
CRP in Outpatients with Inflammatory Bowel Disease Is Linked to the Blood Microbiota
 and
and
Abstract
:
1. Introduction
2. Results
2.1. Patient Characteristics
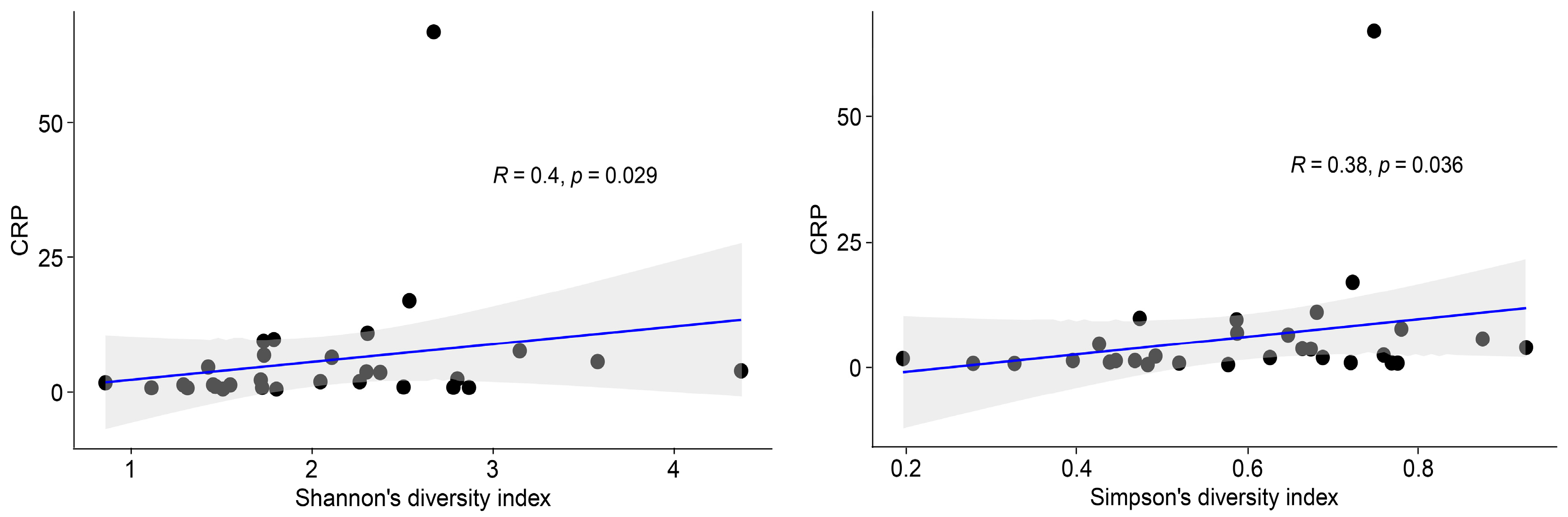
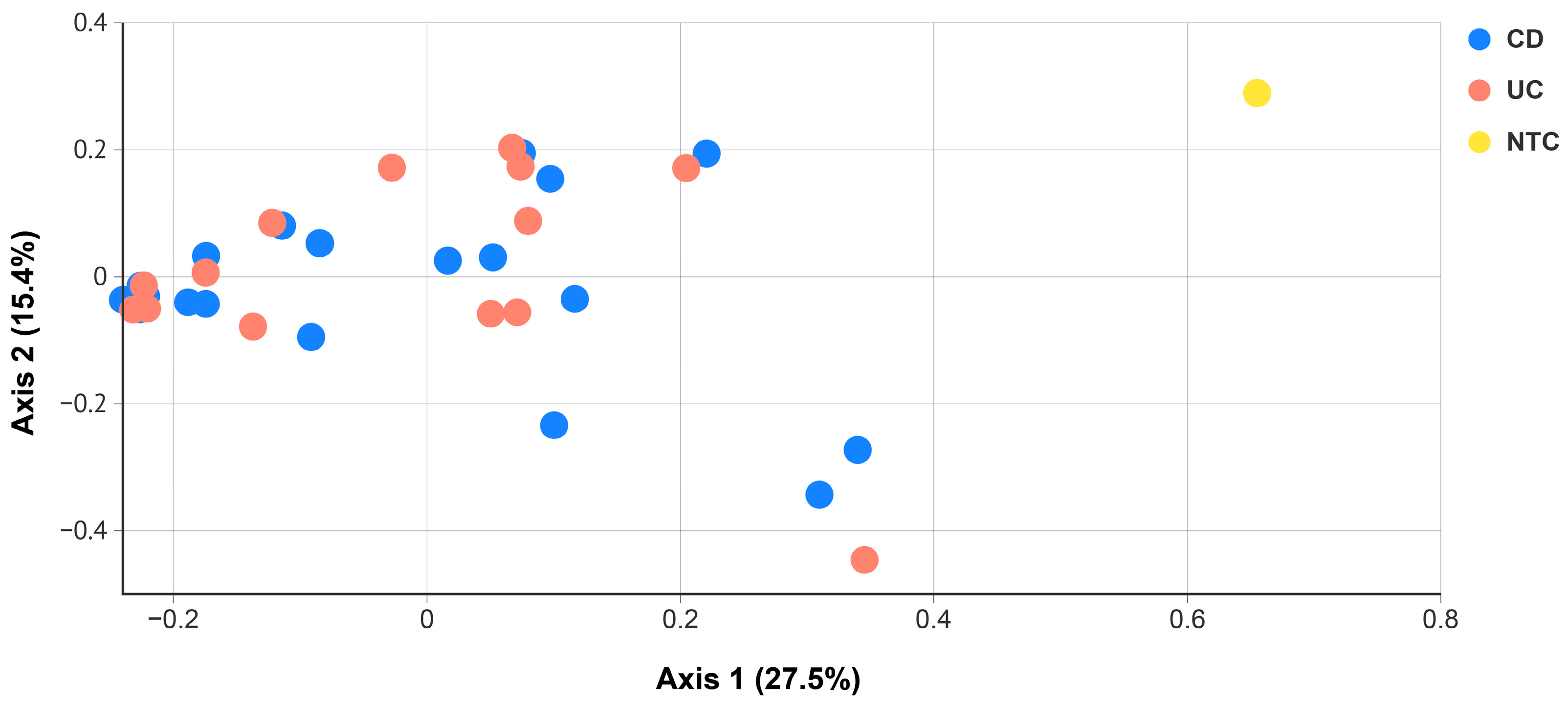
2.2. Blood Microbiota in IBD Patients
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. 16S-Targeted Metagenomics
4.3. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nikkari, S.; McLaughlin, I.J.; Bi, W.L.; Dodge, D.E.; Relman, D.A. Does blood of healthy subjects contain bacterial ribosomal DNA? J. Clin. Microbiol. 2001, 39, 1956–1959. [Google Scholar] [CrossRef] [Green Version]
- Moriyama, K.; Ando, C.; Tashiro, K.; Kuhara, S.; Okamura, S.; Nakano, S.; Takagi, Y.; Miki, T.; Nakashima, Y.; Hirakawa, H. Polymerase chain reaction detection of bacterial 16S rRNA gene in human blood. Microbiol. Immunol. 2008, 52, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Damgaard, C.; Magnussen, K.; Enevold, C.; Nilsson, M.; Tolker-Nielsen, T.; Holmstrup, P.; Nielsen, C.H. Viable Bacteria Associated with Red Blood Cells and Plasma in Freshly Drawn Blood Donations. PLoS ONE 2015, 10, e0120826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo, D.J.; Rifkin, R.F.; Cowan, D.A.; Potgieter, M. The Healthy Human Blood Microbiome: Fact or Fiction? Front. Cell. Infect. Microbiol. 2019, 9, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, M.T.; Amos, G.C.A.; Murphy, A.R.J.; Murch, S.; Wellington, E.M.H.; Arasaradnam, R.P. Microbial imbalance in inflammatory bowel disease patients at different taxonomic levels. Gut Pathog. 2020, 12, 1. [Google Scholar] [CrossRef]
- Frank, D.N.; Amand, A.L.S.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef] [Green Version]
- Ott, S.J.; Musfeldt, M.; Wenderoth, D.F.; Hampe, J.; Brant, O.; Folsch, U.R.; Timmins, K.N.; Schreiber, S. Reduction in diversity of the colonic mucosa associated bacterial microflora in patients with active inflammatory bowel disease. Gut 2004, 53, 685–693. [Google Scholar] [CrossRef] [Green Version]
- Pisani, A.; Rausch, P.; Bang, C.; Ellul, S.; Tabone, T.; Cordina, C.M.; Zahra, G.; Franke, A.; Ellul, P. Dysbiosis in the Gut Microbiota in Patients with Inflammatory Bowel Disease during Remission. Microbiol. Spectr. 2022, 10, e0061622. [Google Scholar] [CrossRef]
- Serrano-Gomez, G.; Mayorga, L.; Oyarzun, I.; Roca, J.; Borruel, N.; Casellas, F.; Varela, E.; Pozuelo, M.; Machiels, K.; Guarner, F.; et al. Dysbiosis and relapse-related microbiome in inflammatory bowel disease: A shotgun metagenomic approach. Comput. Struct. Biotechnol. J. 2021, 19, 6481–6489. [Google Scholar] [CrossRef]
- Khan, I.; Bai, Y.R.; Zha, L.J.; Ullah, N.; Ullah, H.; Shah, S.R.H.; Sun, H.; Zhang, C.J. Mechanism of the Gut Microbiota Colonization Resistance and Enteric Pathogen Infection. Front. Cell. Infect. Microbiol. 2021, 11, 716299. [Google Scholar] [CrossRef] [PubMed]
- Vrakas, S.; Mountzouris, K.C.; Michalopoulos, G.; Karamanolis, G.; Papatheodoridis, G.; Tzathas, C.; Gazouli, M. Intestinal Bacteria Composition and Translocation of Bacteria in Inflammatory Bowel Disease. PLoS ONE 2017, 12, e0170034. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, A.; Frances, R.; Amoros, A.; Zapater, P.; Garmendia, M.; NDongo, M.; Cano, R.; Jover, R.; Such, J.; Perez-Mateo, M. Cytokine Association with Bacterial DNA in Serum of Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2009, 15, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Ishida, N.; Higuchi, T.; Miyazu, T.; Tamura, S.; Tani, S.; Yamade, M.; Iwaizumi, M.; Hamaya, Y.; Osawa, S.; Furuta, T.; et al. C-reactive protein is superior to fecal biomarkers for evaluating colon-wide active inflammation in ulcerative colitis. Sci. Rep.-UK 2021, 11, 12431. [Google Scholar] [CrossRef]
- Lewis, J.D. The Utility of Biomarkers in the Diagnosis and Therapy of Inflammatory Bowel Disease. Gastroenterology 2011, 140, 1817–1826.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiga, H.; Abe, I.; Onodera, M.; Moroi, R.; Kuroha, M.; Kanazawa, Y.; Kakuta, Y.; Endo, K.; Kinouchi, Y.; Masamune, A. Serum C-reactive protein and albumin are useful biomarkers for tight control management of Crohn’s disease in Japan. Sci. Rep.-UK 2020, 10, 511. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Patel, D.; Shah, Y.; Trivedi, C.; Yang, Y.X. Albumin as a prognostic marker for ulcerative colitis. World J. Gastroenterol. 2017, 23, 8008–8016. [Google Scholar] [CrossRef] [PubMed]
- Vernia, F.; Di Ruscio, M.; Stefanelli, G.; Viscido, A.; Frieri, G.; Latella, G. Is fecal calprotectin an accurate marker in the management of Crohn’s disease? J. Gastroen Hepatol. 2020, 35, 390–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Cerezo, J.; Suarez, I.; Rios, J.J.; Pena, P.; de Miguel, M.J.G.; de Jose, M.; Monteagudo, O.; Linares, P.; Barbado-Cano, A.; Vazquez, J.J. Achromobacter xylosoxidans bacteremia: A 10-year analysis of 54 cases. Eur. J. Clin. Microbiol. 2003, 22, 360–363. [Google Scholar] [CrossRef]
- Yoon, S.H.; Kim, H.; Lim, S.M.; Kang, J.M. Nosocomial outbreak of Achromobacter spp. bacteremia due to germicide contamination: A systematic review. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 6374–6381. [Google Scholar]
- Marion-Sanchez, K.; Pailla, K.; Olive, C.; Le Coutour, X.; Derancourt, C. Achromobacter spp. healthcare associated infections in the French West Indies: A longitudinal study from 2006 to 2016. BMC Infect. Dis. 2019, 19, 795. [Google Scholar] [CrossRef]
- Isler, B.; Kidd, T.J.; Stewart, A.G.; Harris, P.; Paterson, D.L. Achromobacter Infections and Treatment Options. Antimicrob. Agents Chemother. 2020, 64, e01025-20. [Google Scholar] [CrossRef] [PubMed]
- Weitkamp, J.H.; Tang, Y.W.; Haas, D.W.; Midha, N.K.; Crowe, J.E., Jr. Recurrent Achromobacter xylosoxidans bacteremia associated with persistent lymph node infection in a patient with hyper-immunoglobulin M syndrome. Clin. Infect. Dis. 2000, 31, 1183–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Z.; Wu, J.J.; Gong, J.L.; Ke, J.; Ding, T.; Zhao, W.J.; Cheng, W.M.; Luo, Z.H.; He, Q.L.; Zeng, W.Y.; et al. Microbiota in mesenteric adipose tissue from Crohn’s disease promote colitis in mice. Microbiome 2021, 9, 228. [Google Scholar] [CrossRef]
- Houry, A.; Gohar, M.; Deschamps, J.; Tischenko, E.; Aymerich, S.; Gruss, A.; Briandet, R. Bacterial swimmers that infiltrate and take over the biofilm matrix. Proc. Natl. Acad. Sci. USA 2012, 109, 13088–13093. [Google Scholar] [CrossRef] [Green Version]
- Walujkar, S.A.; Kumbhare, S.V.; Marathe, N.P.; Patangia, D.V.; Lawate, P.S.; Bharadwaj, R.S.; Shouche, Y.S. Molecular profiling of mucosal tissue associated microbiota in patients manifesting acute exacerbations and remission stage of ulcerative colitis. World J. Microb. Biot. 2018, 34, 76. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Price, L.S.; Weinstein, R.A. Acinetobacter infection. N. Engl. J. Med. 2008, 358, 1271–1281. [Google Scholar] [CrossRef]
- Hafiz, T.A.; Alghamdi, S.S.; Mubaraki, M.A.; Alghamdi, S.S.M.; Alothaybi, A.; Aldawood, E.; Alotaibi, F. A two-year retrospective study of multidrug-resistant Acinetobacter baumannii respiratory infections in critically Ill patients: Clinical and microbiological findings. J. Infect. Public. Health 2023, 16, 313–319. [Google Scholar] [CrossRef]
- van Spanning, R.J.; Stouthamer, A.H.; Baker, S.C.; van Verseveld, H.W. Paracoccus. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Gossner, A.S.; Devereux, R.; Ohnemuller, N.; Acker, G.; Stackebrandt, E.; Drake, H.L. Thermicanus aegyptius gen. nov., sp nov., isolated from oxic soil, a fermentative microaerophile that grows commensally with the thermophilic acetogen Moorella thermoacetica. Appl. Environ. Microb. 1999, 65, 5124–5133. [Google Scholar] [CrossRef] [Green Version]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoetendal, E.G.; Rajilic-Stojanovic, M.; de Vos, W.M. High-throughput diversity and functionality analysis of the gastrointestinal tract microbiota. Gut 2008, 57, 1605–1615. [Google Scholar] [CrossRef]
- Hannigan, G.D.; Grice, E.A. Microbial Ecology of the Skin in the Era of Metagenomics and Molecular Microbiology. Csh Perspect. Med. 2013, 3, a015362. [Google Scholar] [CrossRef] [Green Version]
- Swidsinski, A.; Weber, J.; Loening-Baucke, V.; Hale, L.P.; Lochs, H. Spatial organization and composition of the mucosal flora in patients with inflammatory bowel disease. J. Clin. Microbiol. 2005, 43, 3380–3389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Molin, G.; Ahrne, S.; Adawi, D.; Jeppsson, B. High proportions of proinflammatory bacteria on the colonic mucosa in a young patient with ulcerative colitis as revealed by cloning and sequencing of 16S rRNA genes. Dig. Dis. Sci. 2007, 52, 620–627. [Google Scholar] [CrossRef] [PubMed]
- Imhann, F.; Vila, A.V.; Bonder, M.J.; Fu, J.Y.; Gevers, D.; Visschedijk, M.C.; Spekhorst, L.M.; Alberts, R.; Franke, L.; van Dullemen, H.M.; et al. Interplay of host genetics and gut microbiota underlying the onset and clinical presentation of inflammatory bowel disease. Gut 2018, 67, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Altomare, A.; Putignani, L.; Del Chierico, F.; Cocca, S.; Angeletti, S.; Ciccozzi, M.; Tripiciano, C.; Piccola, B.D.; Cicala, M.; Guarino, M.P.L. Gut mucosal-associated microbiota better discloses inflammatory bowel disease differential patterns than faecal microbiota. Dig. Liver Dis. 2019, 51, 648–656. [Google Scholar] [CrossRef]
- Iwasawa, K.; Suda, W.; Tsunoda, T.; Oikawa-Kawamoto, M.; Umetsu, S.; Takayasu, L.; Inui, A.; Fujisawa, T.; Morita, H.; Sogo, T.; et al. Dysbiosis of the salivary microbiota in pediatric-onset primary sclerosing cholangitis and its potential as a biomarker. Sci. Rep. 2018, 8, 5480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajer, L.; Kverka, M.; Kostovcik, M.; Macinga, P.; Dvorak, J.; Stehlikova, Z.; Brezina, J.; Wohl, P.; Spicak, J.; Drastich, P. Distinct gut microbiota profiles in patients with primary sclerosing cholangitis and ulcerative colitis. World J. Gastroenterol. 2017, 23, 4548–4558. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2 ’ s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Whole Group | Crohn’s Disease | Ulcerative Colitis | |||
|---|---|---|---|---|---|
| Patients (n) | 33 | 19 | 14 | ||
| Age, years (range) | 43 (19–70) | 42 (23–70) | 46 (19–69) | ||
| Duration, years (range) | 12 (0–46) | 14 (0–46) | 9 (0–29) | ||
| Gender | 11 F/22 M | 7 F/12 M | 4 F/10 M | ||
| Localisation | L1 | 6 | E1 | 0 | |
| L2 | 3 | E2 | 5 | ||
| L3 | 10 | E3 | 9 | ||
| Behaviour | B1 | 9 | |||
| B2 | 6 | ||||
| B3 | 2 | ||||
| B2/B3 | 1 | ||||
| B3/B4 | 1 | ||||
| Missing | 0 | ||||
| Smoking (n) | |||||
| Never | 16 | 9 | 7 | ||
| Past | 13 | 6 | 7 | ||
| Current | 4 | 4 | 0 | ||
| Laboratory data | |||||
| CRP, mg/L (range) | 6.0 (0.6–67) | 3.1 (0.8–7.7) | 9.7 (0.6–67) | ||
| Median (IQR) | 2.2 (1.0–6.3) | 2.3 (1.1–4.7) | 2.0 (0.9–9.8) | ||
| Hb, g/L (range) | 134 (102–163) | 136 (102–163) | 132 (108–153) | ||
| Albumin, g/L (range) | 41 (21–49) | 40 (21–49) | 41 (33–47) | ||
| ASVs * | Phylum | Family | Genus | Species | |
|---|---|---|---|---|---|
| CD | 108,217 (49,524–245,291) | 5 | 28 | 36 | 43 |
| UC | 120,973 (75,627–220,242) | 5 | 23 | 32 | 42 |
| NTC | 78,209 | 3 | 6 | 6 | 7 |
| Shannon’s Diversity Index Median [IQR] | Simpson’s Diversity Index Median [IQR] | p-Value | |
|---|---|---|---|
| CD patients (n = 19) | 2.05 [1.53–2.51] | 0.65 [0.47–0.72] | 0.87 |
| UC patients (n = 14) | 2.04 [1.46–2.57] | 0.61 [0.45–0.73] | 0.93 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Molin, G.; Davidson, S.; Roth, B.; Sjöberg, K.; Håkansson, Å. CRP in Outpatients with Inflammatory Bowel Disease Is Linked to the Blood Microbiota. Int. J. Mol. Sci. 2023, 24, 10899. https://doi.org/10.3390/ijms241310899
Xu J, Molin G, Davidson S, Roth B, Sjöberg K, Håkansson Å. CRP in Outpatients with Inflammatory Bowel Disease Is Linked to the Blood Microbiota. International Journal of Molecular Sciences. 2023; 24(13):10899. https://doi.org/10.3390/ijms241310899
Chicago/Turabian StyleXu, Jie, Göran Molin, Sanna Davidson, Bodil Roth, Klas Sjöberg, and Åsa Håkansson. 2023. "CRP in Outpatients with Inflammatory Bowel Disease Is Linked to the Blood Microbiota" International Journal of Molecular Sciences 24, no. 13: 10899. https://doi.org/10.3390/ijms241310899