
Multiple-Factors-Induced Rheumatoid Arthritis Synoviocyte Activation Is Attenuated by the α2-Adrenergic Receptor Agonist Dexmedetomidine

Abstract
:1. Introduction
2. Results
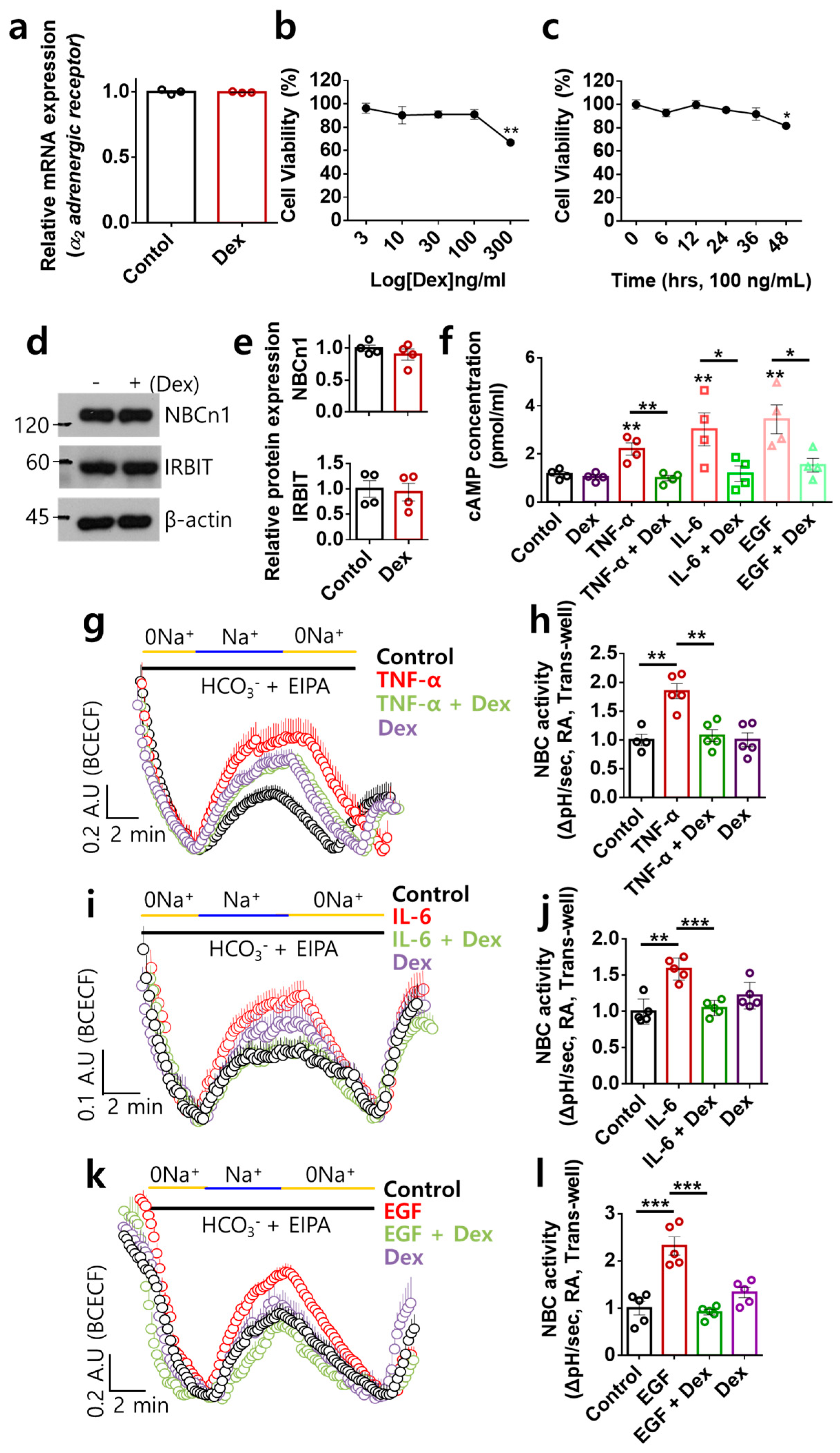
2.1. Dex Inhibited NBC Activities Stimulated by Multiple Inflammatory or Proliferative Inputs in RA-FLSs
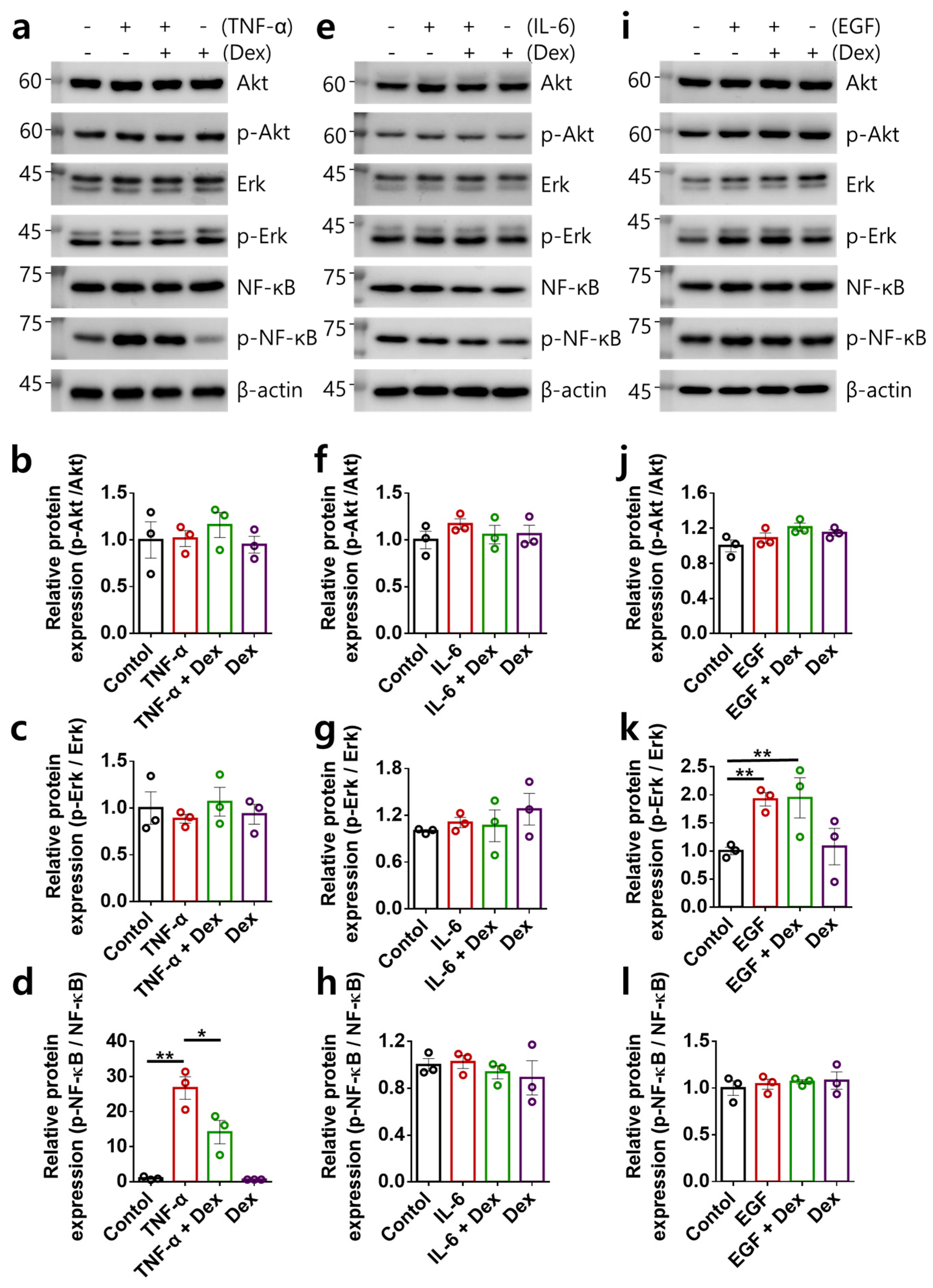
2.2. Multiple-Input-Mediated RA-FLS Activation Is Not Associated with Akt, Erk, and NF-κB Signaling
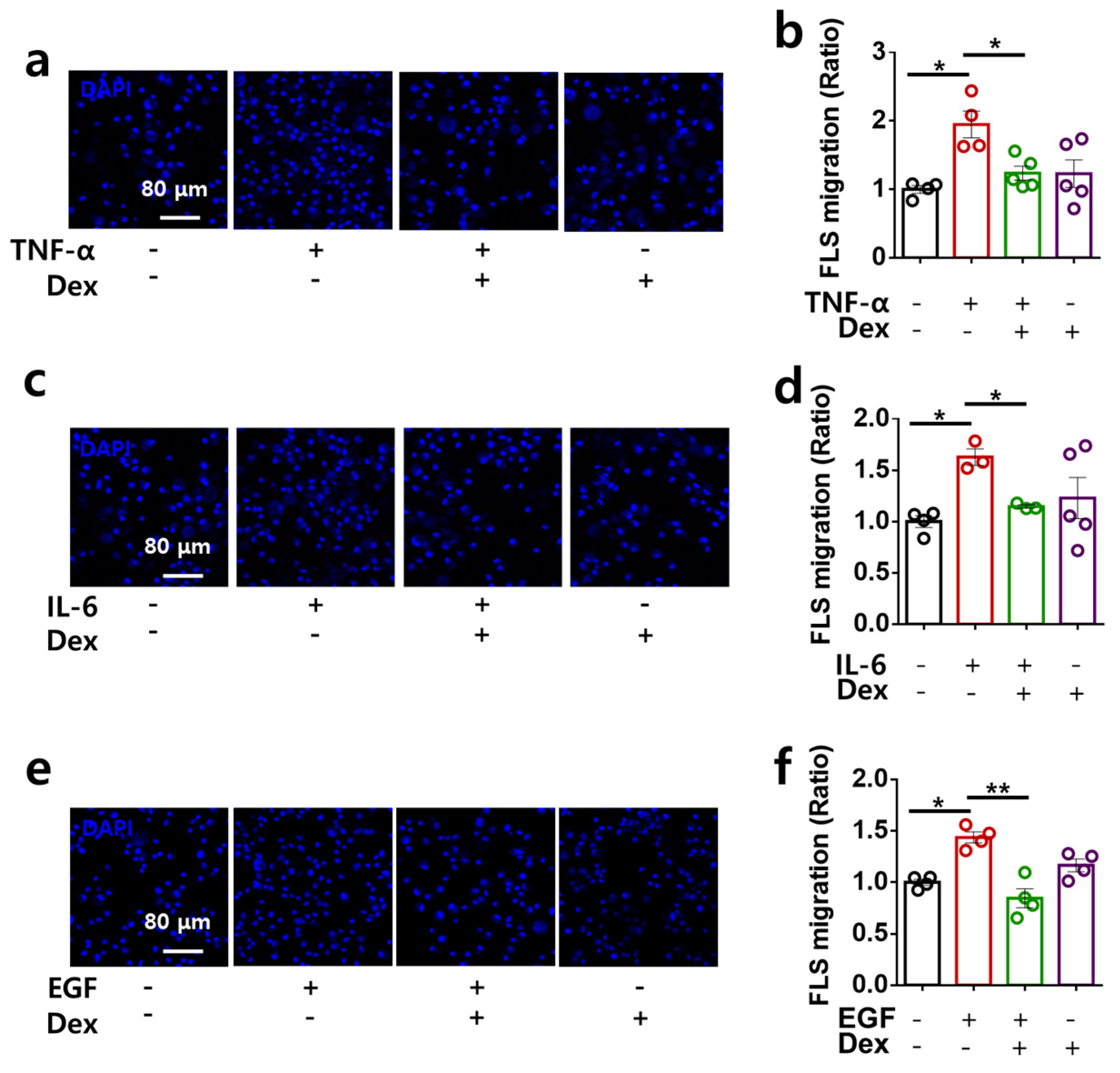
2.3. Dex Attenuates RA-FLS Migration Stimulated by Multiple Inputs
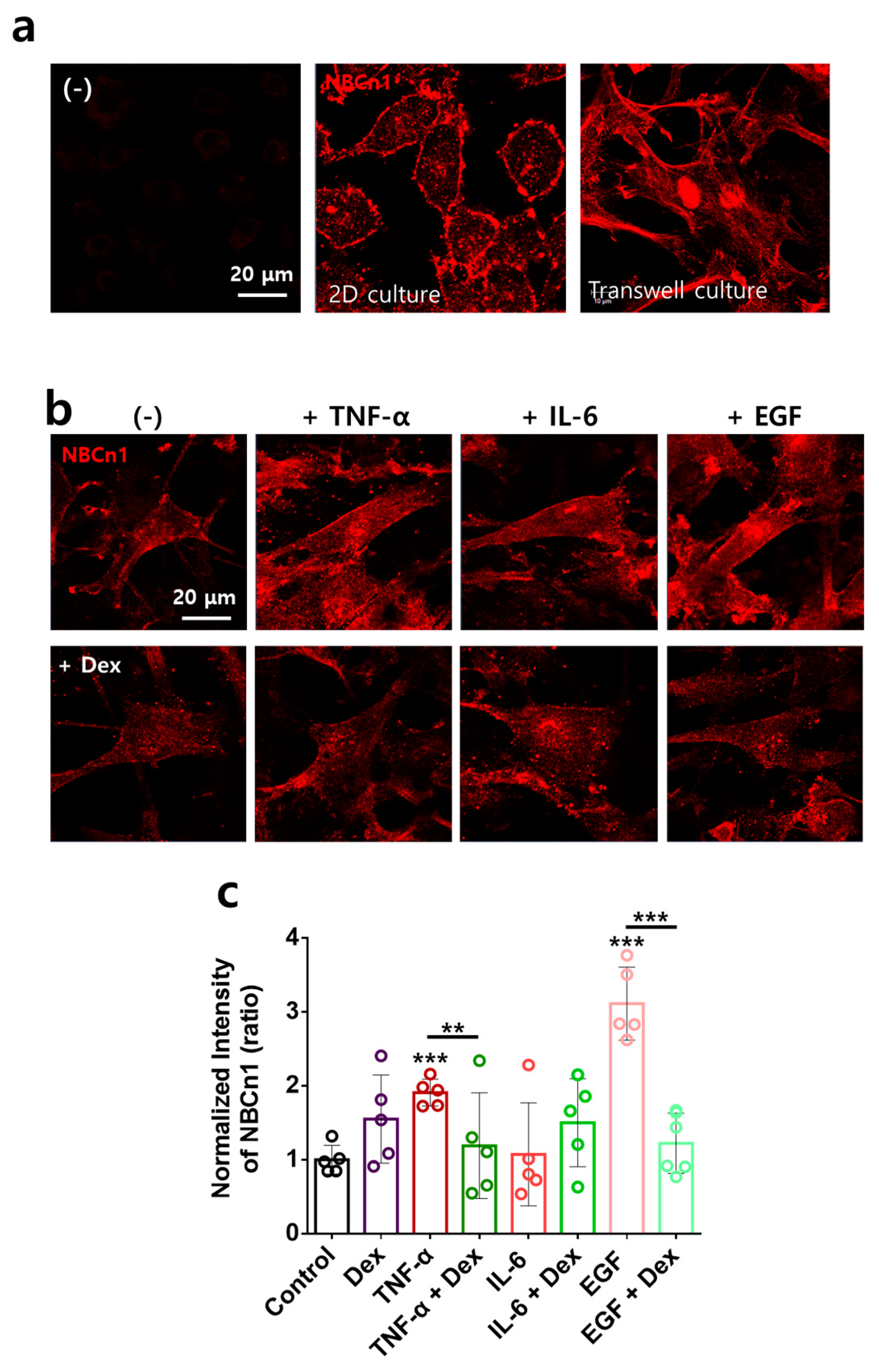
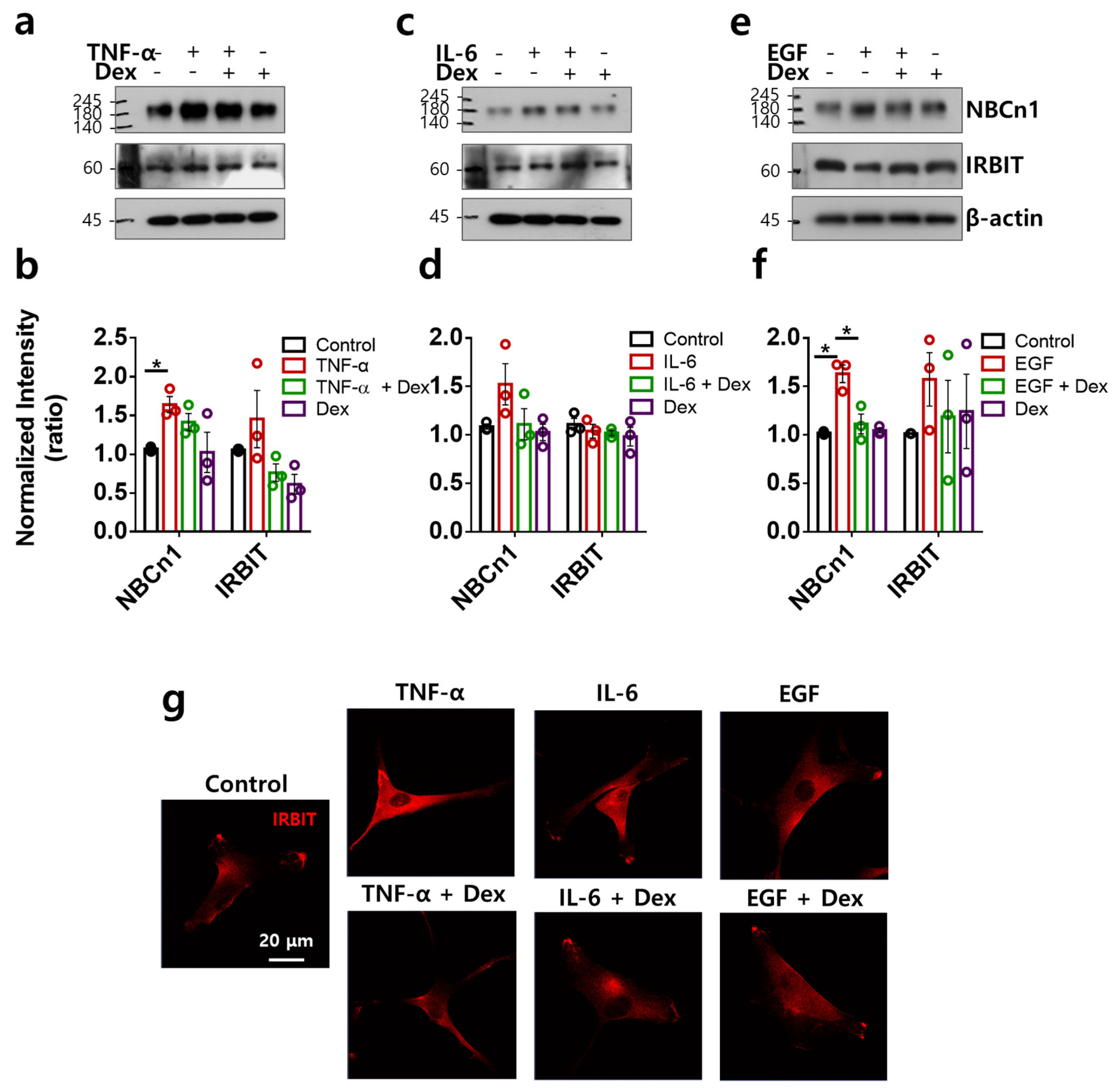
2.4. Directional Stimulation of TNF-α- and EGF-Mediated Membranous NBCn1 Expression in RAFLS Is Attenuated by Dex
2.5. TNF-α- and EGF-Mediated NBCn1 Expression Is Attenuated by Dex with No Change in IRBIT in RA-FLSs
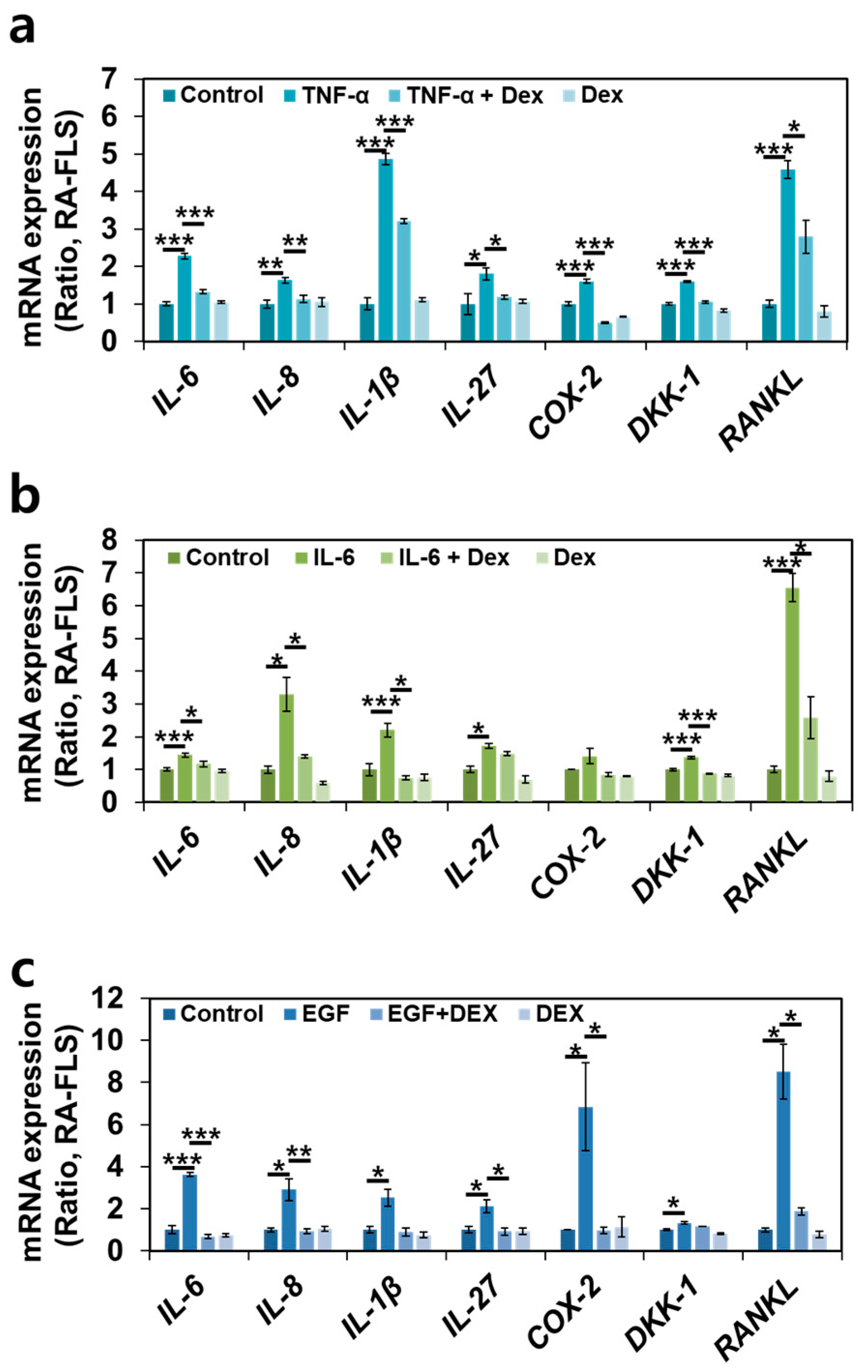
2.6. Dex Treatment Inhibits Multiple RA-Associated Factors in RA-FLSs
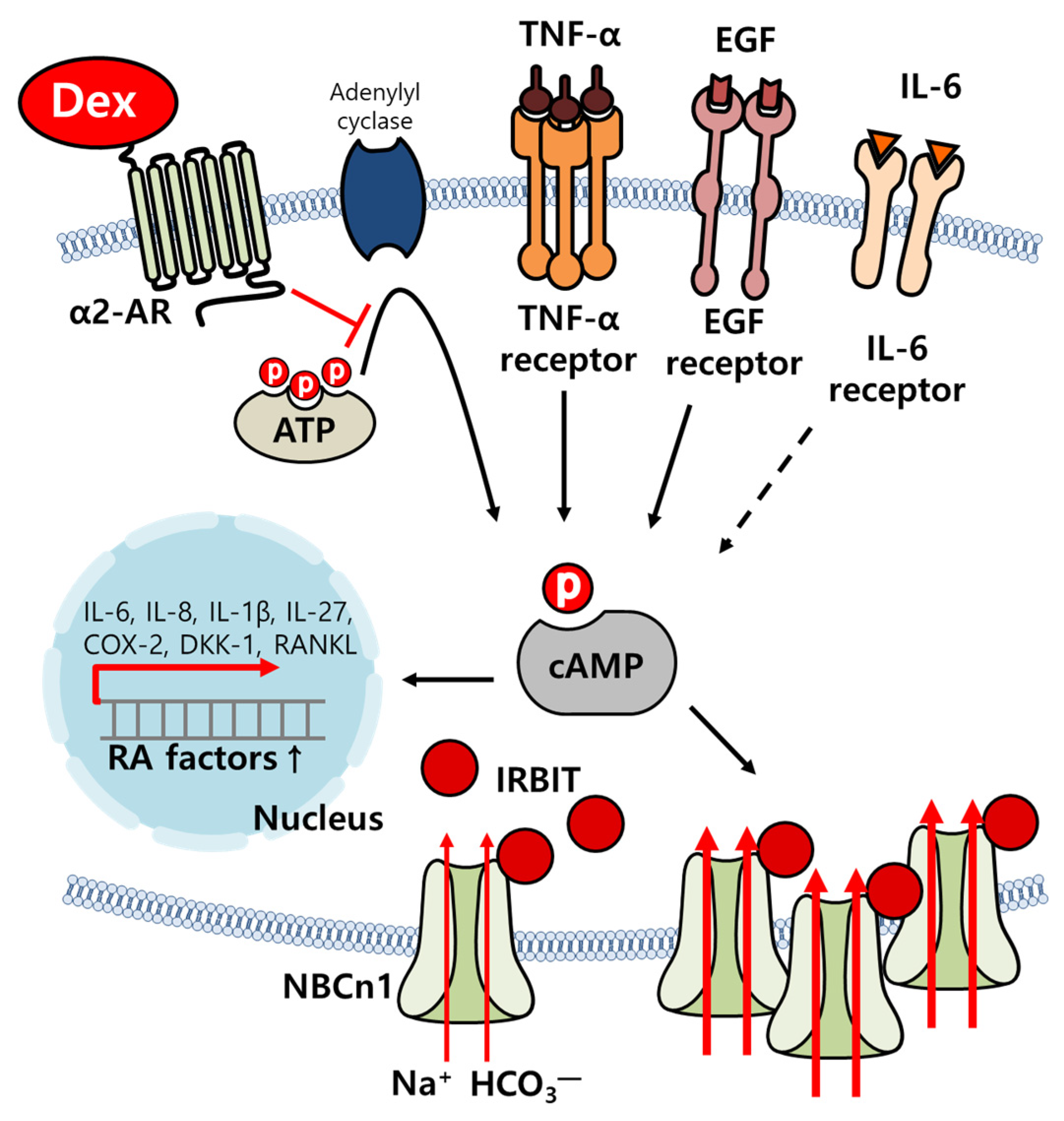
3. Discussion
4. Materials and Methods
4.1. RA-FLS Culture
4.2. Total RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
4.3. MTT Assay
4.4. Western Blot
4.5. Measurement of Cyclic Adenosine Monophosphate (cAMP)
4.6. Measurement of the Na+-HCO3− Cotransporter (NBC) Activity
4.7. Transwell Membrane-Based Migration Assay
4.8. Immunostaining of the Inserted Membrane in the Transwell Culture System
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ji, M.; Ryu, H.J.; Hong, J.H. Signalling and putative therapeutic molecules on the regulation of synoviocyte signalling in rheumatoid arthritis. Bone Jt. Res. 2021, 10, 285–297. [Google Scholar] [CrossRef]
- Palmer, M.; Stanford, E.; Murray, M.M. The Effect of Synovial Fluid Enzymes on the Biodegradability of Collagen and Fibrin Clots. Materials 2011, 4, 1469–1482. [Google Scholar] [CrossRef] [PubMed]
- Schendrigin, I.N.; Timchenko, L.D.; Rzhepakovsky, I.V.; Avanesyan, S.S.; Sizonenko, M.N.; Grimm, W.D.; Povetkin, S.N.; Piskov, S.I. Clinical and Pathogenetic Significance of Amylase Level and Microtomographic Index of Synovial Fluid in Various Joint Lesions. Sovrem. Tehnol. Med. 2022, 14, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Kitano, T.; Eguchi, R.; Okamatsu-Ogura, Y.; Yamaguchi, S.; Otsuguro, K.I. Opposing functions of alpha- and beta-adrenoceptors in the formation of processes by cultured astrocytes. J. Pharmacol. Sci. 2021, 145, 228–240. [Google Scholar] [CrossRef]
- Ji, M.; Park, C.K.; Lee, J.W.; Park, K.Y.; Son, K.H.; Hong, J.H. Two Phase Modulation of NH4+ Entry and Cl−/HCO3− Exchanger in Submandibular Glands Cells by Dexmedetomidine. Front. Physiol. 2017, 8, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, J.; Sun, P.; Zhao, H.; Watts, H.R.; Sanders, R.D.; Terrando, N.; Xia, P.; Maze, M.; Ma, D. Dexmedetomidine provides renoprotection against ischemia-reperfusion injury in mice. Crit. Care 2011, 15, R153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, H.; Chi, X.; Jin, Y.; Wang, Y.; Huang, P.; Wu, S.; Xia, Z.; Cai, J. Dexmedetomidine Inhibits TLR4/NF-kappaB Activation and Reduces Acute Kidney Injury after Orthotopic Autologous Liver Transplantation in Rats. Sci. Rep. 2015, 5, 16849. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Du, X.Y.; Wang, Q.Y.; Xiao, S.L.; Zhi, J.; Gao, H.B.; Yang, D. Dexmedetomidine Protects against Airway Inflammation and Airway Remodeling in a Murine Model of Chronic Asthma through TLR4/NF-kappa B Signaling Pathway. Mediat. Inflamm. 2023, 2023, 3695469. [Google Scholar] [CrossRef]
- de Jonge, W.J.; van der Zanden, E.P.; The, F.O.; Bijlsma, M.F.; van Westerloo, D.J.; Bennink, R.J.; Berthoud, H.R.; Uematsu, S.; Akira, S.; van den Wijngaard, R.M.; et al. Stimulation of the vagus nerve attenuates macrophage activation by activating the Jak2-STAT3 signaling pathway. Nat. Immunol. 2005, 6, 844–851. [Google Scholar] [CrossRef]
- Chatterjee, P.K.; Al-Abed, Y.; Sherry, B.; Metz, C.N. Cholinergic agonists regulate JAK2/STAT3 signaling to suppress endothelial cell activation. Am. J. Physiol. Cell Physiol. 2009, 297, C1294–C1306. [Google Scholar] [CrossRef] [Green Version]
- Bao, N.; Tang, B. Organ-Protective Effects and the Underlying Mechanism of Dexmedetomidine. Mediat. Inflamm. 2020, 2020, 6136105. [Google Scholar] [CrossRef]
- Chen, R.; Dou, X.K.; Dai, M.S.; Sun, Y.; Sun, S.J.; Wu, Y. The role of dexmedetomidine in immune tissue and inflammatory diseases: A narrative review. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 8030–8038. [Google Scholar] [CrossRef]
- Ji, Y.R.; Chen, Y.; Chen, Y.N.; Qiu, G.L.; Wen, J.G.; Zheng, Y.; Li, X.F.; Cheng, H.; Li, Y.H.; Li, J. Dexmedetomidine inhibits the invasion, migration, and inflammation of rheumatoid arthritis fibroblast-like synoviocytes by reducing the expression of NLRC5. Int. Immunopharmacol. 2020, 82, 106374. [Google Scholar] [CrossRef]
- Lachaine, J.; Beauchemin, C. Economic evaluation of dexmedetomidine relative to midazolam for sedation in the intensive care unit. Can. J. Hosp. Pharm. 2012, 65, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Kaur, M.; Singh, P.M. Current role of dexmedetomidine in clinical anesthesia and intensive care. Anesth. Essays Res. 2011, 5, 128–133. [Google Scholar] [CrossRef] [Green Version]
- Bernardo, A.A.; Bernardo, C.M.; Espiritu, D.J.; Arruda, J.A. The sodium bicarbonate cotransporter: Structure, function, and regulation. Semin. Nephrol. 2006, 26, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zahra, A.; Wang, Y.; Wu, J. Understanding the Physiological Role of Electroneutral Na(+)-Coupled HCO(3)(-) Cotransporter and Its Therapeutic Implications. Pharmaceuticals 2022, 15, 1082. [Google Scholar] [CrossRef]
- Ji, M.; Ryu, H.J.; Baek, H.M.; Shin, D.M.; Hong, J.H. Dynamic synovial fibroblasts are modulated by NBCn1 as a potential target in rheumatoid arthritis. Exp. Mol. Med. 2022, 54, 503–517. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, O.; Rossmann, H.; Berger, U.V.; Colledge, W.H.; Ratcliff, R.; Evans, M.J.; Gregor, M.; Seidler, U. cAMP-mediated regulation of murine intestinal/pancreatic Na+/HCO3- cotransporter subtype pNBC1. Am. J. Physiol. Gastrointest Liver Physiol. 2003, 284, G37–G45. [Google Scholar] [CrossRef] [Green Version]
- Espejo, M.S.; Orlowski, A.; Ibanez, A.M.; Di Mattia, R.A.; Velasquez, F.C.; Rossetti, N.S.; Ciancio, M.C.; De Giusti, V.C.; Aiello, E.A. The functional association between the sodium/bicarbonate cotransporter (NBC) and the soluble adenylyl cyclase (sAC) modulates cardiac contractility. Pflug. Arch. Eur. J. Physiol. 2020, 472, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Shcheynikov, N.; Son, A.; Hong, J.H.; Yamazaki, O.; Ohana, E.; Kurtz, I.; Shin, D.M.; Muallem, S. Intracellular Cl- as a signaling ion that potently regulates Na+/HCO3- transporters. Proc. Natl. Acad. Sci. USA 2015, 112, E329–E337. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.; Shin, D.M.; Hong, J.H. Protective Role of IRBIT on Sodium Bicarbonate Cotransporter-n1 for Migratory Cancer Cells. Pharmaceutics 2020, 12, 816. [Google Scholar] [CrossRef] [PubMed]
- Schwab, A.; Fabian, A.; Hanley, P.J.; Stock, C. Role of ion channels and transporters in cell migration. Physiol. Rev. 2012, 92, 1865–1913. [Google Scholar] [CrossRef]
- Morishita, K.; Watanabe, K.; Ichijo, H. Cell volume regulation in cancer cell migration driven by osmotic water flow. Cancer Sci. 2019, 110, 2337–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartok, B.; Firestein, G.S. Fibroblast-like synoviocytes: Key effector cells in rheumatoid arthritis. Immunol. Rev. 2010, 233, 233–255. [Google Scholar] [CrossRef] [PubMed]
- Bo, J.H.; Wang, J.X.; Wang, X.L.; Jiao, Y.; Jiang, M.; Chen, J.L.; Hao, W.Y.; Chen, Q.; Li, Y.H.; Ma, Z.L.; et al. Dexmedetomidine Attenuates Lipopolysaccharide-Induced Sympathetic Activation and Sepsis via Suppressing Superoxide Signaling in Paraventricular Nucleus. Antioxidants 2022, 11, 2395. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ji, F.; Liang, J.; He, H.; Fu, Y.; Cao, M. Inhibition by dexmedetomidine of the activation of spinal dorsal horn glias and the intracellular ERK signaling pathway induced by nerve injury. Brain Res. 2012, 1427, 1–9. [Google Scholar] [CrossRef]
- Wei, B.; Liu, W.; Jin, L.; Guo, S.; Fan, H.; Jin, F.; Wei, C.; Fang, D.; Zhang, X.; Su, S.; et al. Dexmedetomidine Inhibits Gasdermin D-Induced Pyroptosis via the PI3K/AKT/GSK3beta Pathway to Attenuate Neuroinflammation in Early Brain Injury After Subarachnoid Hemorrhage in Rats. Front. Cell. Neurosci. 2022, 16, 899484. [Google Scholar] [CrossRef]
- Ye, X.; Xu, H.; Xu, Y. Dexmedetomidine alleviates intestinal barrier dysfunction and inflammatory response in mice via suppressing TLR4/MyD88/NF-kappaB signaling in an experimental model of ulcerative colitis. Folia Histochem. Cytobiol. 2022, 60, 311–322. [Google Scholar] [CrossRef]
- Ando, H.; Kawaai, K.; Mikoshiba, K. IRBIT: A regulator of ion channels and ion transporters. Biochim. Biophys. Acta 2014, 1843, 2195–2204. [Google Scholar] [CrossRef] [Green Version]
- Ando, H.; Kawaai, K.; Bonneau, B.; Mikoshiba, K. Remodeling of Ca(2+) signaling in cancer: Regulation of inositol 1,4,5-trisphosphate receptors through oncogenes and tumor suppressors. Adv. Biol. Regul. 2018, 68, 64–76. [Google Scholar] [CrossRef]
- Han, L.; Chen, Z.; Yu, K.; Yan, J.; Li, T.; Ba, X.; Lin, W.; Huang, Y.; Shen, P.; Huang, Y.; et al. Interleukin 27 Signaling in Rheumatoid Arthritis Patients: Good or Evil? Front. Immunol. 2021, 12, 787252. [Google Scholar] [CrossRef]
- Ruaro, B.; Casabella, A.; Paolino, S.; Pizzorni, C.; Ghio, M.; Seriolo, C.; Molfetta, L.; Odetti, P.; Smith, V.; Cutolo, M. Dickkopf-1 (Dkk-1) serum levels in systemic sclerosis and rheumatoid arthritis patients: Correlation with the Trabecular Bone Score (TBS). Clin. Rheumatol. 2018, 37, 3057–3062. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.Y.; Kang, N.; Yang, Y.M.; Hong, J.H.; Shin, D.M. The Role of Ca(2+)-NFATc1 Signaling and Its Modulation on Osteoclastogenesis. Int. J. Mol. Sci. 2020, 21, 3646. [Google Scholar] [CrossRef] [PubMed]
- Okada, H.; Kurita, T.; Mochizuki, T.; Morita, K.; Sato, S. The cardioprotective effect of dexmedetomidine on global ischaemia in isolated rat hearts. Resuscitation 2007, 74, 538–545. [Google Scholar] [CrossRef] [Green Version]
- Si, Y.N.; Bao, H.G.; Xu, L.; Wang, X.L.; Shen, Y.; Wang, J.S.; Yang, X.B. Dexmedetomidine protects against ischemia/reperfusion injury in rat kidney. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 1843–1851. [Google Scholar]
- Weng, J.; Cheng, Z.; Li, S. Effects of dexmedetomidine on surgical site wound pain in patients undergoing laparoscopic cholecystectomy: A meta-analysis. Int. Wound J. 2023. [Google Scholar] [CrossRef] [PubMed]
- Abuelazm, M.T.; Ghanem, A.; Johanis, A.; Mahmoud, A.; Hassan, A.R.; Katamesh, B.E.; Amin, M.A.; Abdelazeem, B. Reno-protective effects of perioperative dexmedetomidine in kidney transplantation: A systematic review and meta-analysis of randomized controlled trials. Int. Urol. Nephrol. 2023, 1–12. [Google Scholar] [CrossRef]
- Boron, W.F. Acid-base transport by the renal proximal tubule. J. Am. Soc. Nephrol. 2006, 17, 2368–2382. [Google Scholar] [CrossRef] [Green Version]
- Severin, M.; Pedersen, E.L.; Borre, M.T.; Axholm, I.; Christiansen, F.B.; Ponniah, M.; Czaplinska, D.; Larsen, T.; Pardo, L.A.; Pedersen, S.F. Dynamic localization of the Na+-HCO3- co-transporter NBCn1 to the plasma membrane, centrosomes, spindle and primary cilia. J. Cell Sci. 2023, 136, jcs260687. [Google Scholar] [CrossRef]
- Romero, M.F.; Chen, A.P.; Parker, M.D.; Boron, W.F. The SLC4 family of bicarbonate (HCO(3)(-)) transporters. Mol. Asp. Med. 2013, 34, 159–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khandoudi, N.; Albadine, J.; Robert, P.; Krief, S.; Berrebi-Bertrand, I.; Martin, X.; Bevensee, M.O.; Boron, W.F.; Bril, A. Inhibition of the cardiac electrogenic sodium bicarbonate cotransporter reduces ischemic injury. Cardiovasc. Res. 2001, 52, 387–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remigante, A.; Spinelli, S.; Pusch, M.; Sarikas, A.; Morabito, R.; Marino, A.; Dossena, S. Role of SLC4 and SLC26 solute carriers during oxidative stress. Acta Physiol. 2022, 235, e13796. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Lee, P.C.W.; Hong, J.H. Lamin-A/C Is Modulated by the Involvement of Histamine-Mediated Calcium/Calmodulin-Dependent Kinase II in Lung Cancer Cells. Int. J. Mol. Sci. 2022, 23, 9075. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.; Kim, H.J.; Ahn, C.B.; Son, K.H.; Hong, J.H. Cellular channelopathy mediated by hypergravity: IL-6-mediated Nkcc1 activation and enhanced Trpm2 expression in rat atrium. Cell Tissue Res. 2021, 383, 1017–1024. [Google Scholar] [CrossRef]
- Hong, J.H.; Yang, D.; Shcheynikov, N.; Ohana, E.; Shin, D.M.; Muallem, S. Convergence of IRBIT, phosphatidylinositol (4,5) bisphosphate, and WNK/SPAK kinases in regulation of the Na+-HCO3- cotransporters family. Proc. Natl. Acad. Sci. USA 2013, 110, 4105–4110. [Google Scholar] [CrossRef] [Green Version]
- Ji, M.J.; Hong, J.H. An overview of carbonic anhydrases and membrane channels of synoviocytes in inflamed joints. J. Enzym. Inhib. Med. Chem. 2019, 34, 1615–1622. [Google Scholar] [CrossRef] [Green Version]
- Kondo, N.; Kuroda, T.; Kobayashi, D. Cytokine Networks in the Pathogenesis of Rheumatoid Arthritis. Int. J. Mol. Sci. 2021, 10922. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Hirota, K.; Sakaguchi, S. Synovial Tissue Inflammation Mediated by Autoimmune T Cells. Front. Immunol. 2019, 10, 1989. [Google Scholar] [CrossRef]
- Paunovic, V.; Carroll, H.P.; Vandenbroeck, K.; Gadina, M. Signalling, inflammation and arthritis: Crossed signals: The role of interleukin (IL)-12, -17, -23 and -27 in autoimmunity. Rheumatology 2008, 47, 771–776. [Google Scholar] [CrossRef] [Green Version]
- Tu, J.; Wang, X.; Gong, X.; Hong, W.; Han, D.; Fang, Y.; Guo, Y.; Wei, W. Synovial Macrophages in Rheumatoid Arthritis: The Past, Present, and Future. Mediat. Inflamm. 2020, 2020, 1583647. [Google Scholar] [CrossRef] [PubMed]
- Smolen, J.S.; Aletaha, D.; McInnes, I.B. Rheumatoid arthritis. Lancet 2016, 388, 2023–2038. [Google Scholar] [CrossRef]
- Smolen, J.S.; Aletaha, D.; Koeller, M.; Weisman, M.H.; Emery, P. New therapies for treatment of rheumatoid arthritis. Lancet 2007, 370, 1861–1874. [Google Scholar] [CrossRef]
- Aga, A.B.; Lie, E.; Uhlig, T.; Olsen, I.C.; Wierod, A.; Kalstad, S.; Rodevand, E.; Mikkelsen, K.; Kvien, T.K.; Haavardsholm, E.A. Time trends in disease activity, response and remission rates in rheumatoid arthritis during the past decade: Results from the NOR-DMARD study 2000–2010. Ann. Rheum. Dis. 2015, 74, 381–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negi, S.; Tandel, N.; Sharma, P.; Kumar, R.; Tyagi, R.K. Aceclofenac and methotrexate combination therapy could influence Th1/Th17 axis to modulate rheumatoid-arthritis-induced inflammation. Drug Discov. Today 2023, 20, 103671. [Google Scholar] [CrossRef] [PubMed]
- Rzhepakovsky, I.; Anusha Siddiqui, S.; Avanesyan, S.; Benlidayi, M.; Dhingra, K.; Dolgalev, A.; Enukashvily, N.; Fritsch, T.; Heinz, V.; Kochergin, S.; et al. Anti-arthritic effect of chicken embryo tissue hydrolyzate against adjuvant arthritis in rats (X-ray microtomographic and histopathological analysis). Food Sci. Nutr. 2021, 9, 5648–5669. [Google Scholar] [CrossRef] [PubMed]
- Areshidze, D.A.; Timchenko, L.D.; Rzhepakovsky, I.V.; Kozlova, M.A.; Kusnetsova, I.A.; Makartseva, L.A. Anti-Inflammatory Effect of Nicavet-2500 in Rodent Models of Acute Inflammation. J. Pharm. Nutr. Sci. 2018, 8, 35–41. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequences (5′→3′) |
|---|---|
| Human GAPDH | (Forward) GAC CTG ACC TGC CGT CTA GAA A (Reverse) CCT GCT TCA CCA CCT TCT TGA |
| Human α2-AR | (Forward) CCC GGT CAT CTA CAC CAT CTT (Reverse) CCC GAC AGA GGA TCT TCT TGA |
| Human IL-6 | (Forward) CCC CCA GGA GAA GAT TCC AA (Reverse) CCG TCG AGG ATG TAC CGA ATT |
| Human IL-8 | (Forward) GTG GCT GAA CCA GAG TTG GAA (Reverse) TGG TGC ACT GGA GCT GCT T |
| Human IL-1β | (Forward) CCA CGG CCA CAT TTG GTT (Reverse) AGG GAA GCG GTT GCT CAT C |
| Human IL-27 | (Forward) GCT TGG ATG TCC CGA AAC C (Reverse) CGC CAC CCC TTG CTA AAA T |
| Human COX-2 | (Forward) GCT CAG CCA TAC AGC AAA TCC (Reverse) GTC CGG GTA CAA TCG CAC TT |
| Human DKK-1 | (Forward) TGG AAC TCC CCT GTG ATT GC (Reverse) TGG AAC TCC CCT GTG ATT GC |
| Human RANKL | (Forward) GGG TCT TTG TCG CGA TGG TA (Reverse) CTG GTA CTT ATT CCC GCC CG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.; Hong, J.H. Multiple-Factors-Induced Rheumatoid Arthritis Synoviocyte Activation Is Attenuated by the α2-Adrenergic Receptor Agonist Dexmedetomidine. Int. J. Mol. Sci. 2023, 24, 10756. https://doi.org/10.3390/ijms241310756
Lee D, Hong JH. Multiple-Factors-Induced Rheumatoid Arthritis Synoviocyte Activation Is Attenuated by the α2-Adrenergic Receptor Agonist Dexmedetomidine. International Journal of Molecular Sciences. 2023; 24(13):10756. https://doi.org/10.3390/ijms241310756
Chicago/Turabian StyleLee, Dongun, and Jeong Hee Hong. 2023. "Multiple-Factors-Induced Rheumatoid Arthritis Synoviocyte Activation Is Attenuated by the α2-Adrenergic Receptor Agonist Dexmedetomidine" International Journal of Molecular Sciences 24, no. 13: 10756. https://doi.org/10.3390/ijms241310756