
Mesenchymal Stem Cell Conditioned Medium Modulates Inflammation in Tenocytes: Complete Conditioned Medium Has Superior Therapeutic Efficacy than Its Extracellular Vesicle Fraction
 , , and
, , and
Abstract
:1. Introduction
2. Results
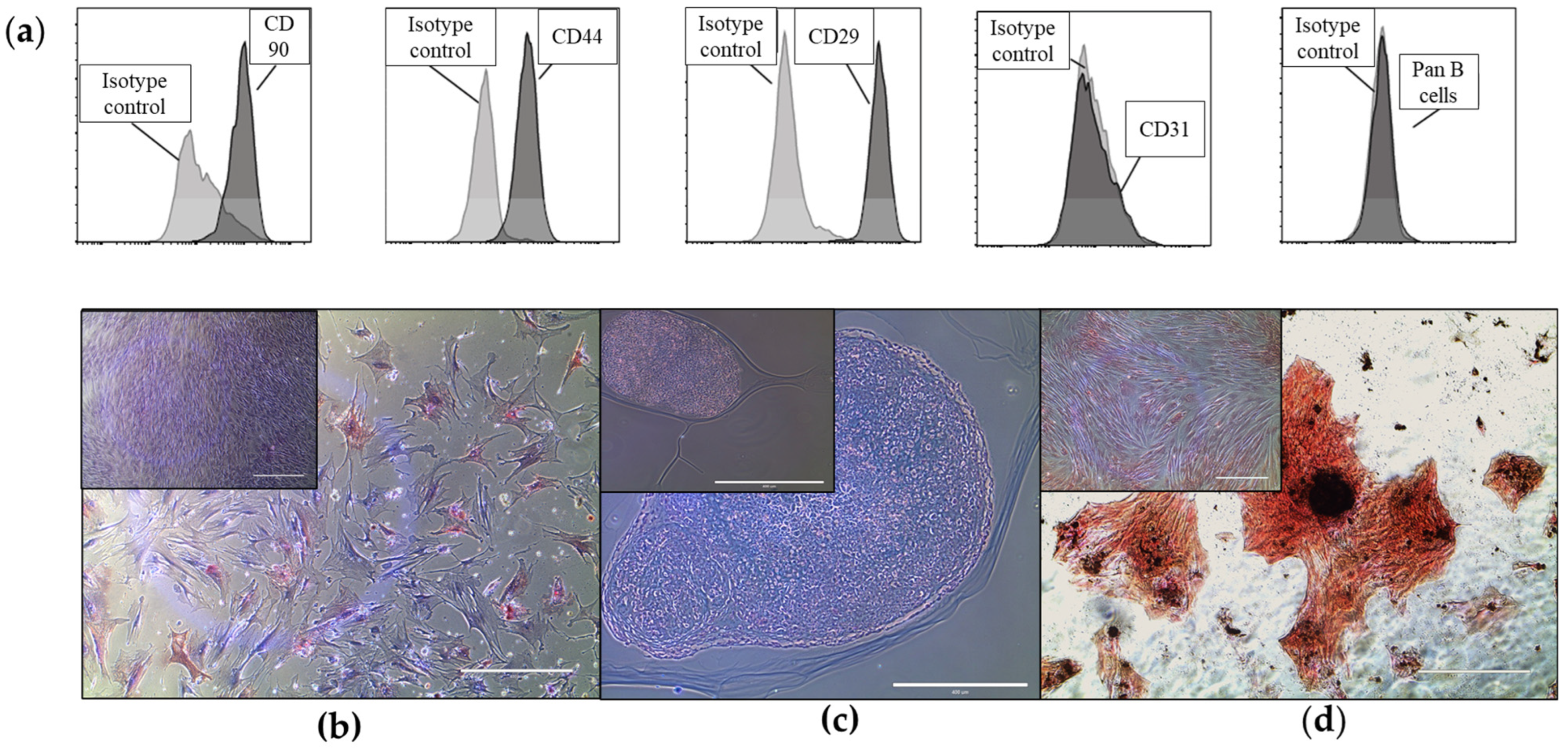
2.1. Equine Bone Marrow-Derived MSCs and EVs Show Characteristic Markers
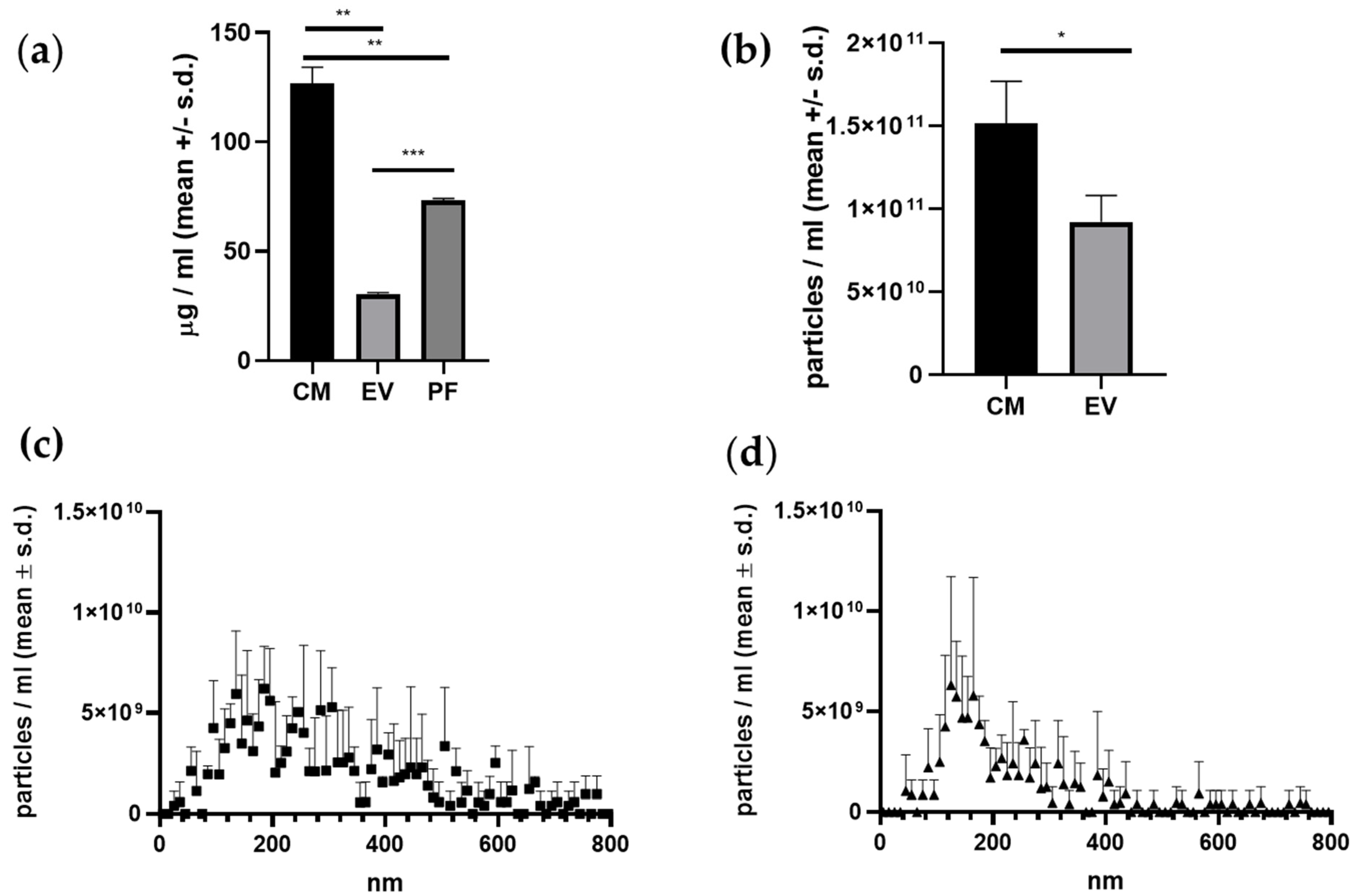
2.2. CM Fraction Has the Highest Protein- and Particle Concentration
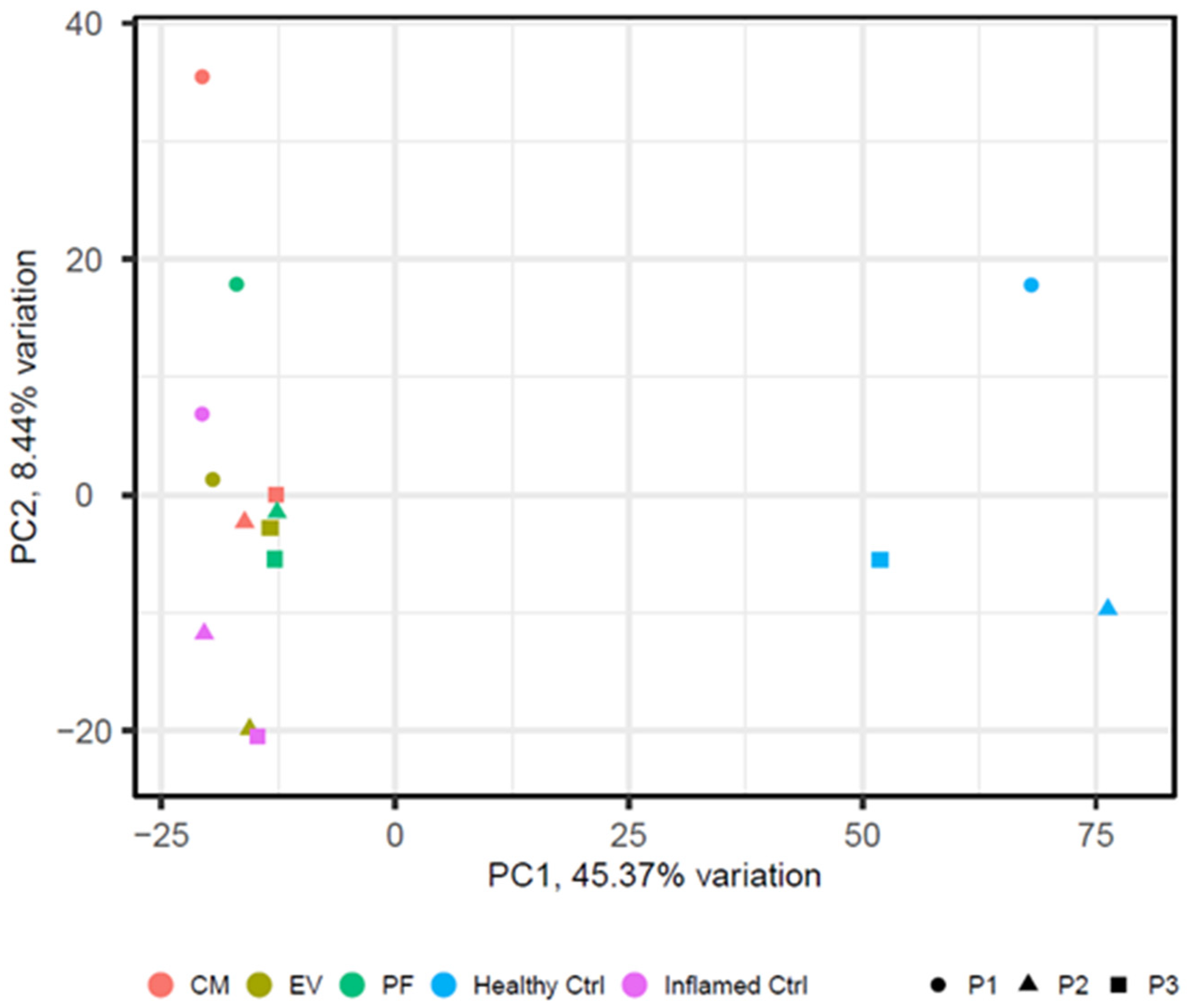
2.3. Inflammation Induces a Significant Change in Tenocytes’ Gene Expression
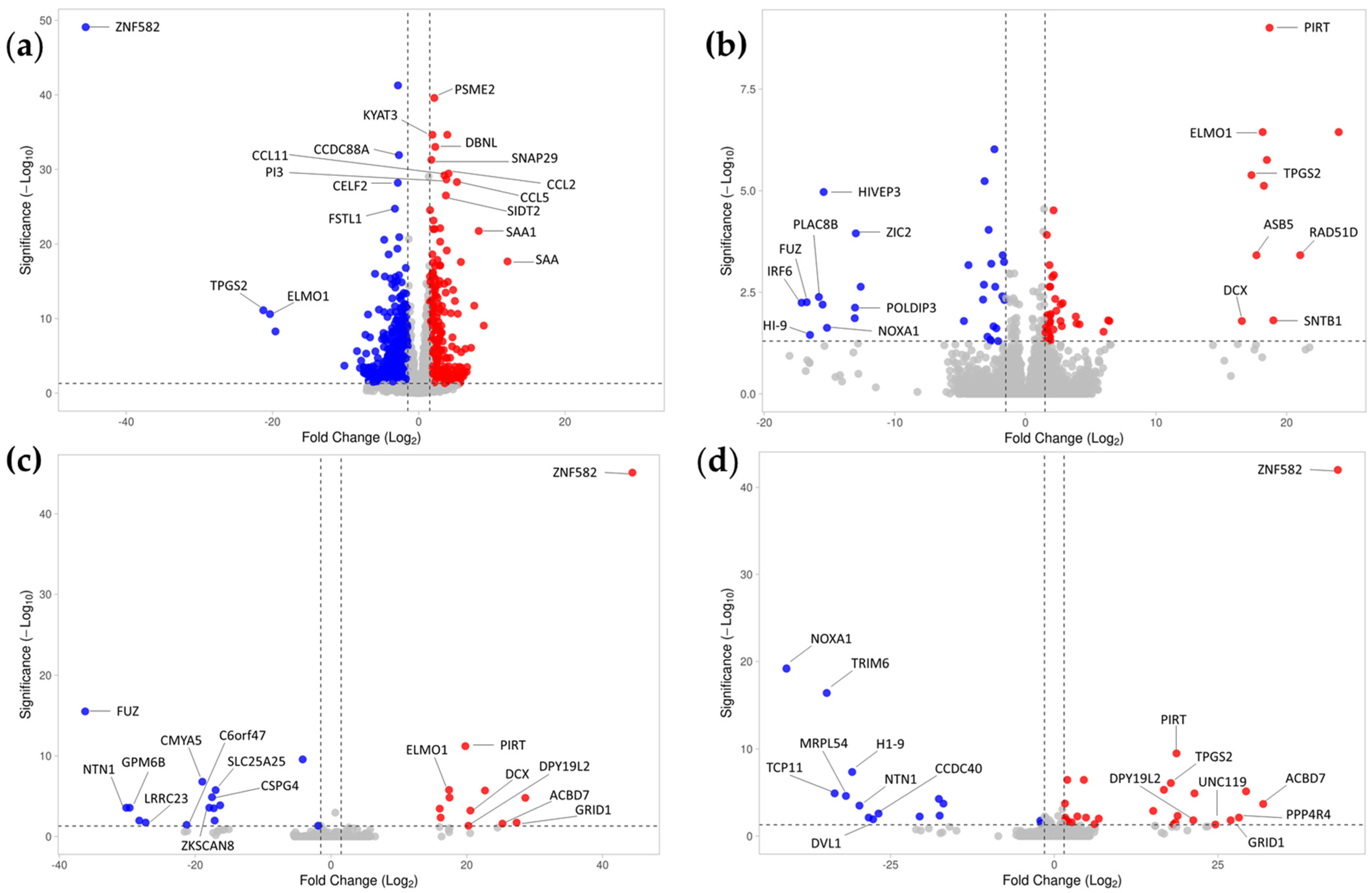
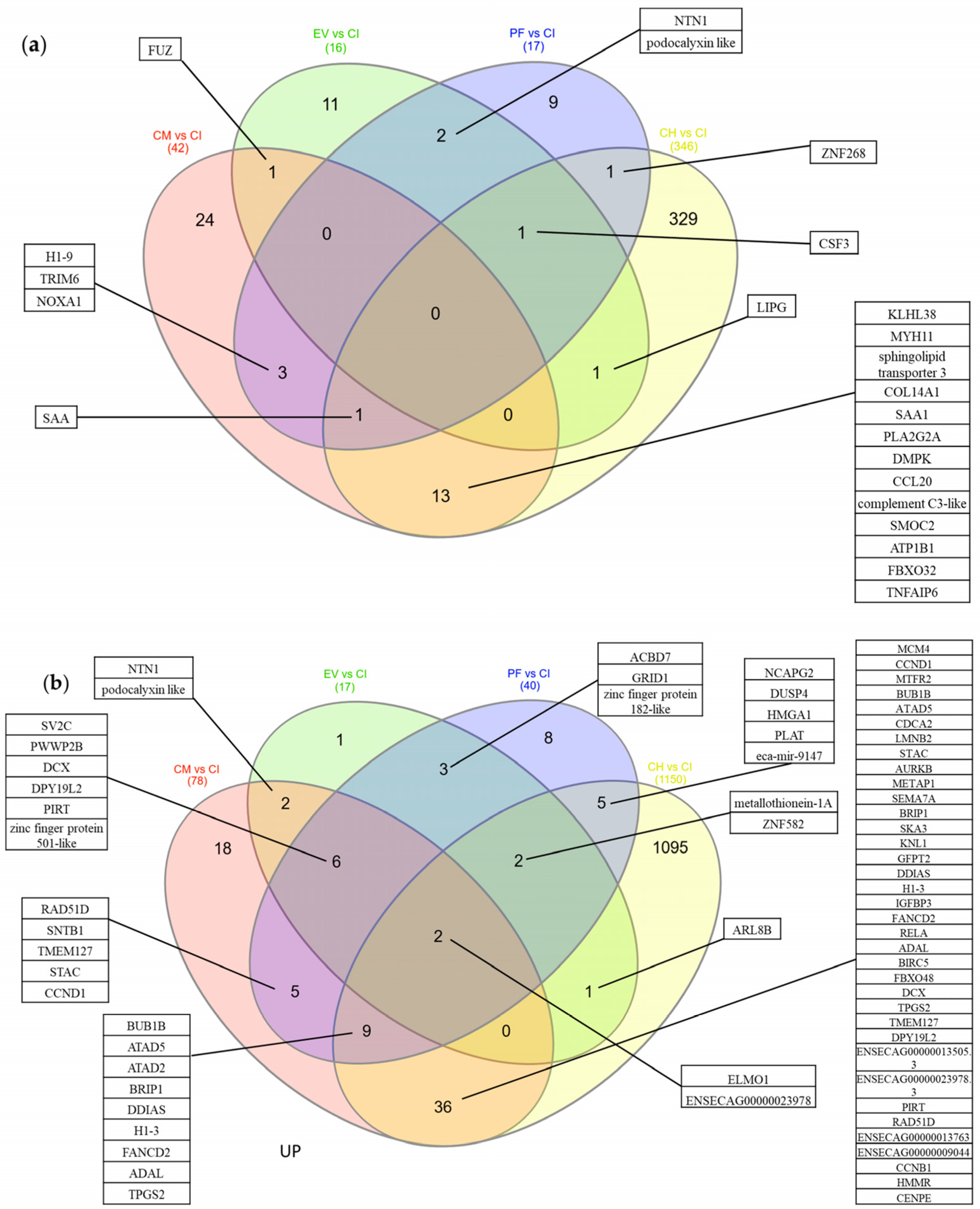
2.4. Conditioned Medium Achieves a Higher Effect on Differential Gene Expression of Inflamed Tenocytes Than EVs or PF Alone
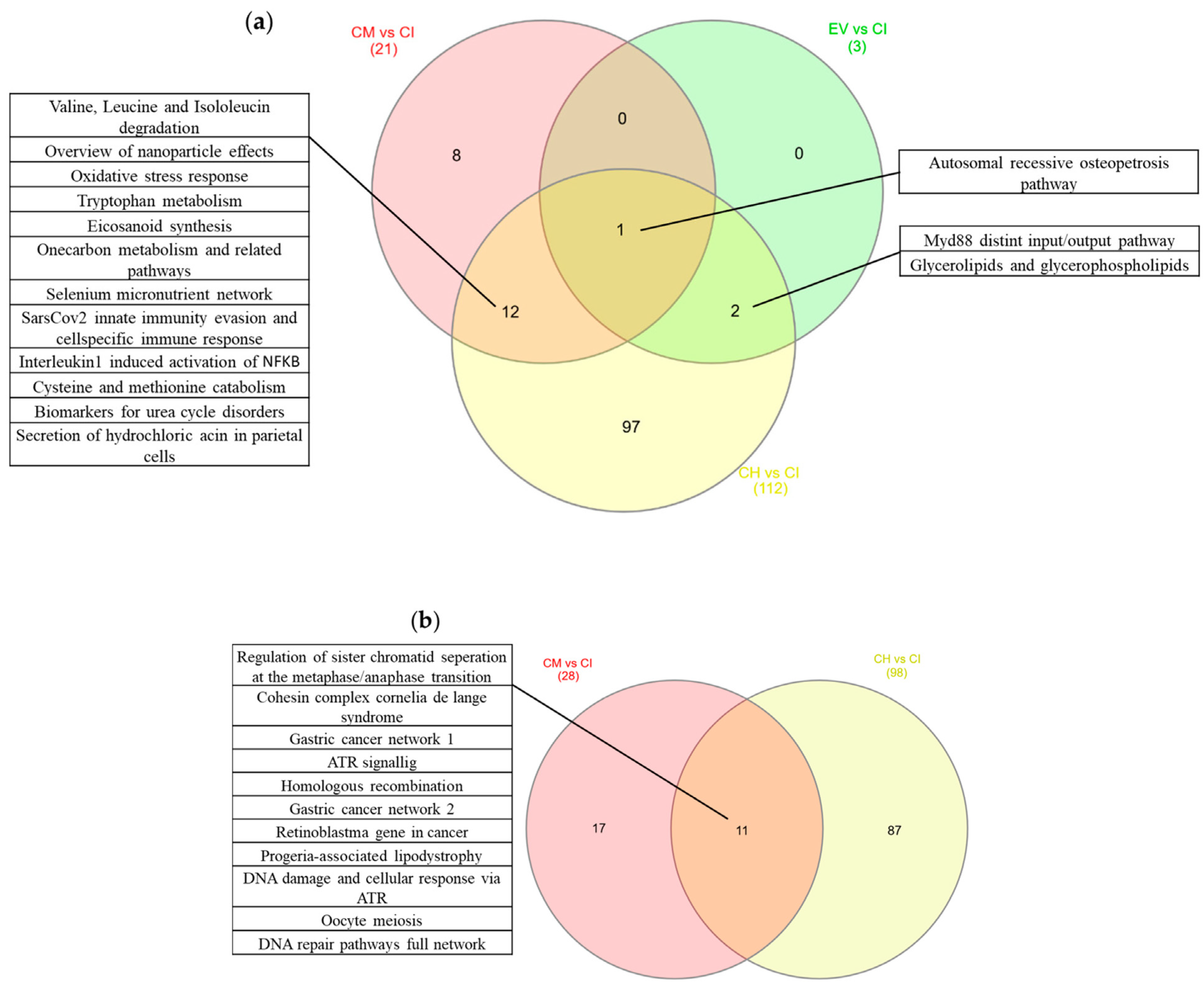
2.5. Pathways
3. Discussion
4. Materials and Methods
4.1. Bone Marrow Collection and MSC Isolation
4.2. Isolation of the Conditioned Medium (Fractions), Characterization, and Quantification
4.3. Tenocyte Isolation
4.4. Inflammation and Treatment Strategy
4.5. Flow Cytometry
4.6. Trilineage Differentiation
4.6.1. Oil Red Staining
4.6.2. Alcian Blue Staining
4.6.3. Von Kossa Stain
4.7. mRNA-Sequencing of the “Patient Cells” (Tenocytes)
4.8. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Docheva, D.; Müller, S.A.; Majewski, M.; Evans, C.H. Biologics for tendon repair. Adv. Drug Deliv. Rev. 2015, 84, 222–239. [Google Scholar] [CrossRef] [Green Version]
- Walden, G.; Liao, X.; Donell, S.; Raxworthy, M.J.; Riley, G.P.; Saeed, A. A Clinical, Biological, and Biomaterials Perspective into Tendon Injuries and Regeneration. Tissue Eng. Part B Rev. 2016, 23, 44–58. [Google Scholar] [CrossRef] [Green Version]
- Millar, N.L.; Silbernagel, K.G.; Thorborg, K.; Kirwan, P.D.; Galatz, L.M.; Abrams, G.D.; Murrell, G.A.C.; McInnes, I.B.; Rodeo, S.A. Tendinopathy. Nat. Rev. Dis. Prim. 2021, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Tanzil, G.; Al-Sadi, O.; Wiegand, E.; Ertel, W.; Busch, C.; Kohl, B.; Pufe, T. The role of pro-inflammatory and immunoregulatory cytokines in tendon healing and rupture: New insights. Scand. J. Med. Sci. Sports 2011, 21, 337–351. [Google Scholar] [CrossRef]
- Thomopoulos, S.; Parks, W.C.; Rifkin, D.B.; Derwin, K.A. Mechanisms of tendon injury and repair. J. Orthop. Res. 2015, 33, 832–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cambré, I.; Gaublomme, D.; Burssens, A.; Jacques, P.; Schryvers, N.; De Muynck, A.; Meuris, L.; Lambrecht, S.; Carter, S.; de Bleser, P.; et al. Mechanical strain determines the site-specific localization of inflammation and tissue damage in arthritis. Nat. Commun. 2018, 9, 4613. [Google Scholar] [CrossRef] [Green Version]
- Maffulli, N.; Ewen, S.W.B.; Waterston, S.W.; Reaper, J.; Barrass, V. Tenocytes from Ruptured and Tendinopathic Achilles Tendons Produce Greater Quantities of Type III Collagen than Tenocytes from Normal Achilles Tendons: An in Vitro Model of Human Tendon Healing. Am. J. Sports Med. 2000, 28, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Nichols, A.E.C.; Best, K.T.; Loiselle, A.E. The cellular basis of fibrotic tendon healing: Challenges and opportunities. Transl. Res. 2019, 209, 156–168. [Google Scholar] [CrossRef]
- Abraham, A.C.; Shah, S.A.; Golman, M.; Song, L.; Li, X.; Kurtaliaj, I.; Akbar, M.; Millar, N.L.; Abu-Amer, Y.; Galatz, L.M.; et al. Targeting the NF-κB signaling pathway in chronic tendon disease. Sci. Transl. Med. 2019, 11, eaav4319. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Chen, Y.; Huang, J.; Zhao, K.; Chen, X.; Yin, Z.; Heng, B.C.; Chen, W.; Shen, W. The roles of inflammatory mediators and immunocytes in tendinopathy. J. Orthop. Transl. 2018, 14, 23–33. [Google Scholar] [CrossRef]
- Dakin, S.G.; Newton, J.; Martinez, F.O.; Hedley, R.; Gwilym, S.; Jones, N.; Reid, H.A.B.; Wood, S.; Wells, G.; Appleton, L.; et al. Chronic inflammation is a feature of Achilles tendinopathy and rupture. Br. J. Sports Med. 2018, 52, 359–367. [Google Scholar] [CrossRef]
- Dakin, S.G. Inflammation or damage: Fibroblasts decide. Sci. Transl. Med. 2019, 11, eaax9562. [Google Scholar] [CrossRef]
- Young, R.G.; Butler, D.L.; Weber, W.; Caplan, A.I.; Gordon, S.L.; Fink, D.J. Use of mesenchymal stem cells in a collagen matrix for achilles tendon repair. J. Orthop. Res. 1998, 16, 406–413. [Google Scholar] [CrossRef]
- Smith, R.K.W.; Korda, M.; Blunn, G.W.; Goodship, A.E. Isolation and implantation of autologous equine mesenchymal stem cells from bone marrow into the superficial digital flexor tendon as a potential novel treatment. Equine Vet. J. 2003, 35, 99–102. [Google Scholar] [CrossRef]
- Smith, R.K.W. Mesenchymal stem cell therapy for equine tendinopathy. Disabil. Rehabil. 2008, 30, 1752–1758. [Google Scholar] [CrossRef]
- Smith, R.K.W.; Webbon, P.M. Harnessing the stem cell for the treatment of tendon injuries: Heralding a new dawn? Br. J. Sports Med. 2005, 39, 582–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.K.W.; Werling, N.J.; Dakin, S.G.; Alam, R.; Goodship, A.E.; Dudhia, J. Beneficial Effects of Autologous Bone Marrow-Derived Mesenchymal Stem Cells in Naturally Occurring Tendinopathy. PLoS ONE 2013, 8, e75697. [Google Scholar] [CrossRef]
- Dyson, S.J. Medical management of superficial digital flexor tendonitis: A comparative study in 219 horses (1992–2000). Equine Vet. J. 2004, 36, 415–419. [Google Scholar] [CrossRef] [Green Version]
- Godwin, E.E.; Young, N.J.; Dudhia, J.; Beamish, I.C.; Smith, R.K.W. Implantation of bone marrow-derived mesenchymal stem cells demonstrates improved outcome in horses with overstrain injury of the superficial digital flexor tendon. Equine Vet. J. 2012, 44, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Costa-Almeida, R.; Calejo, I.; Gomes, M.E. Mesenchymal Stem Cells Empowering Tendon Regenerative Therapies. Int. J. Mol. Sci. 2019, 20, 3002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horwitz, E.M.; Gordon, P.L.; Koo, W.K.K.; Marx, J.C.; Neel, M.D.; McNall, R.Y.; Muul, L.; Hofmann, T. Isolated allogeneic bone marrow-derived mesenchymal cells engraft and stimulate growth in children with osteogenesis imperfecta: Implications for cell therapy of bone. Proc. Natl. Acad. Sci. USA 2002, 99, 8932–8937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, L.; Schubert, S.; Scheibe, P.; Brehm, W.; Franzen, J.; Gross, C.; Burk, J. Tenogenic Properties of Mesenchymal Progenitor Cells Are Compromised in an Inflammatory Environment. Int. J. Mol. Sci. 2018, 19, 2549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnecchi, M.; He, H.; Liang, O.D.; Melo, L.G.; Morello, F.; Mu, H.; Noiseux, N.; Zhang, L.; Pratt, R.E.; Ingwall, J.S.; et al. Paracrine action accounts for marked protection of ischemic heart by Akt-modified mesenchymal stem cells. Nat. Med. 2005, 11, 367–368. [Google Scholar] [CrossRef]
- Caplan, A.I.; Correa, D. The MSC: An injury drugstore. Cell Stem Cell 2011, 9, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Baldari, S.; Di Rocco, G.; Piccoli, M.; Pozzobon, M.; Muraca, M.; Toietta, G. Challenges and Strategies for Improving the Regenerative Effects of Mesenchymal Stromal Cell-Based Therapies. Int. J. Mol. Sci. 2017, 18, 2087. [Google Scholar] [CrossRef] [Green Version]
- Timmers, L.; Lim, S.K.; Hoefer, I.E.; Arslan, F.; Lai, R.C.; van Oorschot, A.A.M.; Goumans, M.J.; Strijder, C.; Sze, S.K.; Choo, A.; et al. Human mesenchymal stem cell-conditioned medium improves cardiac function following myocardial infarction. Stem Cell Res. 2011, 6, 206–214. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, G.; Plaza, A.; Schenffeldt, A.; Alarcón, P.; Gajardo, G.; Uberti, B.; Morán, G.; Henríquez, C. Equine bone marrow-derived mesenchymal stromal cells inhibit reactive oxygen species production by neutrophils. Vet. Immunol. Immunopathol. 2020, 221, 109975. [Google Scholar] [CrossRef]
- Bastos, F.Z.; Barussi, F.C.M.; Leite, L.M.B.; Jamur, V.R.; Soares, A.D.A.; Senegaglia, A.C.; Michelotto, P.V., Jr. Quality control and immunomodulatory potential for clinical-grade equine bone marrow-derived mesenchymal stromal cells and conditioned medium. Res. Vet. Sci. 2020, 132, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Khatab, S.; van Osch, G.J.; Kops, N.; Bastiaansen-Jenniskens, Y.M.; Bos, P.; Verhaar, J.A.; Bernsen, M.R.; van Buul, G.M. Mesenchymal stem cell secretome reduces pain and subchondral bone alterations in a mouse osteoarthritis model. Osteoarthr. Cartil. 2018, 26, S15–S16. [Google Scholar] [CrossRef]
- da Silva Meirelles, L.; Fontes, A.M.; Covas, D.T.; Caplan, A.I. Mechanisms involved in the therapeutic properties of mesenchymal stem cells. Cytokine Growth Factor Rev. 2009, 20, 419–427. [Google Scholar] [CrossRef]
- Lener, T.; Gimona, M.; Aigner, L.; Börger, V.; Buzas, E.; Camussi, G.; Chaput, N.; Chatterjee, D.; Court, F.A.; Del Portillo, H.A.; et al. Applying extracellular vesicles based therapeutics in clinical trials—An ISEV position paper. J. Extracell. Vesicles 2015, 4, 30087. [Google Scholar] [CrossRef] [Green Version]
- Ribitsch, I.; Oreff, G.L.; Jenner, F. Regenerative Medicine for Equine Musculoskeletal Diseases. Animals 2021, 11, 234. [Google Scholar] [CrossRef] [PubMed]
- Keyhanmanesh, R.; Rahbarghazi, R.; Aslani, M.R.; Hassanpour, M.; Ahmadi, M. Systemic delivery of mesenchymal stem cells condition media in repeated doses acts as magic bullets in restoring IFN-γ/IL-4 balance in asthmatic rats. Life Sci. 2018, 212, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Sun, Y.; Gu, X.; Cai, J.; Liu, X.; Zhang, X.; Chen, J.; Hao, Y.; Chen, S. Conditioned medium of human bone marrow-derived stem cells promotes tendon-bone healing of the rotator cuff in a rat model. Biomaterials 2021, 271, 120714. [Google Scholar] [CrossRef] [PubMed]
- Lange-Consiglio, A.; Perrini, C.; Tasquier, R.; Deregibus, M.C.; Camussi, G.; Pascucci, L.; Marini, M.G.; Corradetti, B.; Bizzaro, D.; De Vita, B.; et al. Equine Amniotic Microvesicles and Their Anti-Inflammatory Potential in a Tenocyte Model In Vitro. Stem Cells Dev. 2016, 25, 610–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrini, C.; Strillacci, M.G.; Bagnato, A.; Esposti, P.; Marini, M.G.; Corradetti, B.; Bizzaro, D.; Idda, A.; Ledda, S.; Capra, E.; et al. Microvesicles secreted from equine amniotic-derived cells and their potential role in reducing inflammation in endometrial cells in an in-vitro model. Stem Cell Res. Ther. 2016, 7, 169. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Wang, Q.; Jiang, D. Extracellular vesicles from bone marrow-derived multipotent mesenchymal stromal cells regulate inflammation and enhance tendon healing. J. Transl. Med. 2019, 17, 211. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; He, G.; Guo, Y.; Tang, H.; Shi, Y.; Bian, X.; Zhu, M.; Kang, X.; Zhou, M.; Lyu, J.; et al. Exosomes from tendon stem cells promote injury tendon healing through balancing synthesis and degradation of the tendon extracellular matrix. J. Cell. Mol. Med. 2019, 23, 5475–5485. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Hu, Q.; Song, W.; Yu, W.; He, Y. Adipose Stem Cell–Derived Exosomes Decrease Fatty Infiltration and Enhance Rotator Cuff Healing in a Rabbit Model of Chronic Tears. Am. J. Sports Med. 2020, 48, 1456–1464. [Google Scholar] [CrossRef]
- Jenner, F.; Wagner, A.; Gerner, I.; Ludewig, E.; Trujanovic, R.; Rohde, E.; von Rechenberg, B.; Gimona, M.; Traweger, A. Evaluation of the Potential of Umbilical Cord Mesenchymal Stromal Cell–Derived Small Extracellular Vesicles to Improve Rotator Cuff Healing: A Pilot Ovine Study. Am. J. Sports Med. 2023, 51, 331–342. [Google Scholar] [CrossRef]
- Liu, Y.; Kano, F.; Hashimoto, N.; Xia, L.; Zhou, Q.; Feng, X.; Hibi, H.; Miyazaki, A.; Iwamoto, T.; Matsuka, Y.; et al. Conditioned Medium from the Stem Cells of Human Exfoliated Deciduous Teeth Ameliorates Neuropathic Pain in a Partial Sciatic Nerve Ligation Model. Front. Pharmacol. 2022, 13, 745020. Available online: https://www.frontiersin.org/articles/10.3389/fphar.2022.745020 (accessed on 11 February 2023). [CrossRef]
- Sevivas, N.; Teixeira, F.G.; Portugal, R.; Araújo, L.; Carriço, L.F.; Ferreira, N.; Vieira da Silva, M.; Espregueira-Mendes, J.; Anjo, S.; Manadas, B.; et al. Mesenchymal Stem Cell Secretome: A Potential Tool for the Prevention of Muscle Degenerative Changes Associated with Chronic Rotator Cuff Tears. Am. J. Sports Med. 2016, 45, 179–188. [Google Scholar] [CrossRef]
- Shen, H.; Yoneda, S.; Abu-Amer, Y.; Guilak, F.; Gelberman, R.H. Stem cell-derived extracellular vesicles attenuate the early inflammatory response after tendon injury and repair. J. Orthop. Res. 2020, 38, 117–127. [Google Scholar] [CrossRef]
- Yu, H.; Cheng, J.; Shi, W.; Ren, B.; Zhao, F.; Shi, Y.; Yang, P.; Duan, X.; Zhang, J.; Fu, X.; et al. Bone marrow mesenchymal stem cell-derived exosomes promote tendon regeneration by facilitating the proliferation and migration of endogenous tendon stem/progenitor cells. Acta Biomater. 2020, 106, 328–341. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Kang, X.; Wang, Y.; Bian, X.; He, G.; Zhou, M.; Tang, K. Exosomes Derived from Bone Marrow Stromal Cells (BMSCs) Enhance Tendon-Bone Healing by Regulating Macrophage Polarization. Med. Sci. Monit. 2020, 26, e923328. [Google Scholar] [CrossRef]
- Gissi, C.; Radeghieri, A.; Antonetti Lamorgese Passeri, C.; Gallorini, M.; Calciano, L.; Oliva, F.; Veronesi, F.; Zendrini, A.; Cataldi, A.; Bergese, P.; et al. Extracellular vesicles from rat-bone-marrow mesenchymal stromal/stem cells improve tendon repair in rat Achilles tendon injury model in dose-dependent manner: A pilot study. PLoS ONE 2020, 15, e0229914. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; He, B.; Wang, L.; Yuan, B.; Shu, H.; Zhang, F.; Sun, L. Bone marrow mesenchymal stem cell-derived exosomes promote rotator cuff tendon-bone healing by promoting angiogenesis and regulating M1 macrophages in rats. Stem Cell Res. Ther. 2020, 11, 496. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yao, Z.; Xiong, H.; Cui, H.; Wang, X.; Zheng, W.; Qian, Y.; Fan, C. Extracellular vesicles from hydroxycamptothecin primed umbilical cord stem cells enhance anti-adhesion potential for treatment of tendon injury. Stem Cell Res. Ther. 2020, 11, 500. [Google Scholar] [CrossRef]
- Chen, Q.; Liang, Q.; Zhuang, W.; Zhou, J.; Zhang, B.; Xu, P.; Ju, Y.; Morita, Y.; Luo, Q.; Song, G. Tenocyte proliferation and migration promoted by rat bone marrow mesenchymal stem cell-derived conditioned medium. Biotechnol. Lett. 2018, 40, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Lange-Consiglio, A.; Rossi, D.; Tassan, S.; Perego, R.; Cremonesi, F.; Parolini, O. Conditioned Medium from Horse Amniotic Membrane-Derived Multipotent Progenitor Cells: Immunomodulatory Activity In Vitro and First Clinical Application in Tendon and Ligament Injuries In Vivo. Stem Cells Dev. 2013, 22, 3015–3024. [Google Scholar] [CrossRef] [PubMed]
- Bartekova, M.; Radosinska, J.; Jelemensky, M.; Dhalla, N.S. Role of cytokines and inflammation in heart function during health and disease. Heart Fail. Rev. 2018, 23, 733–758. [Google Scholar] [CrossRef] [PubMed]
- Beohar, N.; Rapp, J.; Pandya, S.; Losordo, D.W. Rebuilding the Damaged Heart: The Potential of Cytokines and Growth Factors in the Treatment of Ischemic Heart Disease. J. Am. Coll. Cardiol. 2010, 56, 1287–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, R.; Mellows, B.; Sheard, J.; Antonioli, M.; Kretz, O.; Chambers, D.; Zeuner, M.-T.; Tomkins, J.E.; Denecke, B.; Musante, L.; et al. Secretome of adipose-derived mesenchymal stem cells promotes skeletal muscle regeneration through synergistic action of extracellular vesicle cargo and soluble proteins. Stem Cell Res. Ther. 2019, 10, 116. [Google Scholar] [CrossRef] [PubMed]
- Wagner, T.; Traxler, D.; Simader, E.; Beer, L.; Narzt, M.-S.; Gruber, F.; Madlener, S.; Laggner, M.; Erb, M.; Vorstandlechner, V.; et al. Different pro-angiogenic potential of γ-irradiated PBMC-derived secretome and its subfractions. Sci. Rep. 2018, 8, 18016. [Google Scholar] [CrossRef] [Green Version]
- Ahrberg, A.B.; Horstmeier, C.; Berner, D.; Brehm, W.; Gittel, C.; Hillmann, A.; Josten, C.; Rossi, G.; Schubert, S.; Winter, K.; et al. Effects of mesenchymal stromal cells versus serum on tendon healing in a controlled experimental trial in an equine model. BMC Musculoskelet. Disord. 2018, 19, 230. [Google Scholar] [CrossRef] [PubMed]
- Preparation of IDEs and INDs for Products Intended to Repair or Replace Knee Cartilage. 2011. Available online: https://www.fda.gov/media/82562/download (accessed on 2 March 2023).
- Reflection Paper on In-Vitro Cultured Chondrocyte Containing Products for Cartilage Repair of the Knee. Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/reflection-paper-vitro-cultured-chondrocyte-containing-products-cartilage-repair-knee_en.pdf (accessed on 2 March 2023).
- Soukup, R.; Gerner, I.; Gültekin, S.; Baik, H.; Oesterreicher, J.; Grillari, J.; Jenner, F. Characterisation of Extracellular Vesicles from Equine Mesenchymal Stem Cells. Int. J. Mol. Sci. 2022, 23, 5858. [Google Scholar] [CrossRef]
- Papait, A.; Ragni, E.; Cargnoni, A.; Vertua, E.; Romele, P.; Masserdotti, A.; Perucca Orfei, C.; Signoroni, P.B.; Magatti, M.; Silini, A.R.; et al. Comparison of EV-free fraction, EVs, and total secretome of amniotic mesenchymal stromal cells for their immunomodulatory potential: A translational perspective. Front. Immunol. 2022, 13, 960909. Available online: https://www.frontiersin.org/articles/10.3389/fimmu.2022.960909 (accessed on 3 March 2023). [CrossRef]
- González-Cubero, E.; González-Fernández, M.L.; Olivera, E.R.; Villar-Suárez, V. Extracellular vesicle and soluble fractions of adipose tissue-derived mesenchymal stem cells secretome induce inflammatory cytokines modulation in an in vitro model of discogenic pain. Spine J. 2022, 22, 1222–1234. [Google Scholar] [CrossRef]
- Giannasi, C.; Niada, S.; Magagnotti, C.; Ragni, E.; Andolfo, A.; Brini, A.T. Comparison of two ASC-derived therapeutics in an in vitro OA model: Secretome versus extracellular vesicles. Stem Cell Res. Ther. 2020, 11, 521. [Google Scholar] [CrossRef]
- Sung, B.H.; Parent, C.A.; Weaver, A.M. Extracellular vesicles: Critical players during cell migration. Dev. Cell 2021, 56, 1861–1874. [Google Scholar] [CrossRef]
- Kriebel, P.W.; Majumdar, R.; Jenkins, L.M.; Senoo, H.; Wang, W.; Ammu, S.; Chen, S.; Narayan, K.; Iijima, M.; Parent, C.A. Extracellular vesicles direct migration by synthesizing and releasing chemotactic signals. J. Cell Biol. 2018, 217, 2891–2910. [Google Scholar] [CrossRef]
- Voga, M.; Adamic, N.; Vengust, M.; Majdic, G. Stem Cells in Veterinary Medicine-Current State and Treatment Options. Front. Vet. Sci. 2020, 7, 278. [Google Scholar] [CrossRef]
- Margolis, L.; Sadovsky, Y. The biology of extracellular vesicles: The known unknowns. PLoS Biol. 2019, 17, e3000363. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lai, Y.; Hua, Z.C. Apoptosis and apoptotic body: Disease message and therapeutic target potentials. Biosci. Rep. 2019, 39, BSR20180992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iraci, N.; Leonardi, T.; Gessler, F.; Vega, B.; Pluchino, S. Focus on Extracellular Vesicles: Physiological Role and Signalling Properties of Extracellular Membrane Vesicles. Int. J. Mol. Sci. 2016, 17, 171. [Google Scholar] [CrossRef] [Green Version]
- Niada, S.; Giannasi, C.; Magagnotti, C.; Andolfo, A.; Brini, A.T. Proteomic analysis of extracellular vesicles and conditioned medium from human adipose-derived stem/stromal cells and dermal fibroblasts. J. Proteom. 2021, 232, 104069. [Google Scholar] [CrossRef]
- Niada, S.; Giannasi, C.; Gualerzi, A.; Banfi, G.; Brini, A.T. Differential Proteomic Analysis Predicts Appropriate Applications for the Secretome of Adipose-Derived Mesenchymal Stem/Stromal Cells and Dermal Fibroblasts. Stem Cells Int. 2018, 2018, 7309031. [Google Scholar] [CrossRef] [Green Version]
- Monguió-Tortajada, M.; Gálvez-Montón, C.; Bayes-Genis, A.; Roura, S.; Borràs, F.E. Extracellular vesicle isolation methods: Rising impact of size-exclusion chromatography. Cell. Mol. Life Sci. 2019, 76, 2369–2382. [Google Scholar] [CrossRef]
- Van Deun, J.; Mestdagh, P.; Sormunen, R.; Cocquyt, V.; Vermaelen, K.; Vandesompele, J.; Bracke, M.; De Wever, O.; Hendrix, A. The impact of disparate isolation methods for extracellular vesicles on downstream RNA profiling. J. Extracell. Vesicles 2014, 3, 24858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, D.B.; Gray, K.M.; Santharam, Y.; Lamichhane, T.N.; Stroka, K.M.; Jay, S.M. Impact of cell culture parameters on production and vascularization bioactivity of mesenchymal stem cell-derived extracellular vesicles. Bioeng. Transl. Med. 2017, 2, 170–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miclau, K.; Hambright, W.S.; Huard, J.; Stoddart, M.J.; Bahney, C.S. Cellular expansion of MSCs: Shifting the regenerative potential. Aging Cell 2023, 22, e13759. [Google Scholar] [CrossRef]
- Gualerzi, A.; Niada, S.; Giannasi, C.; Picciolini, S.; Morasso, C.; Vanna, R.; Rossella, V.; Masserini, M.; Bedoni, M.; Ciceri, F.; et al. Raman spectroscopy uncovers biochemical tissue-related features of extracellular vesicles from mesenchymal stromal cells. Sci. Rep. 2017, 7, 9820. [Google Scholar] [CrossRef]
- Shin, S.; Lee, J.; Kwon, Y.; Park, K.S.; Jeong, J.H.; Choi, S.J.; Bang, S.I.; Chang, J.W.; Lee, C. Comparative proteomic analysis of the mesenchymal stem cells secretome from adipose, bone marrow, placenta and wharton’s jelly. Int. J. Mol. Sci. 2021, 22, 845. [Google Scholar] [CrossRef]
- Uberti, B.; Plaza, A.; Henríquez, C. Pre-conditioning Strategies for Mesenchymal Stromal/Stem Cells in Inflammatory Conditions of Livestock Species. Front. Vet. Sci. 2022, 9, 806069. Available online: https://www.frontiersin.org/articles/10.3389/fvets.2022.806069 (accessed on 8 March 2023). [CrossRef] [PubMed]
- Zhao, L.; Hu, C.; Han, F.; Cai, F.; Wang, J.; Chen, J. Preconditioning is an effective strategy for improving the efficiency of mesenchymal stem cells in kidney transplantation. Stem Cell Res. Ther. 2020, 11, 197. [Google Scholar] [CrossRef]
- Han, K.-H.; Kim, A.-K.; Kim, M.-H.; Kim, D.-H.; Go, H.-N.; Kim, D.-I. Enhancement of angiogenic effects by hypoxia-preconditioned human umbilical cord-derived mesenchymal stem cells in a mouse model of hindlimb ischemia. Cell Biol. Int. 2016, 40, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Matta, A.; Nader, V.; Lebrin, M.; Gross, F.; Prats, A.C.; Cussac, D.; Galinier, M.; Roncalli, J. Pre-Conditioning Methods and Novel Approaches with Mesenchymal Stem Cells Therapy in Cardiovascular Disease. Cells 2022, 11, 1620. [Google Scholar] [CrossRef]
- Carlomagno, C.; Giannasi, C.; Niada, S.; Bedoni, M.; Gualerzi, A.; Brini, A.T. Raman Fingerprint of Extracellular Vesicles and Conditioned Media for the Reproducibility Assessment of Cell-Free Therapeutics. Front. Bioeng. Biotechnol. 2021, 9, 640617. Available online: https://www.frontiersin.org/articles/10.3389/fbioe.2021.640617 (accessed on 8 March 2023). [CrossRef]
- He, C.; Dai, M.; Zhou, X.; Long, J.; Tian, W.; Yu, M. Comparison of two cell-free therapeutics derived from adipose tissue: Small extracellular vesicles versus conditioned medium. Stem Cell Res. Ther. 2022, 13, 86. [Google Scholar] [CrossRef]
- Crum, R.J.; Capella-Monsonís, H.; Badylak, S.F.; Hussey, G.S. Extracellular Vesicles for Regenerative Medicine Applications. Appl. Sci. 2022, 12, 7472. [Google Scholar] [CrossRef]
- Helissey, C.; Guitard, N.; Théry, H.; Goulinet, S.; Mauduit, P.; Girleanu, M.; Favier, A.L.; Drouet, M.; Parnot, C.; Chargari, C.; et al. Two New Potential Therapeutic Approaches in Radiation Cystitis Derived from Mesenchymal Stem Cells: Extracellular Vesicles and Conditioned Medium. Biology 2022, 11, 980. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Chen, X.; Pan, Q.; Wang, Y.; Su, S.; Jiang, C.; Li, Y.; Xu, N.; Wu, L.; Lou, X.; et al. A Comprehensive Proteomics Analysis Reveals a Secretory Path- and Status-Dependent Signature of Exosomes Released from Tumor-Associated Macrophages. J. Proteome Res. 2015, 14, 4319–4331. [Google Scholar] [CrossRef] [PubMed]
- Eddershaw, P.J.; Beresford, A.P.; Bayliss, M.K. ADME/PK as part of a rational approach to drug discovery. Drug Discov. Today 2000, 5, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Charoenviriyakul, C.; Takahashi, Y.; Morishita, M.; Matsumoto, A.; Nishikawa, M.; Takakura, Y. Cell type-specific and common characteristics of exosomes derived from mouse cell lines: Yield, physicochemical properties, and pharmacokinetics. Eur. J. Pharm. Sci. 2017, 96, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Smyth, T.; Kullberg, M.; Malik, N.; Smith-Jones, P.; Graner, M.W.; Anchordoquy, T.J. Biodistribution and delivery efficiency of unmodified tumor-derived exosomes. J. Control. Release 2015, 199, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-R.; Kim, J.-W.; Jun, H.-S.; Roh, J.Y.; Lee, H.-Y.; Hong, I.-S. Stem Cell Secretome and Its Effect on Cellular Mechanisms Relevant to Wound Healing. Mol. Ther. 2018, 26, 606–617. [Google Scholar] [CrossRef] [Green Version]
- Delling, U.; Lindner, K.; Ribitsch, I.; Jülke, H.; Brehm, W. Comparison of bone marrow aspiration at the sternum and the tuber coxae in middle-aged horses. Can. J. Vet. Res. 2012, 76, 52–56. [Google Scholar]
- Ribitsch, I.; Chang-Rodriguez, S.; Egerbacher, M.; Gabner, S.; Gueltekin, S.; Huber, J.; Schuster, T.; Jenner, F. Sheep Placenta Cotyledons: A Noninvasive Source of Ovine Mesenchymal Stem Cells. Tissue Eng. Part C Methods 2017, 23, 298–310. [Google Scholar] [CrossRef]
- Cammack, R.; Atwood, T.; Campbell, P.; Parish, H.; Smith, A.; Vella, F.; Stirling, J. Oxford Dictionary of Biochemistry and Molecular Biology; Oxford University Press: Oxford, UK, 2008; ISBN 9780191727641. [Google Scholar]
- Karnieli, O.; Friedner, O.M.; Allickson, J.G.; Zhang, N.; Jung, S.; Fiorentini, D.; Abraham, E.; Eaker, S.S.; Yong, T.K.; Chan, A.; et al. A consensus introduction to serum replacements and serum-free media for cellular therapies. Cytotherapy 2017, 19, 155–169. [Google Scholar] [CrossRef]
- European Commission. Note for guidance on minimising the risk of transmitting animal spongiform encephalopathy agents via human and veterinary medicinal products (EMA/410/01 rev.3). Off. J. Eur. Union 2011, C, 73/01. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:52011XC0305(04) (accessed on 10 March 2023).
- Mendicino, M.; Bailey, A.M.; Wonnacott, K.; Puri, R.K.; Bauer, S.R. MSC-Based Product Characterization for Clinical Trials: An FDA Perspective. Cell Stem Cell 2014, 14, 141–145. [Google Scholar] [CrossRef] [Green Version]
- Oesterreicher, J.; Pultar, M.; Schneider, J.; Mühleder, S.; Zipperle, J.; Grillari, J.; Holnthoner, W. Fluorescence-Based Nanoparticle Tracking Analysis and Flow Cytometry for Characterization of Endothelial Extracellular Vesicle Release. Int. J. Mol. Sci. 2020, 21, 9278. [Google Scholar] [CrossRef]
- Oreff, G.L.; Fenu, M.; Vogl, C.; Ribitsch, I.; Jenner, F. Species variations in tenocytes’ response to inflammation require careful selection of animal models for tendon research. Sci. Rep. 2021, 11, 12451. [Google Scholar] [CrossRef] [PubMed]
- Kugo, H.; Sukketsiri, W.; Iwamoto, K.; Suihara, S.; Moriyama, T.; Zaima, N. Low glucose and serum levels cause an increased inflammatory factor in 3T3-L1 cell through Akt, MAPKs and NF-κB activation. Adipocyte 2021, 10, 232–241. [Google Scholar] [CrossRef]
- Kogel, V.; Trinh, S.; Gasterich, N.; Beyer, C.; Seitz, J. Long-Term Glucose Starvation Induces Inflammatory Responses and Phenotype Switch in Primary Cortical Rat Astrocytes. J. Mol. Neurosci. 2021, 71, 2368–2382. [Google Scholar] [CrossRef] [PubMed]
- van Vijven, M.; Wunderli, S.L.; Ito, K.; Snedeker, J.G.; Foolen, J. Serum deprivation limits loss and promotes recovery of tenogenic phenotype in tendon cell culture systems. J. Orthop. Res. 2021, 39, 1561–1571. [Google Scholar] [CrossRef]
- Mueller, A.J.; Tew, S.R.; Vasieva, O.; Clegg, P.D.; Canty-Laird, E.G. A systems biology approach to defining regulatory mechanisms for cartilage and tendon cell phenotypes. Sci. Rep. 2016, 6, 33956. [Google Scholar] [CrossRef] [Green Version]
- Brink, H.E.; Miller, G.J.; Beredjiklian, P.K.; Nicoll, S.B. Serum-dependent effects on adult and fetal tendon fibroblast migration and collagen expression. Wound Repair Regen. 2006, 14, 179–186. [Google Scholar] [CrossRef]
- Anders, S. Babraham Bioinformatics—FastQC A Quality Control tool for High Throughput Sequence Data. Soil. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 10 March 2023).
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. Subgroup, 1000 Genome Project Data Processing the Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Zerbino, D.R.; Achuthan, P.; Akanni, W.; Amode, M.R.; Barrell, D.; Bhai, J.; Billis, K.; Cummins, C.; Gall, A.; Girón, C.G.; et al. Ensembl 2018. Nucleic Acids Res. 2018, 46, D754–D761. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Coenye, T. Do results obtained with RNA-sequencing require independent verification? Biofilm 2021, 3, 100043. [Google Scholar] [CrossRef]
- Feng, L.; Liu, H.; Liu, Y.; Lu, Z.; Guo, G.; Guo, S.; Zheng, H.; Gao, Y.; Cheng, S.; Wang, J.; et al. Power of Deep Sequencing and Agilent Microarray for Gene Expression Profiling Study. Mol. Biotechnol. 2010, 45, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Hänzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-Seq data. BMC Bioinformatics 2013, 14, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database Hallmark Gene Set Collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Martens, M.; Ammar, A.; Riutta, A.; Waagmeester, A.; Slenter, D.N.; Hanspers, K.; Miller, R.A.; Digles, D.; Lopes, E.N.; Ehrhart, F.; et al. WikiPathways: Connecting communities. Nucleic Acids Res. 2021, 49, D613–D621. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating viruses and cellular organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [Green Version]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goedhart, J.; Luijsterburg, M.S. VolcaNoseR is a web app for creating, exploring, labeling and sharing volcano plots. Sci. Rep. 2020, 10, 20560. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CM | EV | PF | |
|---|---|---|---|
| Protein (µg/mL) | 130 ± 6.18 | 30.4 ± 0.5 | 73.2 ± 0.9 |
| Particle number (total NTA) (particles/mL) | 1.53 × 1011 ± 2.01 × 1010 | 1.03 × 1011 ± 2.04 × 1010 | Not detected |
| Particle number ≤ 195 nm (NTA) (particles/mL) | 5.36 × 1010 ± 7.67 × 109 | 5.61 × 1010 ± 1.06 × 1010 | Not detected |
| Particle number > 195 nm (NTA) (particles/mL) | 1.09 × 1011 ± 2.51 × 1010 | 4.67 × 1010 ± 9.81 × 109 | Not detected |
| Top 10 Downregulated Genes in the Treatment/Healthy CTRL vs. Inflamed CTRL (Sorted by Ascending Adjusted p-Value) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CM | EV | PF | Healthy CTRL | ||||||||
| Gene | FC | Adj. p-Value | Gene | FC | Adj. p-Value | Gene | FC | Adj. p-Value | Gene | FC | Adj. p-Value |
| ADGRD1 | −23.750 | 9.5536 × 10−7 | FUZ | −36.203 | 3.0566 × 10−16 | NOXA1 | −40.887 | 6.1398 × 10−20 | PSME2 | 2.11877 | 2.5995 × 10−40 |
| SAA | −31.261 | 5.7985 × 10−6 | NTN1 | −30.152 | 0.00027148 | TRIM6 | −34.742 | 3.7594 × 10−17 | KYAT3 | 1.86235 | 2.2722 × 10−35 |
| HIVEP3 | −15.446 | 1.0763 × 10−5 | GPM6B | −29.649 | 0.00027148 | H1-9 | −30.859 | 4.3641 × 10−8 | RAB20 | 3.90099 | 2.2722 × 10−35 |
| COL14A1 | −28.138 | 9.163 × 10−5 | podocalyxin like | −28.229 | 0.01057581 | TCP11 | −33.531 | 1.2429 × 10−5 | DBNL | 2.24989 | 9.5604 × 10−34 |
| ZIC2 | −12.980 | 0.00011 | LRRC23 | −27.298 | 0.01865918 | MRPL54 | −31.808 | 2.4525 × 10−5 | SNAP29 | 1.72103 | 5.1924 × 10−32 |
| VSIR | −17.357 | 0.00038 | C6orf47 | −21.264 | 0.03612666 | ZNF382 | −17.616 | 5.3998 × 10−5 | CCL2 | 4.07345 | 3.5555 × 10−30 |
| FBXO32 | −16.372 | 0.00056 | CMYA5 | −18.940 | 1.5926 × 10−7 | FAM216B | −16.926 | 0.00018 | CCL11 | 3.49896 | 6.2286 × 10−30 |
| PLA2G2A | −26.155 | 0.00062 | ZKSCAN8 | −17.918 | 0.00027148 | NTN1 | −29.748 | 0.00031 | PI3 | 3.77388 | 2.2449 × 10−29 |
| SCARA5 | −43.596 | 0.00067 | CSPG4 | −17.519 | 1.3391 × 10−5 | CCDC40 | −26.832 | 0.00257 | CCL5 | 5.21572 | 4.8974 × 10−29 |
| SPNS3 | −31.756 | 0.00205 | GSK-3 | −17.2511 | 0.00031 | IL17RA | −17.499 | 0.00443 | SIDT2 | 3.70467 | 3.1246 × 10−27 |
| Top 10 Upregulated Genes in the Treatment/Healthy CTRL vs. Inflamed CTRL (Sorted by Ascending Adjusted p-Value) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CM | EV | PF | Healthy CTRL | ||||||||
| Gene | FC | Adj. p-Value | Gene | FC | Adj. p-Value | Gene | FC | Adj. p-Value | Gene | FC | Adj. p-Value |
| PIRT | 186.866 | 9.78 × 10−10 | ZNF582 | 443.953 | 7.7528 × 10−46 | ZNF582 | 432.963 | 9.88 × 10−43 | ZNF582 | −45.537 | 8.3178 × 10−50 |
| ELMO1 | 181.622 | 3.61 × 10−10 | PIRT | 197.946 | 6.2092 × 10−12 | PIRT | 186.640 | 3.19 × 10−10 | ENSECAG00000016730 | −28.492 | 5.4407 × 10−42 |
| ZNF501 | 239.838 | 3.61 × 10−7 | ELMO1 | 173.926 | 1.6926 × 10−6 | CCND1 | 198.841 | 3.53 × 10−7 | CCDC88A | −27.159 | 1.195 × 10−32 |
| ENSECAG00000023978 | 184.840 | 1.75 × 10−6 | ZNF501 | 226.906 | 1.9717 × 10−6 | metallothionein-1A | 451.660 | 3.53 × 10−7 | CELF2 | −28.702 | 6.2556 × 10−29 |
| TPGS2 | 173.051 | 4.13 × 10−6 | ENSECAG00000023978 | 174.590 | 1.4739 × 10−5 | TPGS2 | 178.046 | 8.18 × 10−7 | FSTL1 | −32.601 | 1.8517 × 10−25 |
| ENSECAG00000013505 | 182.588 | 7.55 × 10−6 | ZNF182 | 286.245 | 1.6141 × 10−5 | ELMO1 | 167.381 | 4.86 × 10−6 | PTMA | −26.727 | 1.2088 × 10−21 |
| STAC | 215.231 | 3.03 × 10−5 | ENSECAG00000013505 | 160.195 | 0.00035 | ZNF182 | 292.836 | 7.23 × 10−6 | CALCA | −47.284 | 2.7129 × 10−21 |
| CCND1 | 163.745 | 0.00012 | DCX | 205.337 | 0.00060 | ZNF501 | 214.091 | 1.24 × 10−5 | C7H11orf58 | −29.221 | 4.2637 × 10−20 |
| RAD51D | 210.350 | 0.00038 | ASB5 | 161.541 | 0.00447 | MT1B | 163.702 | 0.00018 | ANXA5 | −41.201 | 2.5081 × 10−19 |
| ASB5 | 176.871 | 0.00038 | GRID1 | 273.429 | 0.01865 | ACBD7 | 319.023 | 0.00020 | APC | −17.720 | 1.6149 × 10−17 |
| Downregulated Pathways | |||||
|---|---|---|---|---|---|
| CM | EV | ||||
| Pathway | FC | Adj. p-Value | Pathway | FC | Adj. p-Value |
| Secretion of hydrochloric acid in parietal cells | −1.1589254 | 0.09355804 | Glycerolipids_and_glycerophospholipids | −0.6118861 | 0.06337278 |
| Nephrogenesis | −1.0681281 | 0.01124854 | Myd88 distinct input/output pathway | −0.5034747 | 0.06337278 |
| Fatty acid omegaoxidation | −1.0049143 | 0.07476631 | Autosomal recessive osteopetrosis pathways | −0.5445 | 0.06826365 |
| Complement activation | −0.7625858 | 0.07859295 | |||
| Interleukin1 induced activation of nfkb | −0.674886 | 0.03094377 | |||
| Tryptophan metabolism | −0.6162714 | 0.04539786 | |||
| Mammalian disorder of sexual development | −0.6081617 | 0.03202702 | |||
| Biomarkers for urea cycle disorders | −0.5883797 | 0.05081052 | |||
| Cysteine and methionine catabolism | −0.5820551 | 0.04634927 | |||
| Oxidative stress response | −0.5797825 | 0.09005984 | |||
| Eicosanoid synthesis | −0.5773651 | 0.03997058 | |||
| Overview of nanoparticle effects | −0.5647847 | 0.03762172 | |||
| Beta alanine metabolism | −0.5222863 | 0.09005984 | |||
| Matrix metalloproteinases | −0.4351830 | 0.07476631 | |||
| S1P receptor signal transduction | −0.4120393 | 0.05402778 | |||
| Autosomal recessive osteopetrosis pathways | −0.4056264 | 0.07915106 | |||
| Valine leucine and isoleucine degradation | −0.3839604 | 0.08790938 | |||
| Sarscov2 innate immunity evasion and cellspecific immune response | −0.3664584 | 0.04539786 | |||
| Selenium micronutrient network | −0.3640307 | 0.05698454 | |||
| Onecarbon metabolism and related pathways | 0.31911033 | 0.08974443 | |||
| Leukocyte transendothelial migration | −0.2761994 | 0.07915106 | |||
| Upregulated Pathways | ||
|---|---|---|
| CM | ||
| Pathway | FC | Adj. p-Value |
| Nucleotide excision repair in xeroderma pigmentosum | 0.3494815 | 0.07130188 |
| Pyrimidine metabolism | 0.37184491 | 0.08655644 |
| Dna repair pathways full network | 0.40690974 | 0.04539786 |
| Oocyte meiosis | 0.40797081 | 0.03997058 |
| Nucleotide excision repair (WP) | 0.42922411 | 0.03776133 |
| Nucleotide excision repair (KEGG) | 0.42922411 | 0.03776133 |
| Progeriaassociated lipodystrophy | 0.4650666 | 0.04539786 |
| Pyrimidine metabolism | 0.47735464 | 0.03202702 |
| Cell cycle (WP) | 0.49283852 | 0.05915343 |
| Cell cycle (KEGG) | 0.4933381 | 0.05577275 |
| Gastric cancer network 1 | 0.49568256 | 0.09520106 |
| DNA irdamage and cellular response via ATR | 0.50092208 | 0.04101562 |
| Base excision repair | 0.55896011 | 0.09005984 |
| Cohesin complex cornelia de lange syndrome | 0.58300193 | 0.01124854 |
| Mammary gland development pathway puberty stage 2 of 4 | 0.58799809 | 0.04539786 |
| Base excision repair | 0.59293411 | 0.09955858 |
| Gastric cancer network 2 | 0.67347864 | 0.04101562 |
| ATR signaling | 0.68398751 | 0.07272085 |
| Homologous recombination | 0.70623154 | 0.04539786 |
| Retinoblastoma gene in cancer | 0.71687919 | 0.03776133 |
| Nucleotide metabolism | 0.76157908 | 0.04101562 |
| Serine metabolism | 0.76326554 | 0.01124854 |
| Mismatch repair | 0.81963998 | 0.03202702 |
| Regulation of sister chromatid separation at the metaphaseanaphase transition | 0.86498193 | 0.03856315 |
| DNA replication | 0.904723 | 0.03094377 |
| DNA mismatch repair | 0.92494463 | 0.03202702 |
| Acquired partial lipodystrophy barraquersimons syndrome | 0.93639734 | 0.03202702 |
| DNA replication | 0.96512563 | 0.03202702 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soukup, R.; Gerner, I.; Mohr, T.; Gueltekin, S.; Grillari, J.; Jenner, F. Mesenchymal Stem Cell Conditioned Medium Modulates Inflammation in Tenocytes: Complete Conditioned Medium Has Superior Therapeutic Efficacy than Its Extracellular Vesicle Fraction. Int. J. Mol. Sci. 2023, 24, 10857. https://doi.org/10.3390/ijms241310857
Soukup R, Gerner I, Mohr T, Gueltekin S, Grillari J, Jenner F. Mesenchymal Stem Cell Conditioned Medium Modulates Inflammation in Tenocytes: Complete Conditioned Medium Has Superior Therapeutic Efficacy than Its Extracellular Vesicle Fraction. International Journal of Molecular Sciences. 2023; 24(13):10857. https://doi.org/10.3390/ijms241310857
Chicago/Turabian StyleSoukup, Robert, Iris Gerner, Thomas Mohr, Sinan Gueltekin, Johannes Grillari, and Florien Jenner. 2023. "Mesenchymal Stem Cell Conditioned Medium Modulates Inflammation in Tenocytes: Complete Conditioned Medium Has Superior Therapeutic Efficacy than Its Extracellular Vesicle Fraction" International Journal of Molecular Sciences 24, no. 13: 10857. https://doi.org/10.3390/ijms241310857