
A Novel Strategy for Screening Tumor-Specific Variable Domain of Heavy-Chain Antibodies

Abstract
:1. Introduction
2. Results
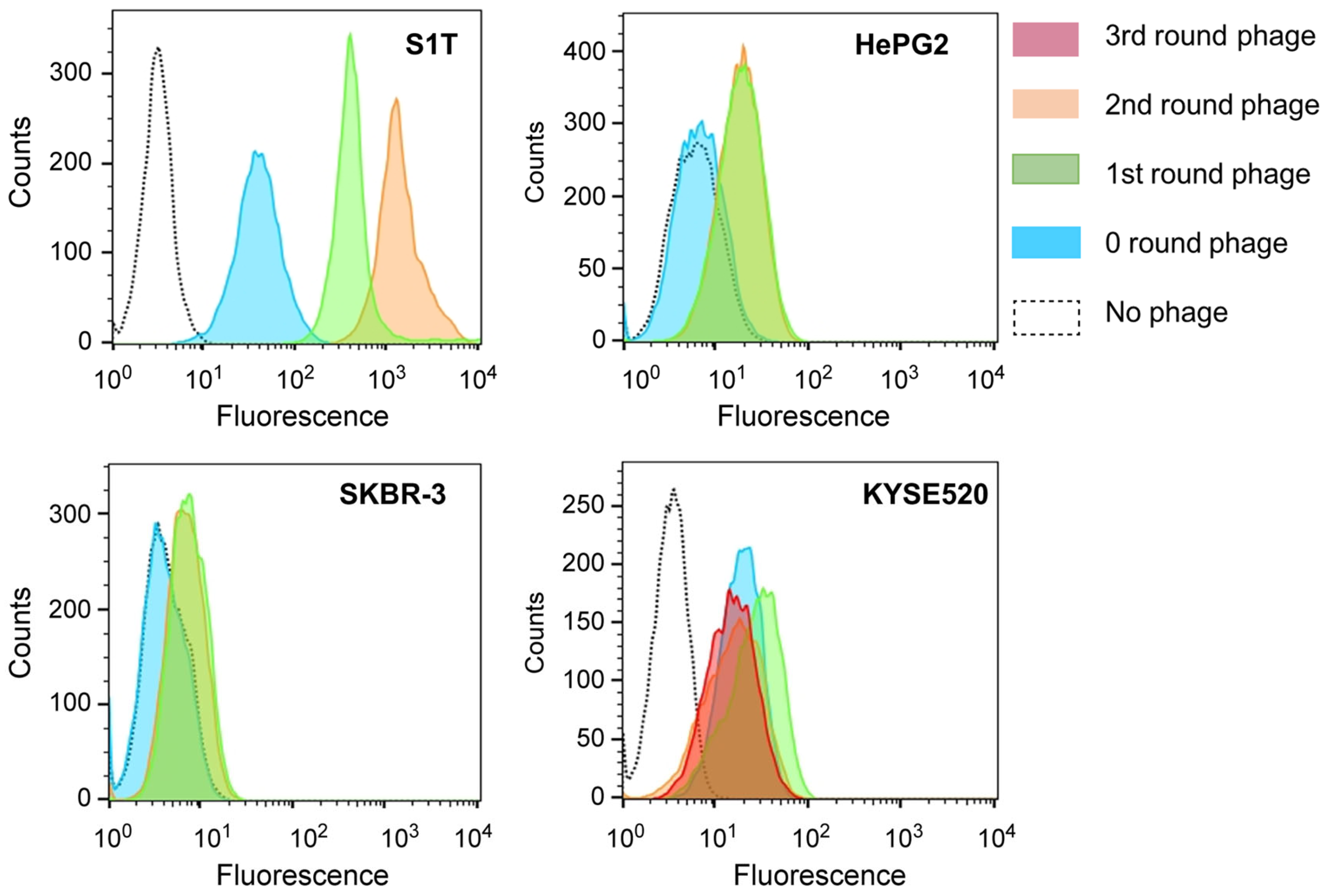
2.1. Construction of Phage Libraries and Enrichment of Phages through Cell Panning
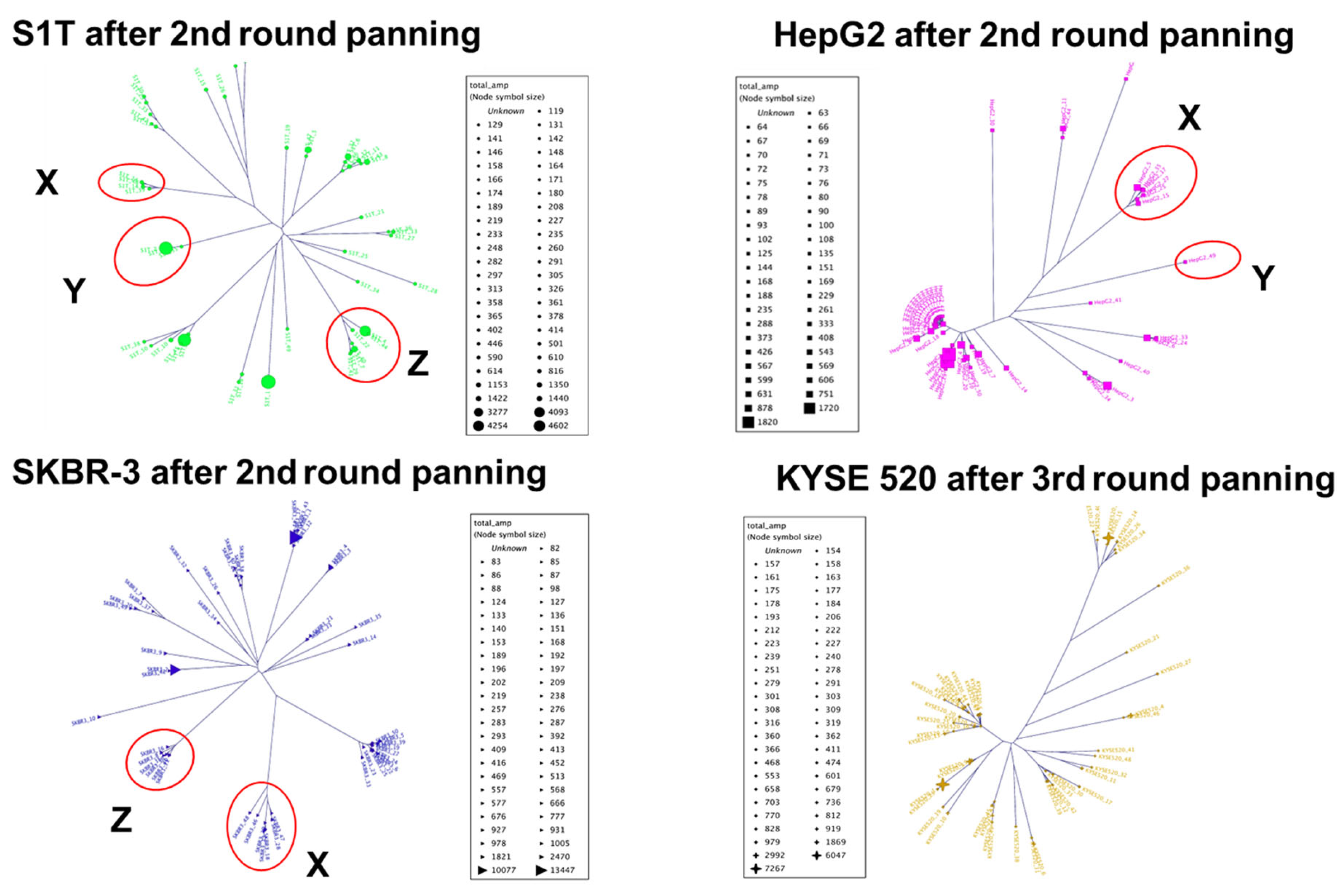
2.2. High-Throughput Sequencing Analysis
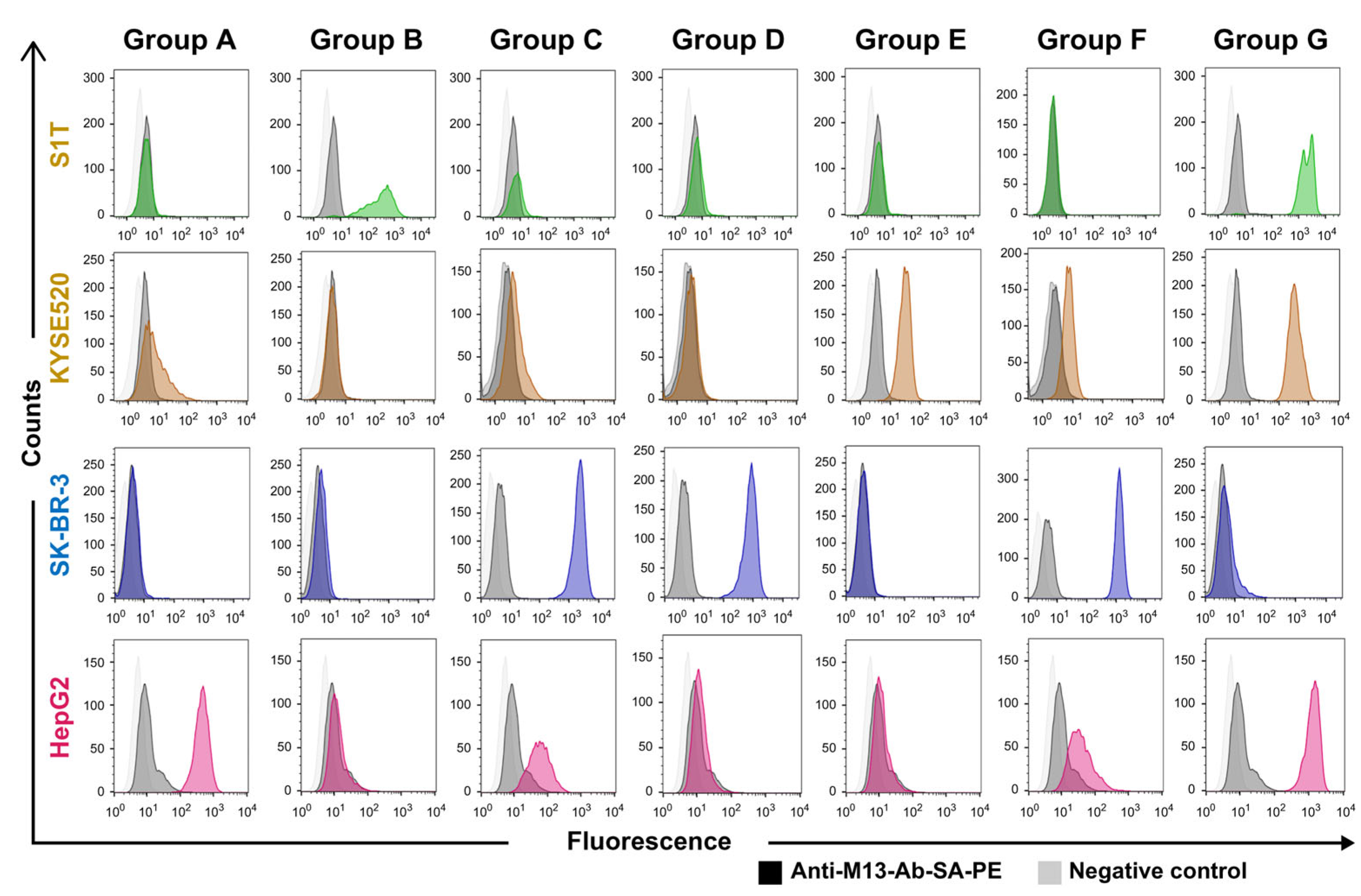
2.3. Binding Properties of NGS Identified Groups of VHH Clones
3. Discussion
4. Materials and Methods
4.1. Alpaca Immunization
4.2. Construction of Tumor Cell-Specific VHH Phage Library
4.3. Phage Propagation
4.4. Phage Library Enrichment by Cell Sorter
4.5. Enriched Phage Binding with Tumor-Specific Cells by FACS
4.6. NGS Sample Preparation and Analysis
4.7. Regeneration of NGS-Identified Clones and Binding Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef]
- de Martel, C.; Georges, D.; Bray, F.; Ferlay, J.; Clifford, G.M. Global burden of cancer attributable to infections in 2018: A worldwide incidence analysis. Lancet Glob. Health 2020, 8, e180–e190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplon, H.; Muralidharan, M.; Schneider, Z.; Reichert, J.M. Antibodies to watch in 2020. mAbs 2020, 12, 1703531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitamura, T.; Dong, P.; Ihira, K.; Kudo, M.; Watari, H. Molecular-targeted therapies and precision medicine for endometrial cancer. Jpn. J. Clin. Oncol. 2019, 49, 108–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhang, Z. The history and advances in cancer immunotherapy: Understanding the characteristics of tumor-infiltrating immune cells and their therapeutic implications. Cell. Mol. Immunol. 2020, 17, 807–821. [Google Scholar] [CrossRef]
- Diver, E.J.; Foster, R.; Rueda, B.R.; Growdon, W.B. The therapeutic challenge of targeting HER2 in endometrial cancer. Oncologist 2015, 20, 1058–1068. [Google Scholar] [CrossRef] [Green Version]
- Ratner, L. Biomarkers and preclinical models for adult T-cell leukemia-lymphoma treatment. Front. Microbiol. 2019, 10, 2109. [Google Scholar] [CrossRef]
- Melo, R.C.C.; Longhini, A.L.; Bigarella, C.L.; Baratti, M.O.; Traina, F.; Favaro, P.; de Melo Campos, P.D.M.; Saad, S.T.O. CXCR7 is highly expressed in acute lymphoblastic leukemia and potentiates CXCR4 response to CXCL12. PLoS ONE 2014, 9, e85926. [Google Scholar] [CrossRef]
- Toboni, M.D.; Mutch, D.G. The emerging role of precision medicine in the treatment of endometrial cancer. Expert Rev. Precis. Med. Drug Dev. 2020, 5, 87–93. [Google Scholar] [CrossRef]
- Pérez, J.M.J.; Renisio, J.G.; Prompers, J.J.; Van Platerink, C.J.; Cambillau, C.; Darbon, H.; Frenken, L.G.J. Thermal unfolding of a llama antibody fragment: A two-state reversible process. Biochemistry 2001, 40, 74–83. [Google Scholar] [CrossRef]
- Ladenson, R.C.; Crimmins, D.L.; Landt, Y.; Ladenson, J.H. Isolation and characterization of a thermally stable recombinant anti-caffeine heavy-chain antibody fragment. Anal. Chem. 2006, 78, 4501–4508. [Google Scholar] [CrossRef]
- Omidfar, K.; Rasaee, M.J.; Kashanian, S.; Paknejad, M.; Bathaie, Z. Studies of thermostability in Camelus bactrianus (Bactrian Camel) single-domain antibody specific for the mutant epidermal-growth-factor receptor expressed by pichia. Biotechnol. Appl. Biochem. 2007, 46, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Ewert, S.; Cambillau, C.; Conrath, K.; Plückthun, A. Biophysical properties of Camelid v HH domains compared to those of human V(H)3 domains. Biochemistry 2002, 41, 3628–3636. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, J.W.; Yang, L.; De Plano, L.M.; Stackhouse, M.A.; Petrenko, V.A. Evolution of a landscape phage library in a mouse xenograft model of human breast cancer. Viruses 2019, 11, 988. [Google Scholar] [CrossRef] [Green Version]
- Rafique, A.; Satake, K.; Kishimoto, S.; Hasan Khan, K.; Kato, D.I.; Ito, Y. Efficient screening and design of variable domain of heavy chain antibody ligands through high throughput sequencing for affinity chromatography to purify Fab fragments. Monoclon. Antib. Immunodiagn. Immunother. 2019, 38, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.L.; Alfaleh, M.A.; Kumble, S.; Zhang, S.; Osborne, G.W.; Yeh, M.; Arora, N.; Hou, J.J.C.; Howard, C.B.; Chin, D.Y.; et al. Targeting membrane proteins for antibody discovery using phage display. Sci. Rep. 2016, 6, 26240. [Google Scholar] [CrossRef] [Green Version]
- Reddy, S.T.; Ge, X.; Miklos, A.E.; Hughes, R.A.; Kang, S.H.; Hoi, K.H.; Chrysostomou, C.; Hunicke-Smith, S.P.; Iverson, B.L.; Tucker, P.W.; et al. Monoclonal antibodies isolated without screening by analyzing the variable-gene repertoire of plasma cells. Nat. Biotechnol. 2010, 28, 965–969. [Google Scholar] [CrossRef]
- Zambrano, N.; Froechlich, G.; Lazarevic, D.; Passariello, M.; Nicosia, A.; De Lorenzo, C.; Morelli, M.J.; Sasso, E. High-throughput monoclonal antibody discovery from phage libraries: Challenging the current preclinical pipeline to keep the pace with the increasing MAb demand. Cancers 2022, 14, 1325. [Google Scholar] [CrossRef]
- Wiedmann, K.H.; Trejdosiewicz, L.K.; Southgate, J.; Thomas, H.C. Human hepatocellular carcinoma: Cross-reactive and idiotypic antigens associated with malignant transformation of epithelial cells. Hepatology 1987, 7, 543–550. [Google Scholar] [CrossRef]
- Zhu, J.; Ofek, G.; Yang, Y.; Zhang, B.; Louder, M.K.; Lu, G.; McKee, K.; Pancera, M.; Skinner, J.; Zhang, Z.; et al. Mining the antibodyome for HIV-1-neutralizing antibodies with next-generation sequencing and phylogenetic pairing of heavy/light chains. Proc. Natl Acad. Sci. USA 2013, 110, 6470–6475. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, A.; Kringelum, J.V.; Hansen, C.S.; Bøgh, K.L.; Sullivan, E.; Patel, J.; Rigby, N.M.; Eiwegger, T.; Szépfalusi, Z.; Masi, F.D.; et al. High-throughput sequencing enhanced phage display enables the identification of patient-specific epitope motifs in serum. Sci. Rep. 2015, 5, 12913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Weber, G.F. Quantitative analysis of protein evolution: The phylogeny of osteopontin. Front. Genet. 2021, 12, 700789. [Google Scholar] [CrossRef] [PubMed]
- Falzone, L.; Salomone, S.; Libra, M. Evolution of cancer pharmacological treatments at the turn of the third millennium. Front. Pharmacol. 2018, 9, 1300. [Google Scholar] [CrossRef] [Green Version]
- Croswell, J.M.; Ransohoff, D.F.; Kramer, B.S. Principles of cancer screening: Lessons from history and study design issues. Semin. Oncol. 2010, 37, 202–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, H.; Khatami, M. Analyses of repeated failures in cancer therapy for solid tumors: Poor tumor-selective drug delivery, low therapeutic efficacy and unsustainable costs. Clin. Transl. Med. 2018, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Allen, T.M.; Cleland, L.G. Serum-induced leakage of liposome contents. Biochim. Biophys. Acta 1980, 597, 418–426. [Google Scholar] [CrossRef]
- Chioccioli, M.; Hankamer, B.; Ross, I.L. Flow cytometry pulse width data enables rapid and sensitive estimation of biomass dry weight in the microalgae Chlamydomonas reinhardtii and Chlorella vulgaris. PLoS ONE 2014, 9, e97269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, A.I.; Canuso, C.M.; Brenner, M.J.; Wojcik, J.D. Detection and management of comorbidity in patients with schizophrenia. Psychiatr. Clin. N. Am. 2003, 26, 115–139. [Google Scholar] [CrossRef]
- Buckler, D.R.; Park, A.; Viswanathan, M.; Hoet, R.M.; Ladner, R.C. Screening isolates from antibody phage-display libraries. Drug Discov. Today 2008, 13, 318–324. [Google Scholar] [CrossRef]
- Oteng, E.K.; Gu, W.; McKeague, M. High-efficiency enrichment enables identification of aptamers to circulating Plasmodium falciparum-infected erythrocytes. Sci. Rep. 2020, 10, 9706. [Google Scholar] [CrossRef]
- Balaban, M.; Moshiri, N.; Mai, U.; Jia, X.; Mirarab, S. TreeCluster: Clustering biological sequences using phylogenetic trees. PLoS ONE 2019, 14, e0221068. [Google Scholar] [CrossRef] [Green Version]
- Pett, W.; Adamski, M.; Adamska, M.; Francis, W.R.; Eitel, M.; Pisani, D.; Wörheide, G. The role of homology and orthology in the phylogenomic analysis of metazoan gene content. Mol. Biol. Evol. 2019, 36, 643–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, L.S.; Colwell, L.J. Analysis of nanobody paratopes reveals greater diversity than classical antibodies. Protein Eng. Des. Sel. 2018, 31, 267–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knowles, B.B.; Howe, C.C.; Aden, D.P. Human hepatocellular carcinoma cell lines secrete the major plasma proteins and hepatitis B surface antigen. Science 1980, 209, 497–499. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhao, S.; Karnad, A.; Freeman, J.W. The biology and role of CD44 in cancer progression: Therapeutic implications. J. Hematol. Oncol. 2018, 11, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vongchan, P.; Linhardt, R.J. Characterization of a new monoclonal anti-Glypican-3 antibody specific to the hepatocellular carcinoma cell line, HepG2. World J. Hepatol. 2017, 9, 368–384. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Cell Line | 1st Round Absorbent Cell Line | 2nd Round Absorbent Cell Line | 3rd Round Absorbent Cell Line |
|---|---|---|---|
| S1T 2.0 × 106 cell | HepG2, SKBR-3, KYSE520 6.0 × 106 cell (2.0 × 106 cell/cell line) | HepG2, SKBR-3, KYSE520 6.0 × 106 cell (2.0 × 106 cell/cell line) | |
| KYSE520 2.0 × 106 cell | HepG2, SKBR-3, S1T 6.0 × 106 cell (2.0 × 106 cell/cell line) | HepG2, SKBR-3, S1T 6.0 × 106 cell (2.0 × 106 cell/cell line) | HepG2, SKBR-3, S1T 6.0 × 106 cell (2.0 × 106 cell/cell line) |
| SKBR-3 2.0 × 106 cell | HepG2, S1T, KYSE520 6.0 × 106 cell (2.0 × 106 cell/cell line) | HepG2, S1T, KYSE520 6.0 × 106 cell (2.0 × 106 cell/cell line) | |
| HepG2 2.0 × 106 cell | S1T, SKBR-3, KYSE520 6.0 × 106 cell (2.0 × 106 cell/cell line) | S1T, SKBR-3, KYSE520 6.0 × 106 cell (2.0 × 106 cell/cell line) |
| Parameters | ‘0′ Round | 2nd Round S1T Library | 2nd Round HepG2 Library | 2nd Round SKBR-3 Library | 3rd Round KYSE 520 Library |
|---|---|---|---|---|---|
| Total number of read sequences | 398,055 | 471,933 | 354,002 | 499,052 | 261,839 |
| Number of merged sequences | 116,952 (100%) | 186,946 (100%) | 129,050 (100%) | 209,908 (100%) | 94,059 (100%) |
| Single occurrence sequences | 31,497 (26.93%) | 26,685 (14.27%) | 4837 (3.75%) | 9397 (4.48%) | 10,082 (10.72%) |
| Unique sequence | 37,206 (31.81%) | 32,652 (17.47%) | 6958 (5.39%) | 13,733 (6.54%) | 13,358 (14.20%) |
| Highest frequency | 17,611 (15.06%) | 22,471 (12.02%) | 61,377 (47.56%) | 42,054 (20.03%) | 6940 (7.38%) |
| Identical CDR3 Sequences (Group) | S1T Library Clone Number (S1T) | HepG2 Library Clone Number (HepG2) | SKBR3 Library Clone Number (SKBR3) | KYSE 520 Library Clone Number (KYSE520) |
|---|---|---|---|---|
| Group X | 14, 24 & 39 | 5, 15, 17, 25, 27, & 35 | 8, 15, 16, 30, 31, & 36 | |
| Group Y | 2, 9 & 35 | 49 | ||
| Group Z | 4, 7, 22, 29, 40, 41, & 44 | 7, 20, 37, & 49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rafique, A.; Hichiwa, G.; Jatnika, M.F.; Ito, Y. A Novel Strategy for Screening Tumor-Specific Variable Domain of Heavy-Chain Antibodies. Int. J. Mol. Sci. 2023, 24, 10804. https://doi.org/10.3390/ijms241310804
Rafique A, Hichiwa G, Jatnika MF, Ito Y. A Novel Strategy for Screening Tumor-Specific Variable Domain of Heavy-Chain Antibodies. International Journal of Molecular Sciences. 2023; 24(13):10804. https://doi.org/10.3390/ijms241310804
Chicago/Turabian StyleRafique, Abdur, Genki Hichiwa, Muhammad Feisal Jatnika, and Yuji Ito. 2023. "A Novel Strategy for Screening Tumor-Specific Variable Domain of Heavy-Chain Antibodies" International Journal of Molecular Sciences 24, no. 13: 10804. https://doi.org/10.3390/ijms241310804