J.; Köhler, C.; et al. Arabidopsis SWC4 binds DNA and recruits the SWR1 complex to modulate histone H2A.Z deposition at key regulatory genes. Mol. Plant 2018, 11, 815–832. [Google Scholar] [CrossRef] [Green Version]
- Mao, Y.; Pavangadkar, K.A.; Thomashow, M.F.; Triezenberg, S.J. Physical and functional interactions of Arabidopsis ADA2 transcriptional coactivator proteins with the acetyltransferase GCN5 and with the cold-induced transcription factor CBF1. Biochim. Biophys. Acta 2006, 1759, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.C.; Lin, W.C.; Cheng, W.H. Salt hypersensitive mutant 9, a nucleolar APUM23 protein, is essential for salt sensitivity in association with the ABA signaling pathway in Arabidopsis. BMC Plant Biol. 2018, 18, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, T.; Zhu, M.; Yu, J.; Han, R.; Tang, C.; Xu, T.; Liu, J.; Li, Z. RNA-Seq and iTRAQ reveal multiple pathways involved in storage root formation and development in sweet potato (Ipomoea batatas L.). BMC Plant Biol. 2019, 19, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.; Zhou, Y.; Zhai, H.; He, S.; Zhao, N.; Liu, Q. Transcriptome profiling reveals insights into the molecular mechanism of drought tolerance in sweetpotato. J. Integr. Agric. 2019, 18, 9–23. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Q.; Zhai, H.; Li, Y.; Wang, X.; Liu, Q.; He, S. Transcript profile analysis reveals important roles of jasmonic acid signalling pathway in the response of sweet potato to salt stress. Sci. Rep. 2017, 7, 40819. [Google Scholar] [CrossRef] [Green Version]
- Zhu, P.; Dong, T.; Xu, T.; Kang, H. Identification, characterisation and expression analysis of MADS-box genes in sweetpotato wild relative Ipomoea trifida. Acta Physiol. Plant. 2020, 42, 163. [Google Scholar] [CrossRef]
- Wan, R.; Liu, J.; Yang, Z.; Zhu, P.; Cao, Q.; Xu, T. Genome-wide identification, characterisation and expression profile analysis of DEAD-box family genes in sweet potato wild ancestor Ipomoea trifida under abiotic stresses. Genes Genom. 2020, 42, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhu, P.; Wu, S.; Lu, Y.; Sun, J.; Cao, Q.; Li, Z.; Xu, T. Identification and expression analysis of GRAS transcription factors in the wild relative of sweet potato Ipomoea trifida. BMC Genom. 2019, 20, 911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Sun, J.; Chen, Y.; Zhu, P.; Zhang, L.; Wu, S.; Ma, D.; Cao, Q.; Li, Z.; Xu, T. Genome-wide identification, structural and gene expression analysis of the bZIP transcription factor family in sweet potato wild relative Ipomoea trifida. BMC Genet. 2019, 20, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, D.; Saha, D.; Acharya, D.; Mukherjee, A.; Chakraborty, S.; Ghosh, T.C. The role of introns in the conservation of the metabolic genes of Arabidopsis thaliana. Genomics 2018, 110, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Morello, L.; Gianì, S.; Troina, F.; Breviario, D. Testing the IMEter on rice introns and other aspects of intron-mediated enhancement of gene expression. J. Exp. Bot. 2011, 62, 533–544. [Google Scholar] [CrossRef]
- Bustin, S.A. Quantification of mRNA using real-time reverse transcription PCR (RT-PCR): Trends and problems. J. Mol. Endocrinol. 2002, 29, 23–39. [Google Scholar] [CrossRef]
- Thellin, O.; Zorzi, W.; Lakaye, B.; De Borman, B.; Coumans, B.; Hennen, G.; Grisar, T.; Igout, A.; Heinen, E. Housekeeping genes as internal standards: Use and limits. J. Biotechnol. 1999, 75, 291–295. [Google Scholar] [CrossRef] [Green Version]
- McDowell, J.M.; An, Y.Q.; Huang, S.; McKinney, E.C.; Meagher, R.B. The arabidopsis ACT7 actin gene is expressed in rapidly developing tissues and responds to several external stimuli. Plant Physiol. 1996, 111, 699–711. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Liu, G. Research advances in plant actin isoforms. Chin. Bull. Bot. 2006, 23, 242–248. [Google Scholar]
- Ling, Y.; Zhao, W. Specific expression of pea isoactin genes. Chin. Bull. Bot. 2001, 18, 76–80. [Google Scholar]
- Parveen, S.; Rahman, A. Actin isovariant ACT7 modulates root thermomorphogenesis by altering intracellular auxin homeostasis. Int. J. Mol. Sci. 2021, 22, 7749. [Google Scholar] [CrossRef] [PubMed]
- Numata, T.; Sugita, K.; Ahamed, R.A.; Rahman, A. Actin isovariant ACT7 controls root meristem development in Arabidopsis through modulating auxin and ethylene responses. J. Exp. Bot. 2022, 73, 6255–6271. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Jeevaraj, T.; Yunus, M.H.; Chakraborty, S.; Chakraborty, N. The plant cytoskeleton takes center stage in abiotic stress responses and resilience. Plant Cell Environ. 2023, 46, 5–22. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xu, H.; Lin, S.; Deng, W.; Zhou, J.; Zhang, Y.; Shi, Y.; Peng, D.; Xue, Y. GPS 5.0: An update on the prediction of kinase-specific phosphorylation sites in proteins. Genom. Proteom. Bioinform. 2020, 18, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Kohl, M.; Wiese, S.; Warscheid, B. Cytoscape: Software for visualization and analysis of biological networks. Methods Mol. Biol. 2011, 696, 291–303. [Google Scholar]
- Dai, Z.; Yan, P.; He, S.; Jia, L.; Wang, Y.; Liu, Q.; Zhai, H.; Zhao, N.; Gao, S.; Zhang, H. Genome-Wide Identification and Expression Analysis of SWEET Family Genes in Sweet Potato and Its Two Diploid Relatives. Int. J. Mol. Sci. 2022, 23, 15848. [Google Scholar] [CrossRef]
- Li, X.; Zhao, L.; Zhang, H.; Liu, Q.; Zhai, H.; Zhao, N.; Gao, S.; He, S. Genome-Wide Identification and Characterization of CDPK Family Reveal Their Involvements in Growth and Development and Abiotic Stress in Sweet Potato and Its Two Diploid Relatives. Int. J. Mol. Sci. 2022, 23, 3088. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Gene ID | Gene Name | PI | Molecular Weight (kDa) | CDS Length (bp) | Phosphorylation Site | Protein Size (aa) | Aliphatic Index | Instability Index | GRAVY | Subcellular Locations | Arabidopsis Homologs | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ser | Thr | Tyr | ||||||||||||
| 1 | g40378 | IbACTIN1 | 5.17 | 36.83 | 1008 | 14 | 12 | 9 | 330 | 73.27 | 36.01 | −0.268 | Cytoplasm | ACTIN1 |
| 2 | g5780 | IbACTIN2.1 | 5.05 | 48.02 | 1329 | 24 | 12 | 7 | 435 | 84.97 | 39.93 | −0.289 | Nucleus | ACTIN2 |
| 3 | g52926 | IbACTIN2.2 | 6.98 | 58.59 | 1599 | 29 | 10 | 8 | 524 | 81.49 | 51.07 | −0.355 | Nucleus | ACTIN2 |
| 4 | g4902 | IbACTIN5.1 | 5.31 | 41.73 | 1152 | 17 | 6 | 8 | 377 | 84.08 | 35.25 | −0.202 | Cytoskeleton | ACTIN5 |
| 5 | g40273 | IbACTIN5.2 | 5.31 | 41.70 | 1152 | 16 | 7 | 8 | 377 | 84.62 | 34.16 | −0.186 | Cytoskeleton | ACTIN5 |
| 6 | g9979 | IbACTIN5.3 | 5.42 | 42.32 | 1167 | 17 | 10 | 7 | 382 | 84.27 | 33.16 | −0.157 | Cytoskeleton | ACTIN5 |
| 7 | g18180 | IbACTIN5.4 | 7.87 | 12.45 | 345 | 2 | 2 | 1 | 113 | 106.02 | 36.03 | 0.15 | Cytoplasm | ACTIN5 |
| 8 | g24983 | IbACTIN5.5 | 5.31 | 41.73 | 1152 | 15 | 8 | 8 | 377 | 84.08 | 34.71 | −0.206 | Cytoskeleton | ACTIN5 |
| 9 | g43021 | IbACTIN5.6 | 5.47 | 40.56 | 1110 | 17 | 8 | 8 | 363 | 86.25 | 34.98 | −0.211 | Cytoplasm | ACTIN5 |
| 10 | g3675 | IbACTIN5.7 | 5.07 | 38.73 | 1065 | 16 | 5 | 7 | 349 | 82.12 | 34.98 | −0.143 | Cytoskeleton | ACTIN5 |
| 11 | g24981 | IbACTIN5.8 | 5.31 | 41.76 | 1152 | 17 | 8 | 8 | 377 | 83.55 | 35.22 | −0.22 | Cytoskeleton | ACTIN5 |
| 12 | g52274 | IbACTIN6 | 9.33 | 39.10 | 1053 | 26 | 8 | 6 | 345 | 79.88 | 55.76 | −0.266 | Chloroplast | ACTIN6 |
| 13 | g51848 | IbACTIN7 | 9.78 | 24.93 | 693 | 29 | 5 | 3 | 227 | 78.19 | 69.01 | −0.406 | Chloroplast | ACTIN7 |
| 14 | g25621 | IbACTIN9 | 5.38 | 41.68 | 1149 | 14 | 9 | 8 | 376 | 85.88 | 36.46 | −0.178 | Cytoskeleton | ACTIN9 |
| 15 | g2351 | IbACTIN10.1 | 5.68 | 73.78 | 1965 | 32 | 10 | 9 | 644 | 68.32 | 53.54 | −0.738 | Cytoplasm | ACTIN10 |
| 16 | g2496 | IbACTIN10.2 | 5.78 | 82.66 | 2196 | 35 | 11 | 9 | 720 | 68.83 | 54.01 | −0.749 | Cytoplasm | ACTIN10 |
| 17 | g64096 | IbACTIN12.1 | 6.11 | 57.53 | 1551 | 20 | 14 | 7 | 508 | 81.48 | 46.25 | −0.305 | Cytoplasm | ACTIN12 |
| 18 | g48567 | IbACTIN12.2 | 8.54 | 33.65 | 915 | 11 | 9 | 5 | 300 | 83.47 | 43.54 | −0.202 | Chloroplast | ACTIN12 |
| 19 | g18621 | IbACTIN12.3 | 9.05 | 28.13 | 771 | 18 | 10 | 4 | 252 | 80.91 | 53 | −0.208 | Nucleus | ACTIN12 |
| 20 | g26691 | IbACTIN13 | 6.43 | 57.52 | 1584 | 31 | 21 | 3 | 519 | 91.81 | 42.68 | −0.125 | Cytoplasm | ACTIN13 |
| 21 | g58376 | IbACTIN15 | 5.54 | 38.77 | 1065 | 16 | 8 | 7 | 349 | 86.1 | 36.11 | −0.193 | Cytoskeleton | ACTIN15 |
| 22 | g42892 | IbACTIN16.1 | 4.71 | 27.97 | 783 | 7 | 6 | 2 | 256 | 96.41 | 45.39 | −0.026 | Cytoplasm | ACTIN16 |
| 23 | g63730 | IbACTIN16.2 | 4.79 | 40.02 | 1116 | 11 | 10 | 5 | 365 | 91.37 | 45.53 | −0.081 | Cytoplasm | ACTIN16 |
| 24 | g46627 | IbACTIN17.1 | 5.1 | 41.61 | 1152 | 19 | 6 | 8 | 377 | 86.13 | 31.71 | −0.154 | Cytoskeleton | ACTIN17 |
| 25 | g60046 | IbACTIN17.2 | 5.31 | 41.74 | 1152 | 17 | 8 | 8 | 377 | 85.86 | 33.06 | −0.162 | Cytoskeleton | ACTIN17 |
| 26 | g27691 | IbACTIN17.3 | 5.42 | 36.18 | 996 | 15 | 4 | 5 | 326 | 89.45 | 42.1 | −0.085 | Cytoplasm | ACTIN17 |
| 27 | g723 | IbACTIN17.4 | 5.57 | 46.15 | 1275 | 19 | 12 | 7 | 418 | 92.8 | 40.29 | −0.057 | Cytoplasm | ACTIN17 |
| 28 | g13139 | IbACTIN18 | 5.86 | 62.01 | 1674 | 28 | 18 | 10 | 548 | 85.05 | 53.27 | −0.389 | Chloroplast | ACTIN18 |
| 29 | g61314 | IbACTIN19 | 7.89 | 55.85 | 1533 | 36 | 17 | 10 | 502 | 83.92 | 43.11 | 0.001 | Chloroplast | ACTIN19 |
| 30 | g20278 | IbACTIN20 | 5.37 | 41.79 | 1152 | 15 | 11 | 8 | 377 | 84.62 | 35.09 | −0.19 | Cytoskeleton | ACTIN20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, S.; Zhang, H.; He, S. Genome-Wide Identification and Expression Analysis of ACTIN Family Genes in the Sweet Potato and Its Two Diploid Relatives. Int. J. Mol. Sci. 2023, 24, 10930. https://doi.org/10.3390/ijms241310930
Xia S, Zhang H, He S. Genome-Wide Identification and Expression Analysis of ACTIN Family Genes in the Sweet Potato and Its Two Diploid Relatives. International Journal of Molecular Sciences. 2023; 24(13):10930. https://doi.org/10.3390/ijms241310930
Chicago/Turabian StyleXia, Shuanghong, Huan Zhang, and Shaozhen He. 2023. "Genome-Wide Identification and Expression Analysis of ACTIN Family Genes in the Sweet Potato and Its Two Diploid Relatives" International Journal of Molecular Sciences 24, no. 13: 10930. https://doi.org/10.3390/ijms241310930