
Genome-Wide Identification and Expression Analysis of AS2 Genes in Brassica rapa Reveal Their Potential Roles in Abiotic Stress

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification and Physicochemical Characterization of the Brassica rapa AS2 Genes
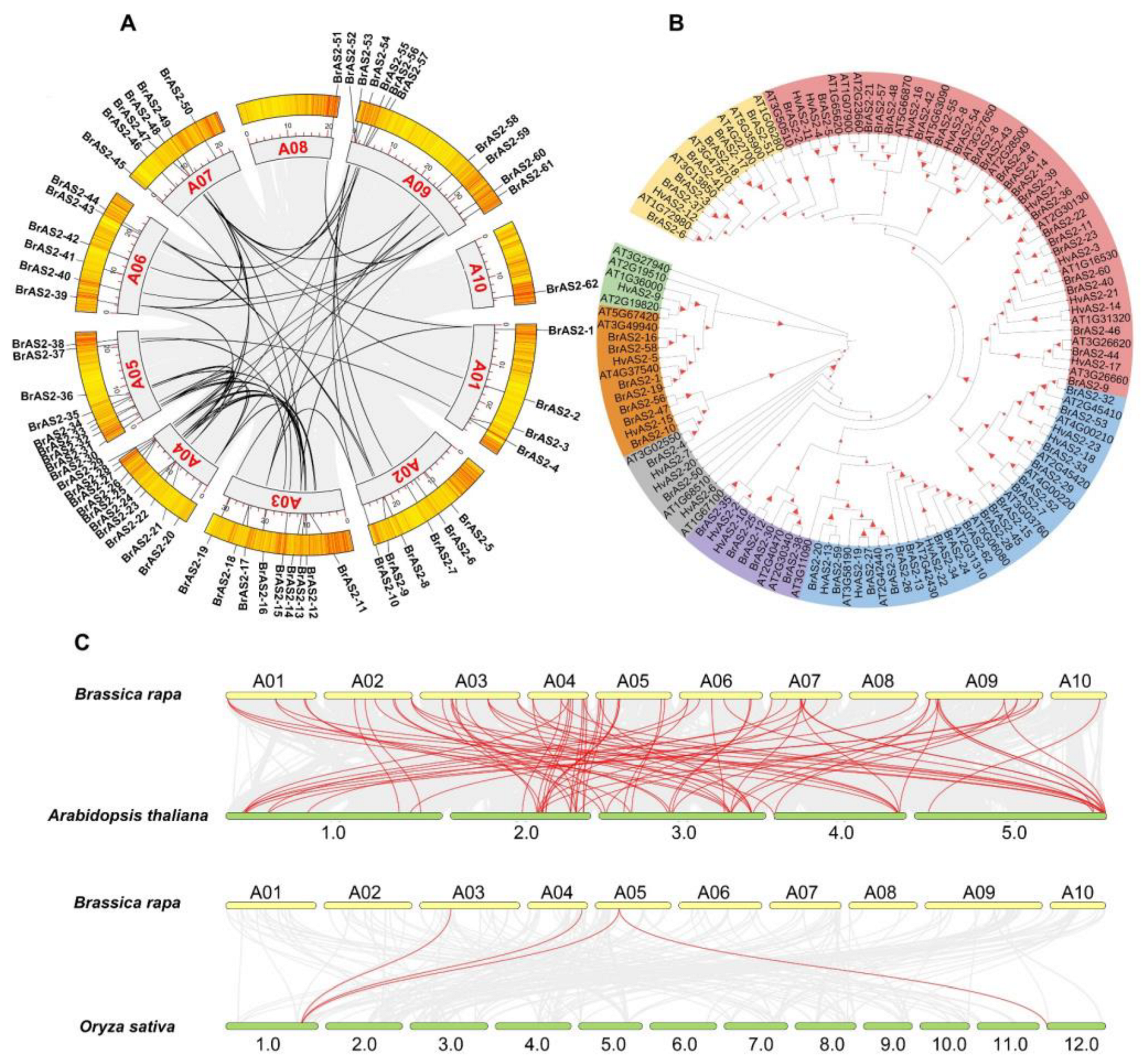
2.2. Phylogenetic Relationships and Synteny Analysis
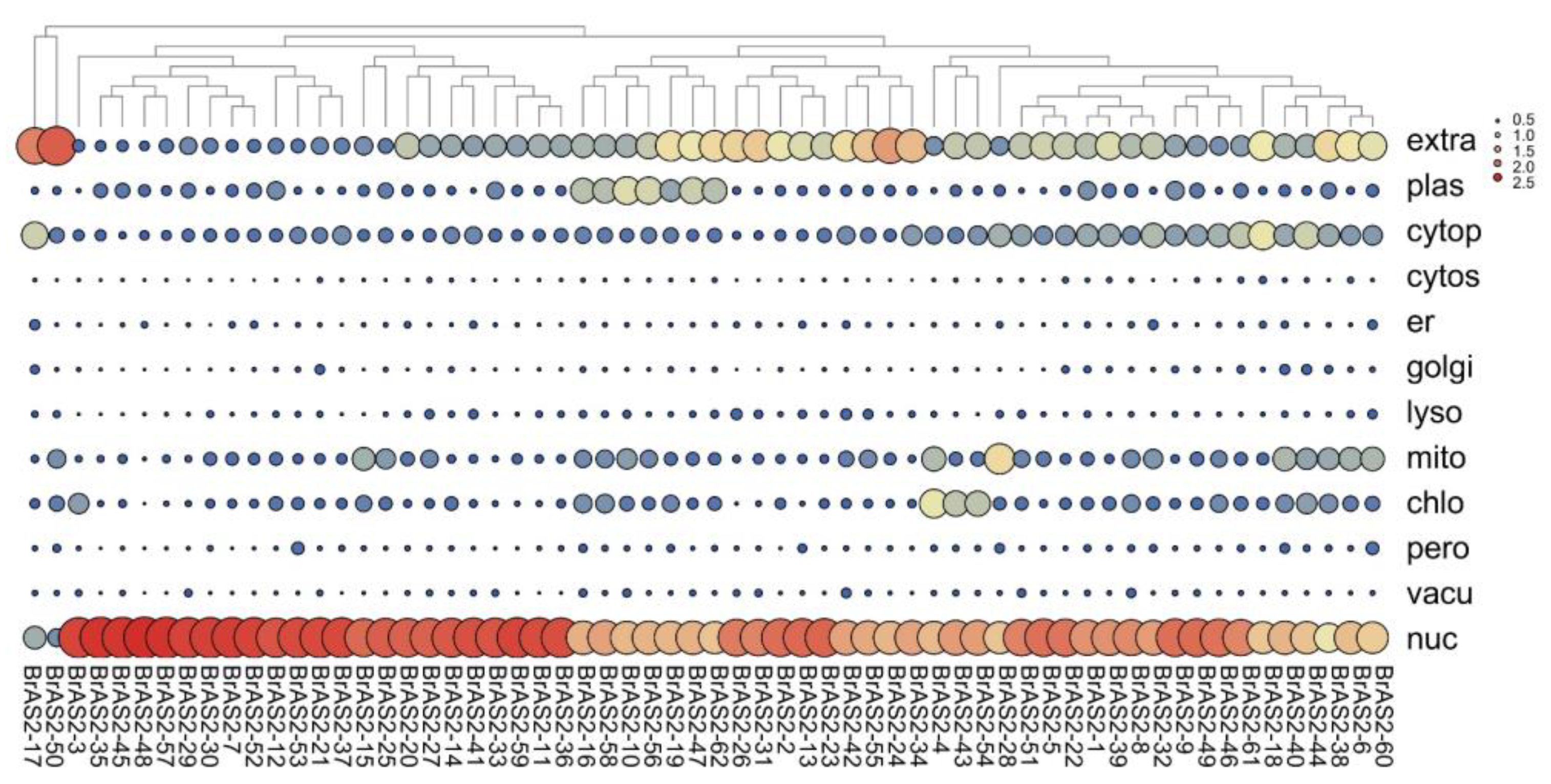
2.3. Subcellular Localization Analysis
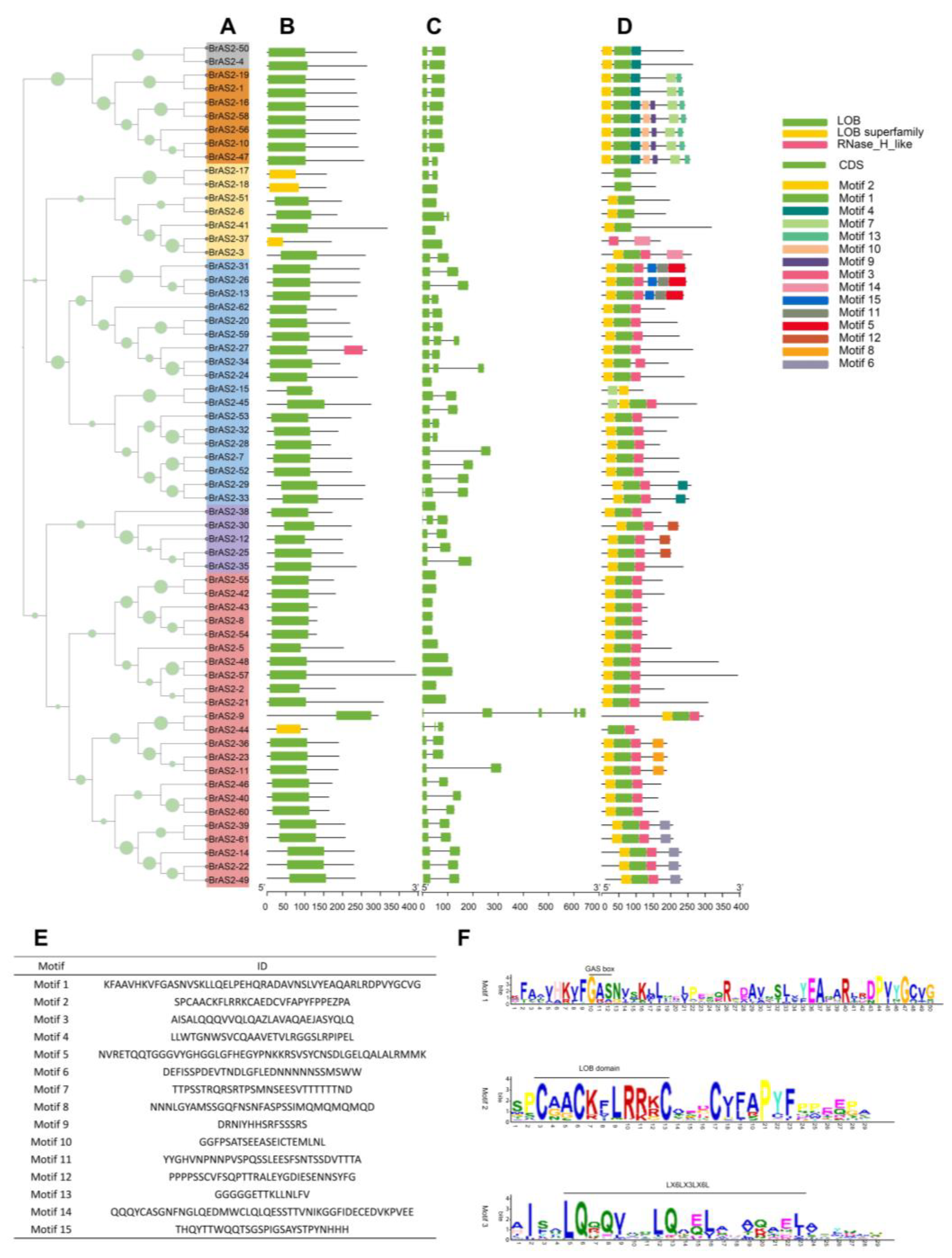
2.4. Gene Structure and Conserved Motif Analysis
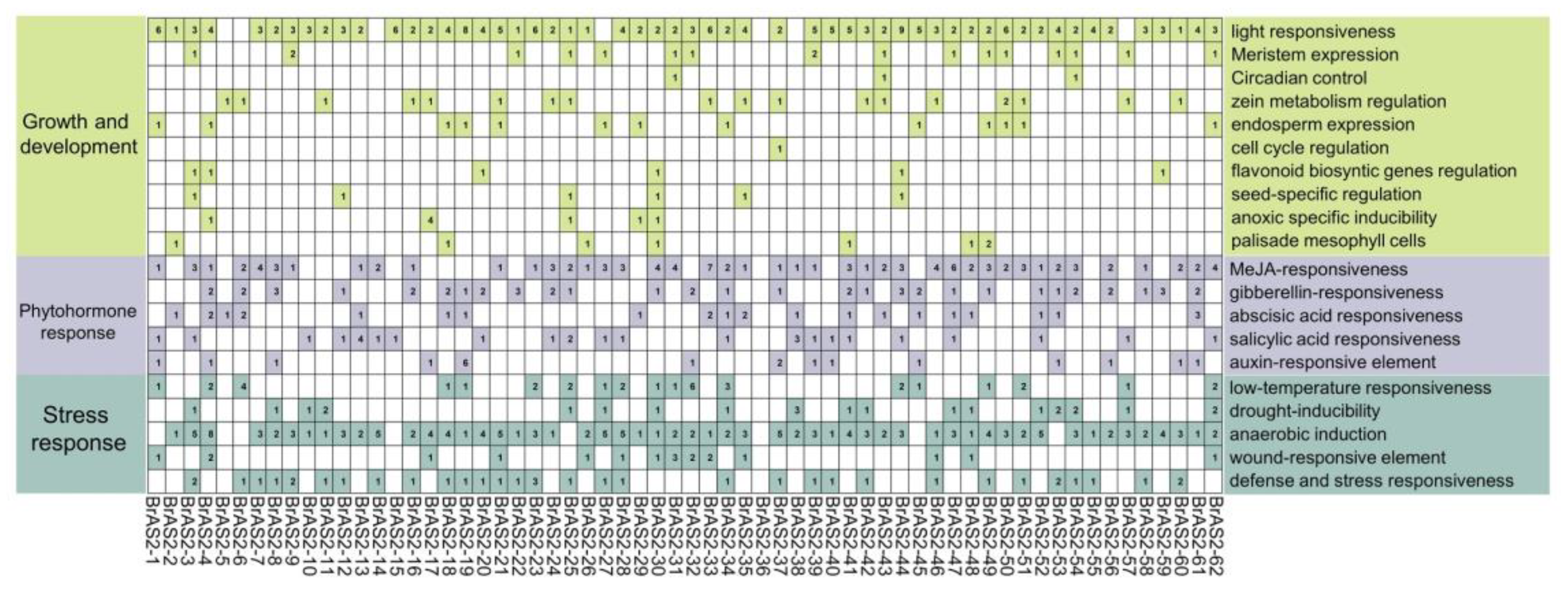
2.5. cis-Element Analysis
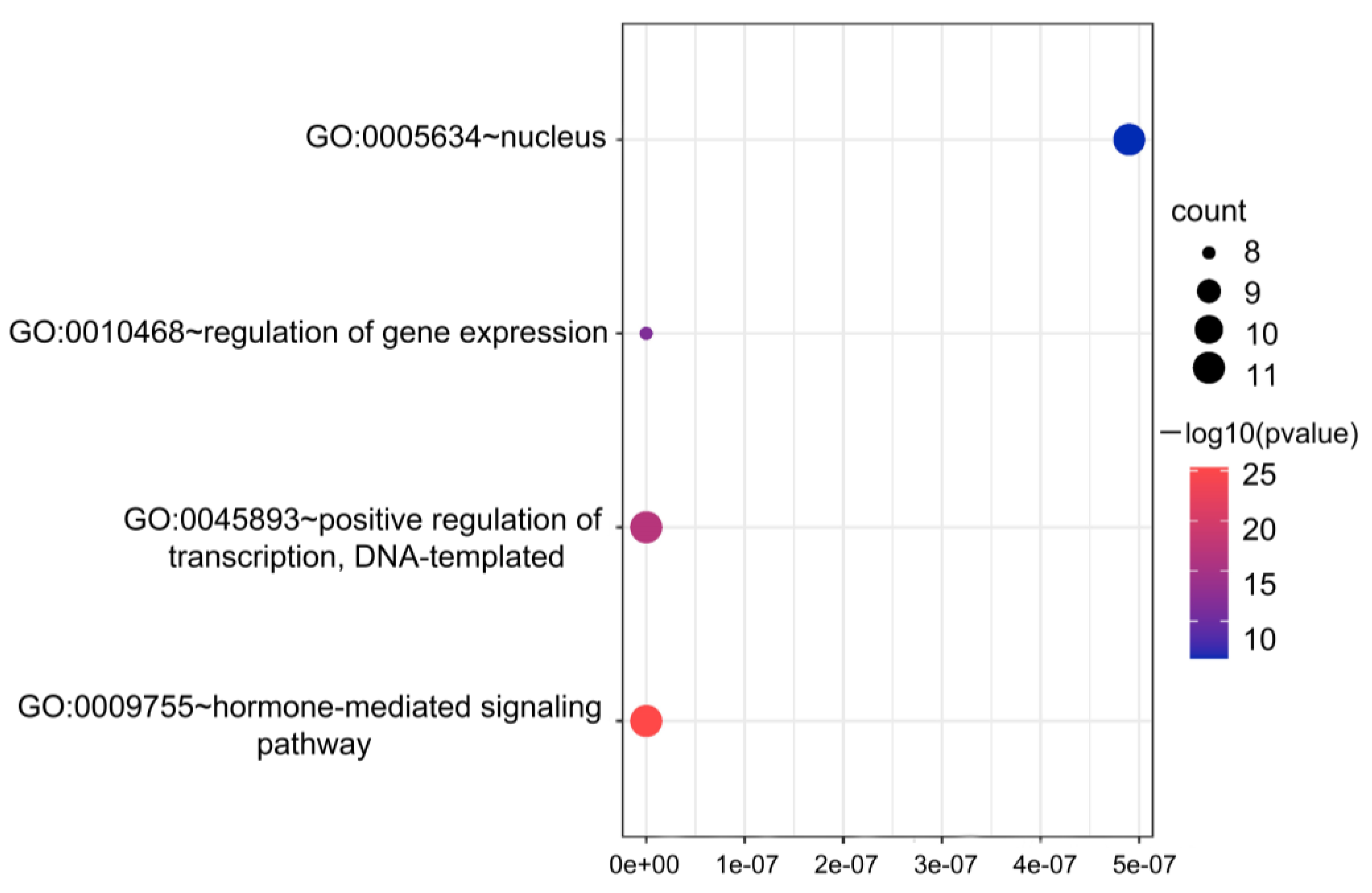
2.6. GO Analysis
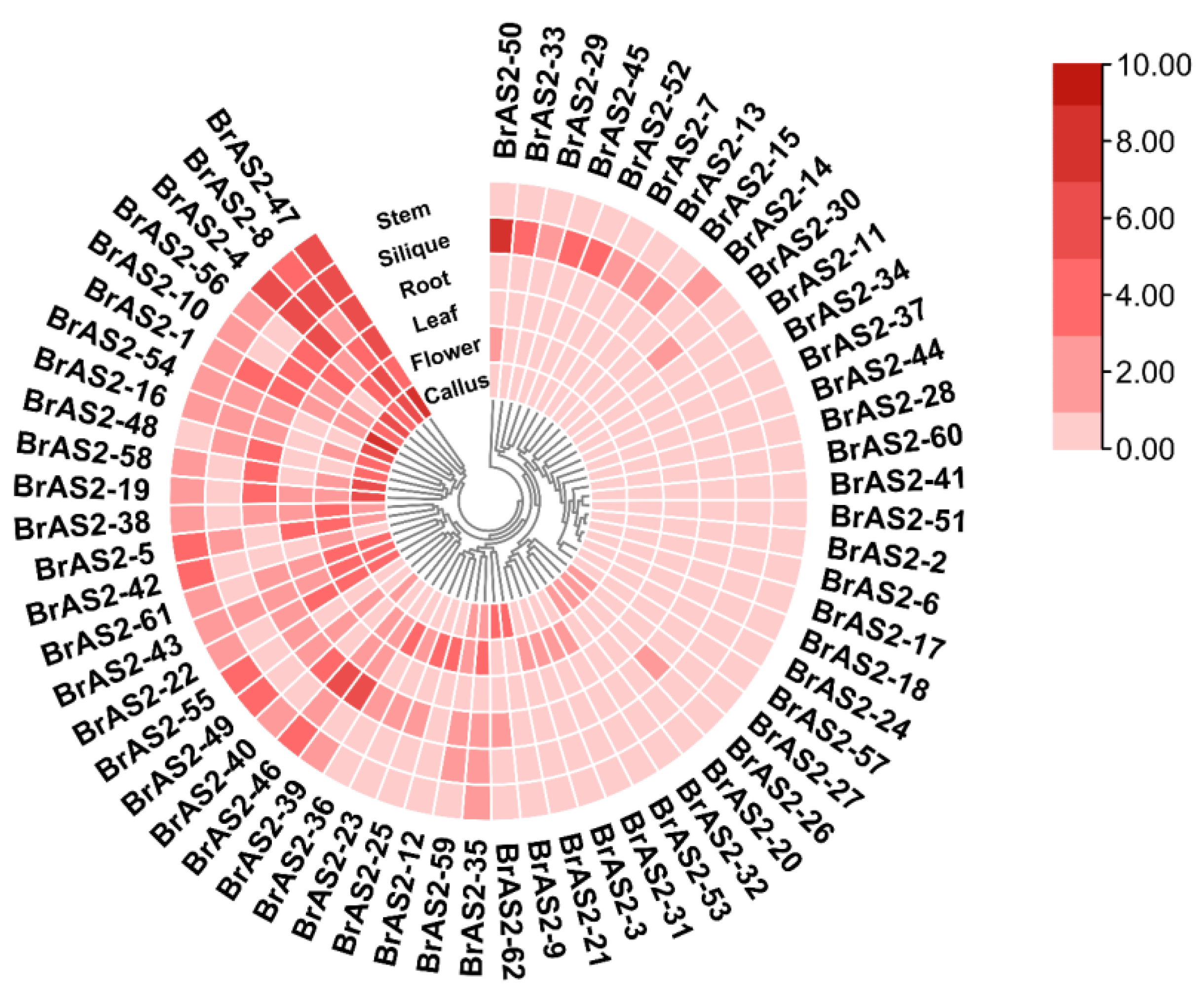
2.7. Tissue-Specific Expression Analysis
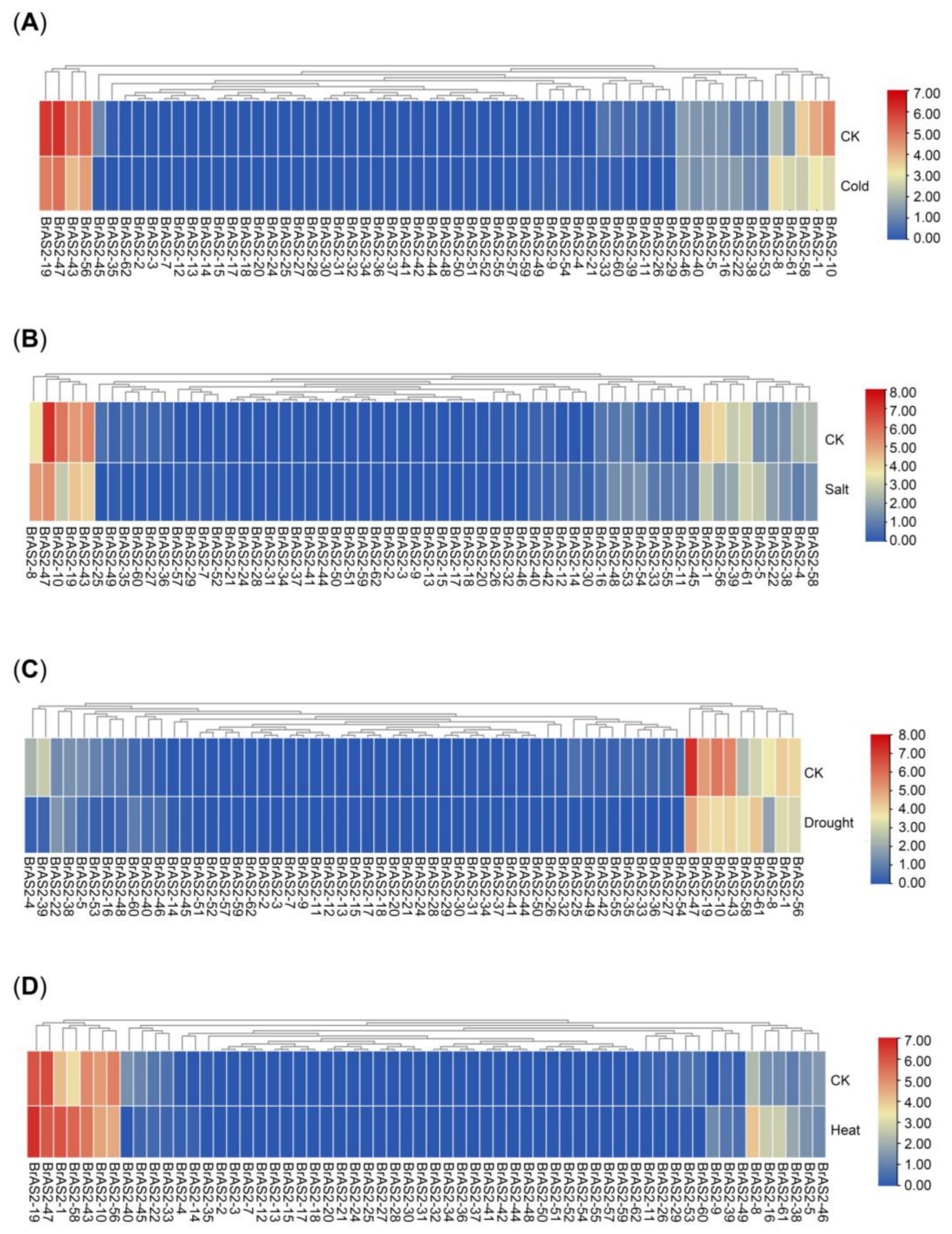
2.8. Expression Patterns in Response to Abiotic Stress Analysis
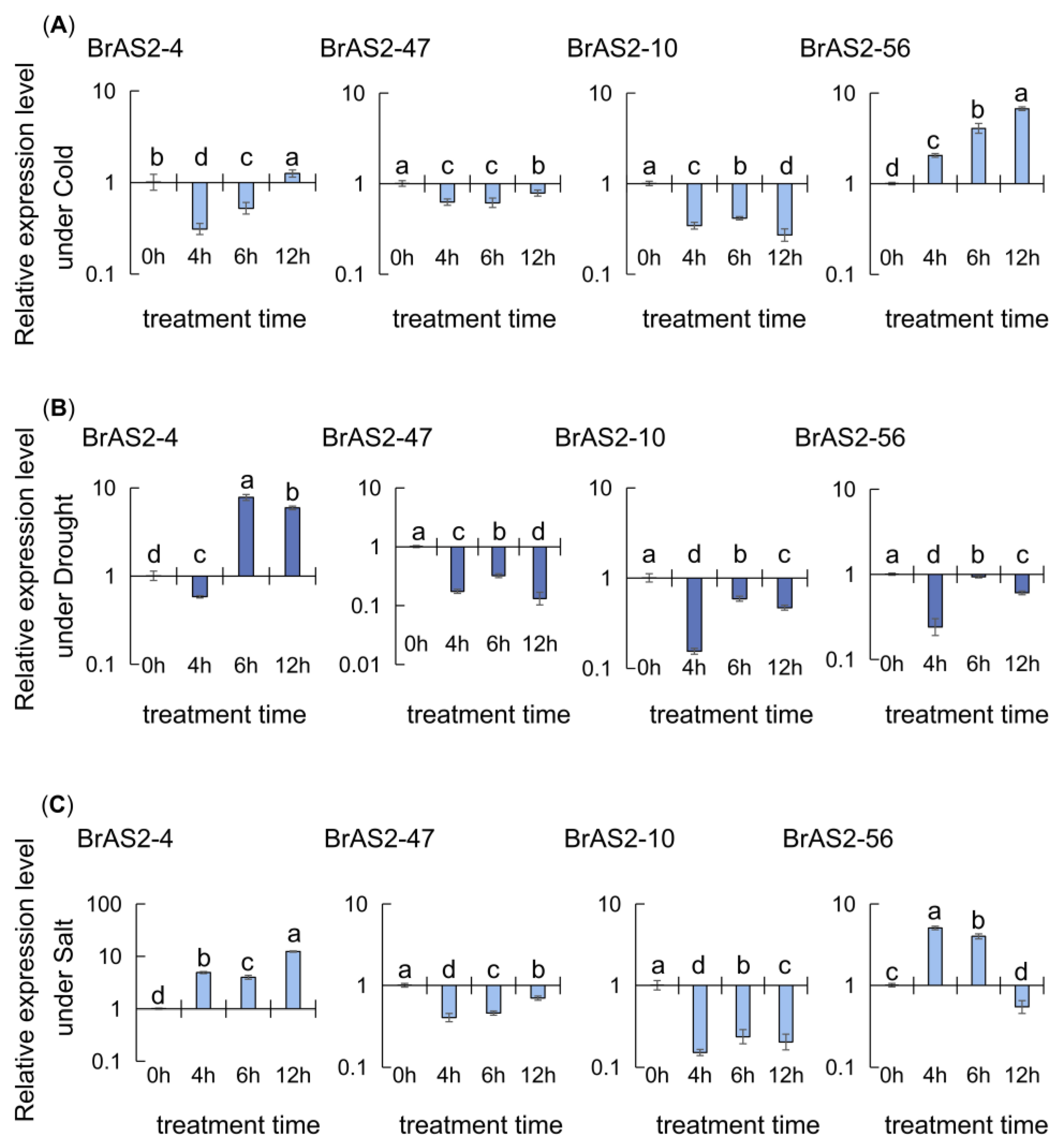
2.9. RT-qPCR Analysis
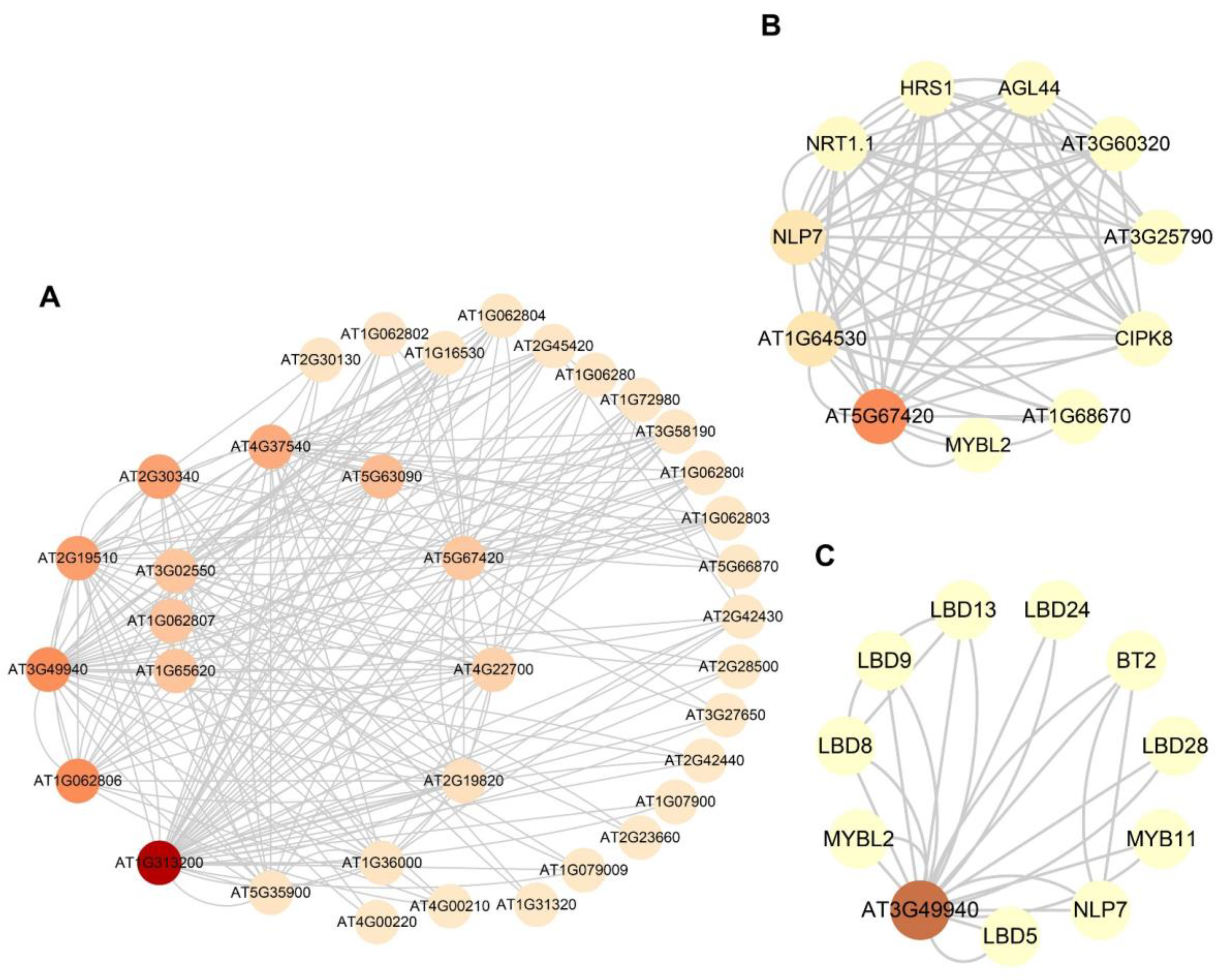
2.10. Prediction of Protein–Protein Interaction Network Analysis
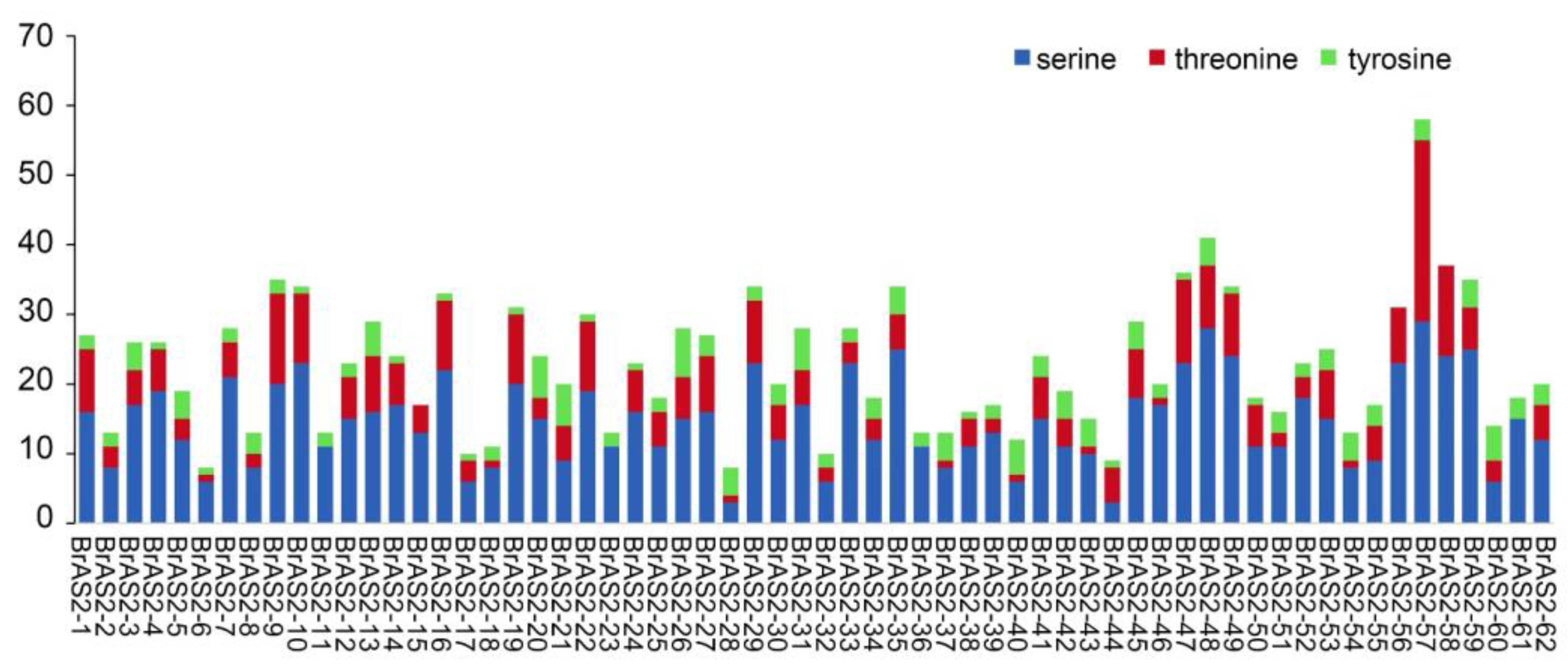
2.11. Phosphorylation Site Analysis
3. Discussion
4. Materials and Methods
4.1. Identification of AS2 Family Gene Members in B. rapa
4.2. Chromosomal Localization and Collinearity Analysis
4.3. Subcellular Localization Analysis
4.4. Phylogenetic Tree Construction, Conserved Motifs, and Gene Structural Analysis
4.5. Cis-Element and Gene Ontology Analyses
4.6. Plant Material, Stress Treatments, and Total RNA Extraction
4.7. Expression Patterns Analysis
4.8. RT-qPCR Analysis
4.9. Statistical Analysis
4.10. Protein–Protein Interaction Networks and Phosphorylation Site Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shuai, B.; Reynaga-Peña, C.G.; Springer, P.S. The Lateral Organ Boundaries Gene Defines a Novel, Plant-Specific Gene Family. Plant Physiol. 2002, 129, 747–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwakawa, H.; Ueno, Y.; Semiarti, E.; Onouchi, H.; Kojima, S.; Tsukaya, H.; Hasebe, M.; Soma, T.; Ikezaki, M.; Machida, C.; et al. The ASYMMETRIC LEAVES2 Gene of Arabidopsis Thaliana, Required for Formation of a Symmetric Flat Leaf Lamina, Encodes a Member of a Novel Family of Proteins Characterized by Cysteine Repeats and a Leucine Zipper. Plant Cell Physiol. 2002, 43, 467–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, B.-J.; Wang, J.; Lin, S.; Tian, Z.; Zhou, K.; Luan, H.-Y.; Lyu, C.; Zhang, X.-Z.; Xu, R.-G. A Genome-Wide Analysis of the ASYMMETRIC LEAVES2/LATERAL ORGAN BOUNDARIES (AS2/LOB) Gene Family in Barley (Hordeum vulgare L.). J. Zhejiang Univ. Sci. B 2016, 17, 763–774. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Fang, G.Y.; He, H.; Chen, J. Genome-Wide Identification, Evolutionary Analysis and Expression Profiles of LATERAL ORGAN BOUNDARIES DOMAIN Gene Family in Lotus Japonicus and Medicago Truncatula. PLoS ONE 2016, 11, e0161901. [Google Scholar] [CrossRef] [Green Version]
- Grimplet, J.; Pimentel, D.; Agudelo-Romero, P.; Martinez-Zapater, J.M.; Fortes, A.M. The LATERAL ORGAN BOUNDARIES Domain Gene Family in Grapevine: Genome-Wide Characterization and Expression Analyses during Developmental Processes and Stress Responses. Sci. Rep. 2017, 7, 15968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Cao, M.; Chen, X.; Ye, M.; Zhao, P.; Nan, Y.; Li, W.; Zhang, C.; Kong, L.; Kong, N.; et al. Genome-Wide Analysis of the Lateral Organ Boundaries Domain (LBD) Gene Family in Solanum Tuberosum. Int. J. Mol. Sci. 2019, 20, 5360. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Shi, G.; Du, H.; Wang, H.; Zhang, Z.; Hu, D.; Wang, J.; Huang, F.; Yu, D. Genome-Wide Analysis of Soybean LATERAL ORGAN BOUNDARIES Domain-Containing Genes: A Functional Investigation of GmLBD12. Plant Genome 2017, 10. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Huang, Z.; Ma, R.; Ramakrishnan, M.; Chen, J.; Zhang, Z.; Yrjälä, K. Genome-wide identification and expression analysis of LBD transcription factor genes in Moso bamboo (Phyllostachys edulis). BMC Plant Biol. 2021, 21, 296. [Google Scholar] [CrossRef]
- Guo, Z.; Xu, H.; Lei, Q.; Du, J.; Li, C.; Wang, C.; Yang, Y.; Yang, Y.; Sun, X. The Arabidopsis transcription factor LBD15 mediates ABA signaling and tolerance of water-deficit stress by regulating ABI4 expression. Plant J. 2020, 104, 510–521. [Google Scholar] [CrossRef]
- Ariel, F.D.; Diet, A.; Crespi, M.; Chan, R.L. The LOB-like Transcription Factor Mt LBD1 Controls Medicago Truncatula Root Architecture under Salt Stress. Plant Signal Behav. 2010, 5, 1666–1668. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Xiong, J.; Zhang, W.; Guan, H.; Zheng, D.; Xiong, H.; Jia, L.; Hu, Y.; Zhou, H.; Wen, Y.; et al. ZmLBD5, a class-II LBD gene, negatively regulates drought tolerance by impairing abscisic acid synthesis. Plant J. 2022, 112, 1364–1376. [Google Scholar] [CrossRef] [PubMed]
- Jeon, E.; Kang, N.Y.; Cho, C.; Seo, P.J.; Suh, M.C.; Kim, J. LBD14/ASL17 Positively Regulates Lateral Root Formation and is Involved in ABA Response for Root Architecture in Arabidopsis. Plant Cell Physiol. 2017, 58, 2190–2201. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Zhang, J.; Xu, J.; Li, Y.; Guo, L.; Wang, Z.; Zhang, X.; Zhao, B.; Guo, Y.-D.; Zhang, N. CRISPR/Cas9 targeted mutagenesis of SlLBD40, a lateral organ boundaries domain transcription factor, enhances drought tolerance in tomato. Plant Sci. 2020, 301, 110683. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Z.; Ma, B.; Hou, Q.; Wan, X. Phylogeny and Functions of LOB Domain Proteins in Plants. Int. J. Mol. Sci. 2020, 21, 2278. [Google Scholar] [CrossRef] [Green Version]
- Thatcher, L.F.; Powell, J.J.; Aitken, E.A.B.; Kazan, K.; Manners, J.M. The lateral organ boundaries domain transcription factor LBD20 functions in Fusarium wilt Susceptibility and jasmonate signaling in Arabidopsis. Plant Physiol. 2012, 160, 407–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Yan, H.; Liu, Y.; Yi, Y. Genome-Wide Analysis of LATERAL ORGAN BOUNDARIES DOMAIN-in Physcomitrella Patens and Stress Responses. Genes Genom. 2020, 42, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Azuma, A.; Yakushiji, H.; Koshita, Y.; Kobayashi, S. Flavonoid biosynthesis-related genes in grape skin are differentially regulated by temperature and light conditions. Planta 2012, 236, 1067–1080. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, S.A.; Levizou, E.; Ntatsi, G.; Fernandes, Â.; Petrotos, K.; Akoumianakis, K.; Barros, L.; Ferreira, I.C. Salinity effect on nutritional value, chemical composition and bioactive compounds content of Cichorium spinosum L. Food Chem. 2017, 214, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Drought stress enhances nutritional and bioactive compounds, phenolic acids and antioxidant capacity of Amaranthus leafy vegetable. BMC Plant Biol. 2018, 18, 258. [Google Scholar] [CrossRef] [Green Version]
- Gharibi, S.; Tabatabaei, B.E.S.; Saeidi, G.; Talebi, M.; Matkowski, A. The effect of drought stress on polyphenolic compounds and expression of flavonoid biosynthesis related genes in Achillea pachycephala Rech. f. Phytochemistry 2019, 162, 90–98. [Google Scholar] [CrossRef]
- Rubin, G.; Tohge, T.; Matsuda, F.; Saito, K.; Scheible, W.R. Members of the LBD Family of Transcription Factors Repress Anthocyanin Synthesis and Affect Additional Nitrogen Responses in Arabidopsis. Plant Cell 2009, 21, 3567–3584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albinsky, D.; Kusano, M.; Higuchi, M.; Hayashi, N.; Kobayashi, M.; Fukushima, A.; Mori, M.; Ichikawa, T.; Matsui, K.; Kuroda, H.; et al. Metabolomic Screening Applied to Rice FOX Arabidopsis, Lines Leads to the Identification of a Gene-Changing Nitrogen Metabolism. Mol. Plant 2010, 3, 125–142. [Google Scholar] [CrossRef]
- Li, C.; Zhu, S.; Zhang, H.; Chen, L.; Cai, M.; Wang, J.; Chai, J.; Wu, F.; Cheng, Z.; Guo, X.; et al. OsLBD37 and OsLBD38, two class II type LBD proteins, are involved in the regulation of heading date by controlling the expression of Ehd1 in rice. Biochem. Biophys. Res. Commun. 2017, 486, 720–725. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; He, Y.; He, W.; Su, H.; Wang, Y.; Hong, G.; Xu, P. Structural and functional insights into the LBD family involved in abiotic stress and flavonoid synthases in Camellia sinensis. Sci. Rep. 2019, 9, 15651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwakawa, H.; Iwasaki, M.; Kojima, S.; Ueno, Y.; Soma, T.; Tanaka, H.; Semiarti, E.; Machida, Y.; Machida, C. Expression of the ASYMMETRIC LEAVES2 Gene in the Adaxial Domain of Arabidopsis Leaves Represses Cell Proliferation in This Domain and Is Critical for the Development of Properly Expanded Leaves. Plant J. 2007, 51, 173–184. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Y.; Kohalmi, S.E.; Amyot, L.; Hannoufa, A. SQUAMOSA PROMOTER BINDING PROTEIN-LIKE 2 controls floral organ development and plant fertility by activating ASYMMETRIC LEAVES 2 in Arabidopsis thaliana. Plant Mol. Biol. 2016, 92, 661–674. [Google Scholar] [CrossRef]
- Borghi, L.; Bureau, M.; Simon, R. Arabidopsis JAGGED LATERAL ORGANS Is Expressed in Boundaries and Coordinates KNOX and PIN Activity. Plant Cell 2007, 19, 1795–1808. [Google Scholar] [CrossRef] [Green Version]
- Ori, N.; Eshed, Y.; Chuck, G.; Bowman, J.L.; Hake, S. Mechanisms That Control Knox Gene Expression in the Arabidopsis Shoot. Development 2000, 127, 5523–5532. [Google Scholar] [CrossRef]
- Semiarti, E.; Ueno, Y.; Tsukaya, H.; Iwakawa, H.; Machida, C.; Machida, Y. The ASYMMETRIC LEAVES2 Gene of Arabidopsis Thaliana Regulates Formation of a Symmetric Lamina, Establishment of Venation and Repression of Meristem-Related Homeobox Genes in Leaves. Development 2001, 128, 1771–1783. [Google Scholar] [CrossRef]
- Byrne, M.E.; Simorowski, J.; Martienssen, R.A. ASYMMETRIC LEAVES1 Reveals Knox Gene Redundancy in Arabidopsis. Development 2002, 129, 1957–1965. [Google Scholar] [CrossRef]
- Guo, M.; Thomas, J.; Collins, G.; Timmermans, M.C.P. Direct Repression of KNOX Loci by the ASYMMETRIC LEAVES1 Complex of Arabidopsis. Plant Cell 2008, 20, 48–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, B.; Li, Z.; Zhu, Y.; Wang, H.; Ma, H.; Dong, A.; Huang, H. Arabidopsis Genes AS1, AS2, and JAG Negatively Regulate Boundary-Specifying Genes to Promote Sepal and Petal Development. Plant Physiol. 2008, 146, 566–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Hou, H.; Zhang, Y.; Hou, X. Overexpression of a Pak Choi Gene, BcAS2, Causes Leaf Curvature in Arabidopsis Thaliana. Genes 2021, 12, 102. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Schranz, M.E. Network approaches for plant phylogenomic synteny analysis. Curr. Opin. Plant Biol. 2017, 36, 129–134. [Google Scholar] [CrossRef]
- Alberts, B. The Cell as a Collection of Protein Machines: Preparing the next Generation of Molecular Biologists. Cell 1998, 92, 291–294. [Google Scholar] [CrossRef] [Green Version]
- Konishi, M.; Yanagisawa, S. The Role of Protein-Protein Interactions Mediated by the PB1 Domain of NLP Transcription Factors in Nitrate-Inducible Gene Expression. BMC Plant Biol. 2019, 19, 90. [Google Scholar] [CrossRef] [Green Version]
- Konishi, M.; Okitsu, T.; Yanagisawa, S. Nitrate-Responsive NIN-like Protein Transcription Factors Perform Unique and Redundant Roles in Arabidopsis. J. Exp. Bot. 2021, 72, 5735–5750. [Google Scholar] [CrossRef]
- Xie, Y.; Tan, H.; Ma, Z.; Huang, J. DELLA Proteins Promote Anthocyanin Biosynthesis via Sequestering MYBL2 and JAZ Suppressors of the MYB/bHLH/WD40 Complex in Arabidopsis thaliana. Mol. Plant 2016, 9, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Feng, J.; Wang, R.; Liu, H.; Yang, H.; Rodriguez, P.L.; Qin, H.; Liu, X.; Wang, D. HRS1 acts as a negative regulator of abscisic acid signaling to promote timely germination of Arabidopsis seeds. PLoS ONE 2012, 7, e35764. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zhou, L.; Li, Y.; Zhang, D.; Gao, Y. Plant NIGT1/HRS1/HHO Transcription Factors: Key Regulators with Multiple Roles in Plant Growth, Development, and Stress Responses. Int. J. Mol. Sci. 2021, 22, 8685. [Google Scholar] [CrossRef]
- Ding, Y.; Yang, H.; Wu, S.; Fu, D.; Li, M.; Gong, Z.; Yang, S. CPK28-NLP7 Module Integrates Cold-Induced Ca2+ Signal and Transcriptional Reprogramming in Arabidopsis. Sci. Adv. 2022, 8, eabn7901. [Google Scholar] [CrossRef]
- Mandadi, K.K.; Misra, A.; Ren, S.; McKnight, T.D. BT2, a BTB protein, mediates multiple responses to nutrients, stresses, and hormones in Arabidopsis. Plant Physiol. 2009, 150, 1930–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stracke, R.; Jahns, O.; Keck, M.; Tohge, T.; Niehaus, K.; Fernie, A.R.; Weisshaar, B. Analysis of PRODUCTION OF FLAVONOL GLYCOSIDES-Dependent Flavonol Glycoside Accumulation in Arabidopsis Thaliana Plants Reveals MYB11-, MYB12- and MYB111- Independent Flavonol Glycoside Accumulation. New Phytol. 2010, 188, 985–1000. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, S.J.; James, D.E.; Mann, M. Protein Phosphorylation: A Major Switch Mechanism for Metabolic Regulation. Trends Endocrinol. Metab. 2015, 26, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Yaffe, M.B.; Smerdon, S.J. The use of in vitro peptide-library screens in the analysis of phosphoserine/threonine-binding domain structure and function. Annu. Rev. Biophys. Biomol. Struct. 2004, 33, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Kole, C. The Brassica rapa Genome; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- The Brassica rapa Genome Sequencing Project Consortium; Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.-H.; Bancroft, I.; et al. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Mandáková, T.; Wu, J.; Xie, Q.; Lysak, M.A.; Wang, X. Deciphering the diploid ancestral genome of the mesohexaploid Brassica rapa. Plant Cell 2013, 25, 1541–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Luo, F.; Hochholdinger, F. LOB Domain Proteins: Beyond Lateral Organ Boundaries. Trends Plant Sci. 2016, 21, 159–167. [Google Scholar] [CrossRef]
- Feng, Z.; Zhu, J.; Du, X.; Cui, X. Effects of three auxin-inducible LBD members on lateral root formation in Arabidopsis thaliana. Planta 2012, 236, 1227–1237. [Google Scholar] [CrossRef]
- Lee, H.W.; Cho, C.; Kim, J. Lateral Organ Boundaries Domain16 and 18 Act Downstream of the AUXIN1 and LIKE-AUXIN3 Auxin Influx Carriers to Control Lateral Root Development in Arabidopsis. Plant Physiol. 2015, 168, 1792–1806. [Google Scholar] [CrossRef] [Green Version]
- Scheible, W.-R.; Morcuende, R.; Czechowski, T.; Fritz, C.; Osuna, D.; Palacios-Rojas, N.; Schindelasch, D.; Thimm, O.; Udvardi, M.; Stitt, M. Genome-Wide Reprogramming of Primary and Secondary Metabolism, Protein Synthesis, Cellular Growth Processes, and the Regulatory Infrastructure of Arabidopsis in Response to Nitrogen. Plant Physiol. 2004, 136, 2483–2499. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, Y.; Iwakawa, H.; Machida, Y.; Machida, C. Characterization of genes in the ASYMMETRIC LEA VES2/LATERAL ORGAN BOUNDARIES (AS2/LOB) family in Arabidopsis thaliana, and functional and molecular comparisons between AS2 and other family members. Plant J. 2009, 58, 525–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.; Wu, H.; Ma, S.; Xiang, D.; Liu, R.; Xiong, L. OsJAZ1 Attenuates Drought Resistance by Regulating JA and ABA Signaling in Rice. Front. Plant Sci. 2017, 8, 2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraus, E.C.; Stout, M.J. Seed treatment using methyl jasmonate induces resistance to rice water weevil but reduces plant growth in rice. PLoS ONE 2019, 14, e0222800. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Zhang, W.; Zhang, Y.; Zhang, X.; Lang, D.; Zhang, X. The roles of methyl jasmonate to stress in plants. Funct. Plant Biol. 2019, 46, 197–212. [Google Scholar] [CrossRef]
- Ding, F.; Wang, X.; Li, Z.; Wang, M. Jasmonate Positively Regulates Cold Tolerance by Promoting ABA Biosynthesis in Tomato. Plants 2022, 12, 60. [Google Scholar] [CrossRef] [PubMed]
- Nie, G.; Zhou, J.; Jiang, Y.; He, J.; Wang, Y.; Liao, Z.; Appiah, C.; Li, D.; Feng, G.; Huang, L.; et al. Transcriptome characterization of candidate genes for heat tolerance in perennial ryegrass after exogenous methyl Jasmonate application. BMC Plant Biol. 2022, 22, 68. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.-K. Abiotic Stress Responses in Plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef]
- Deutscher, J.; Saier, M.H., Jr. Ser/Thr/Tyr protein phosphorylation in bacteria—For long time neglected, now well established. J. Mol. Microbiol. Biotechnol. 2005, 9, 125–131. [Google Scholar] [CrossRef]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive Oxygen Species Signalling in Plant Stress Responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB Bioinformatics Resource Portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Rombauts, S.; Déhais, P.; Van Montagu, M.; Rouzé, P. PlantCARE, a Plant Cis-Acting Regulatory Element Database. Nucleic Acids Res. 1999, 27, 295–296. [Google Scholar] [CrossRef] [Green Version]
- Dennis, G.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 2003, 4, R60. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Chromosome Distribution | Classification | Protein Length (aa) | MW (Da) | PI | A. thaliana ID |
|---|---|---|---|---|---|---|---|
| BrAS2-1 | Bra011772 | A01 (631422–632295) | IIa | 237 | 26,167.73 | 8.14 | AT4G37540 |
| BrAS2-2 | Bra030013 | A01 (15744550–15745095) | Ia | 181 | 20,472.25 | 6.28 | AT3G50510 |
| BrAS2-3 | Bra021513 | A01 (24855421–24856203) | Id | 260 | 29,654.86 | 5.19 | AT3G13850 |
| BrAS2-4 | Bra021433 | A01 (25233613–25234491) | IIb | 264 | 28,181.17 | 8.9 | AT3G02550 |
| BrAS2-5 | Bra039733 | A02 (9622401–9623009) | Ia | 202 | 22,093.85 | 7.1 | AT1G65620 |
| BrAS2-6 | Bra008062 | A02 (12966840–12967397) | Id | 185 | 21,098.41 | 8.3 | AT1G72980 |
| BrAS2-7 | Bra008514 | A02 (16097898–16100589) | Ic | 224 | 23,732.84 | 6.53 | AT4G00220 |
| BrAS2-8 | Bra033019 | A02 (21869558–21869956) | Ia | 132 | 14,678.52 | 6.89 | AT3G27650 |
| BrAS2-9 | Bra032938 | A02 (22395463–22401919) | Ia | 294 | 32,145.26 | 4.78 | AT3G26660 |
| BrAS2-10 | Bra031833 | A02 (27641733–27642526) | IIa | 241 | 25,985.18 | 8.46 | AT5G67420 |
| BrAS2-11 | Bra022780 | A03 (7018824–7021941) | Ia | 188 | 20,973.78 | 6.21 | AT2G30130 |
| BrAS2-12 | Bra000188 | A03 (9790598–9791563) | Ib | 199 | 21,455.45 | 8.8 | AT2G40470 |
| BrAS2-13 | Bra000257 | A03 (10204676–10206488) | Ic | 238 | 25,950.97 | 6.88 | AT2G42430 |
| BrAS2-14 | Bra000491 | A03 (11383294–11384774) | Ia | 231 | 25,396.57 | 4.9 | AT2G28500 |
| BrAS2-15 | Bra001087 | A03 (14672898–14673260) | Ic | 120 | 12,555.11 | 10.63 | AT3G03760 |
| BrAS2-16 | Bra012913 | A03 (21540949–21541767) | IIa | 241 | 26,607.1 | 6.89 | AT3G49940 |
| BrAS2-17 | Bra019365 | A03 (24669629–24670232) | Id | 157 | 17,587.86 | 4.73 | AT5G35900 |
| BrAS2-18 | Bra019364 | A03 (24677058–24677660) | Id | 156 | 17,566.68 | 4.72 | AT5G63090 |
| BrAS2-19 | Bra017831 | A03 (30772548–30773423) | IIa | 232 | 25,301.1 | 9.1 | AT4G37540 |
| BrAS2-20 | Bra014581 | A04 (1568864–1569640) | Ic | 219 | 24,375.97 | 6.28 | AT3G58190 |
| BrAS2-21 | Bra032153 | A04 (10493828–10494754) | Ia | 308 | 34,410.77 | 4.83 | AT2G23660 |
| BrAS2-22 | Bra035698 | A04 (12731349–12732755) | Ia | 229 | 25,158.4 | 4.82 | AT2G28500 |
| BrAS2-23 | Bra021612 | A04 (13416815–13417633) | Ia | 190 | 21,127.98 | 5.91 | AT2G30130 |
| BrAS2-24 | Bra021737 | A04 (14176806–14179238) | Ic | 239 | 26,536.2 | 7.05 | AT2G31310 |
| BrAS2-25 | Bra016992 | A04 (17230175–17231282) | Ib | 201 | 21,738.62 | 7.68 | AT2G40470 |
| BrAS2-26 | Bra016877 | A04 (17780078–17781492) | Ic | 246 | 26,618.56 | 8.11 | AT2G42430 |
| BrAS2-27 | Bra016876 | A04 (17786570–17788024) | Ic | 264 | 29,442.97 | 6.29 | AT2G42440 |
| BrAS2-28 | Bra040312 | A04 (18539194–18539789) | Ic | 168 | 18,604.4 | 7.68 | AT2G45410 |
| BrAS2-29 | Bra040311 | A04 (18545871–18547692) | Ic | 259 | 27,132.39 | 8.19 | AT2G45420 |
| BrAS2-30 | Bra004572 | A05 (848386–849380) | Ib | 223 | 24,453.85 | 8.73 | AT2G40470 |
| BrAS2-31 | Bra004693 | A05 (1378014–1379047) | Ic | 244 | 26,262.19 | 8.15 | AT2G42430 |
| BrAS2-32 | Bra004908 | A05 (2423168–2423825) | Ic | 188 | 20,738.62 | 6.29 | AT2G45410 |
| BrAS2-33 | Bra004910 | A05 (2431903–2433701) | Ic | 253 | 26,065.29 | 8.51 | AT2G45420 |
| BrAS2-34 | Bra018260 | A05 (6977723–6978406) | Ic | 193 | 21,602.68 | 5.93 | AT2G31310 |
| BrAS2-35 | Bra018320 | A05 (7426670–7428607) | Ib | 236 | 25,340.52 | 8.88 | AT2G30340 |
| BrAS2-36 | Bra018335 | A05 (7526963–7527792) | Ia | 189 | 21,079 | 6.03 | AT2G30130 |
| BrAS2-37 | Bra027392 | A05 (20677617–20678129) | Id | 170 | 19,756.71 | 4.5 | AT3G13850 |
| BrAS2-38 | Bra034867 | A05 (21803639–21804157) | Ib | 172 | 18,765.48 | 6.58 | AT3G11090 |
| BrAS2-39 | Bra018675 | A06 (2680223–2681288) | Ia | 206 | 22,866.92 | 4.96 | AT2G28500 |
| BrAS2-40 | Bra026042 | A06 (6372131–6373653) | Ia | 163 | 18,075.77 | 8.53 | AT1G16530 |
| BrAS2-41 | Bra018102 | A06 (10251078–10252128) | Id | 318 | 36,165.12 | 5.07 | AT3G47870 |
| BrAS2-42 | Bra038606 | A06 (14136819–14137364) | Ia | 181 | 19,910.5 | 8.29 | AT5G63090 |
| BrAS2-43 | Bra025294 | A06 (22339538–22339936) | Ia | 132 | 14,563.43 | 7.58 | AT3G27650 |
| BrAS2-44 | Bra025217 | A06 (22711432–22712256) | Ia | 107 | 11,876.75 | 6.57 | AT3G26620 |
| BrAS2-45 | Bra036436 | A07 (499213–500551) | Ic | 275 | 29,136.32 | 7.27 | AT3G03760 |
| BrAS2-46 | Bra014907 | A07 (5302258–5303262) | Ia | 172 | 18,669.09 | 7.61 | AT1G31320 |
| BrAS2-47 | Bra012164 | A07 (9636805–9637668) | IIa | 256 | 27,868.24 | 8.4 | AT5G67420 |
| BrAS2-48 | Bra012112 | A07 (9939966–9940982) | Ia | 338 | 37,528.62 | 6.54 | AT5G66870 |
| BrAS2-49 | Bra011942 | A07 (11079984–11081436) | Ia | 233 | 25,579.89 | 4.88 | AT2G28500 |
| BrAS2-50 | Bra004315 | A07 (17939314–17940220) | IIb | 237 | 25,763.1 | 8.05 | AT1G68510 |
| BrAS2-51 | Bra030647 | A08 (20962015–20962608) | Id | 197 | 22,476.04 | 9.68 | AT1G06280 |
| BrAS2-52 | Bra037323 | A09 (1067402–1069396) | Ic | 224 | 23,588.82 | 6.43 | AT4G00220 |
| BrAS2-53 | Bra037322 | A09 (1073182–1074567) | Ic | 222 | 24,234.34 | 6.16 | AT4G00210 |
| BrAS2-54 | Bra039072 | A09 (1261020–1261415) | Ia | 131 | 14,582.52 | 7.6 | AT3G27650 |
| BrAS2-55 | Bra035860 | A09 (3268177–3268707) | Ia | 176 | 19,265.76 | 8.56 | AT5G63090 |
| BrAS2-56 | Bra037847 | A09 (3873152–3873956) | IIa | 236 | 25,719.08 | 9.03 | AT5G67420 |
| BrAS2-57 | Bra037142 | A09 (4287000–4288184) | Ia | 394 | 44,228.21 | 5.62 | AT5G66870 |
| BrAS2-58 | Bra036040 | A09 (24873579–24874625) | IIa | 245 | 26,699.27 | 6.82 | AT3G49940 |
| BrAS2-59 | Bra007385 | A09 (28936037–28936836) | Ic | 225 | 24,753.19 | 6.01 | AT3G58190 |
| BrAS2-60 | Bra026716 | A09 (33360480–33361740) | Ia | 164 | 18,506.26 | 9.15 | AT1G16530 |
| BrAS2-61 | Bra031599 | A09 (35646044–35647161) | Ia | 207 | 23,134.33 | 4.87 | AT2G28500 |
| BrAS2-62 | Bra009161 | A10 (15474429–15475061) | Ic | 183 | 20,624.28 | 5.72 | AT5G06080 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Q.; Wu, X.; Zhang, X.; Ji, Z.; Cao, Y.; Duan, Q.; Huang, J. Genome-Wide Identification and Expression Analysis of AS2 Genes in Brassica rapa Reveal Their Potential Roles in Abiotic Stress. Int. J. Mol. Sci. 2023, 24, 10534. https://doi.org/10.3390/ijms241310534
Jiang Q, Wu X, Zhang X, Ji Z, Cao Y, Duan Q, Huang J. Genome-Wide Identification and Expression Analysis of AS2 Genes in Brassica rapa Reveal Their Potential Roles in Abiotic Stress. International Journal of Molecular Sciences. 2023; 24(13):10534. https://doi.org/10.3390/ijms241310534
Chicago/Turabian StyleJiang, Qiwei, Xiaoyu Wu, Xiaoyu Zhang, Zhaojing Ji, Yunyun Cao, Qiaohong Duan, and Jiabao Huang. 2023. "Genome-Wide Identification and Expression Analysis of AS2 Genes in Brassica rapa Reveal Their Potential Roles in Abiotic Stress" International Journal of Molecular Sciences 24, no. 13: 10534. https://doi.org/10.3390/ijms241310534