
The Pannexin 1 Channel and the P2X7 Receptor Are in Complex Interplay to Regulate the Release of Soluble Ectonucleotidases in the Murine Bladder Lamina Propria
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of PANX1 on the Release of s-ENTDs in the Bladder LP
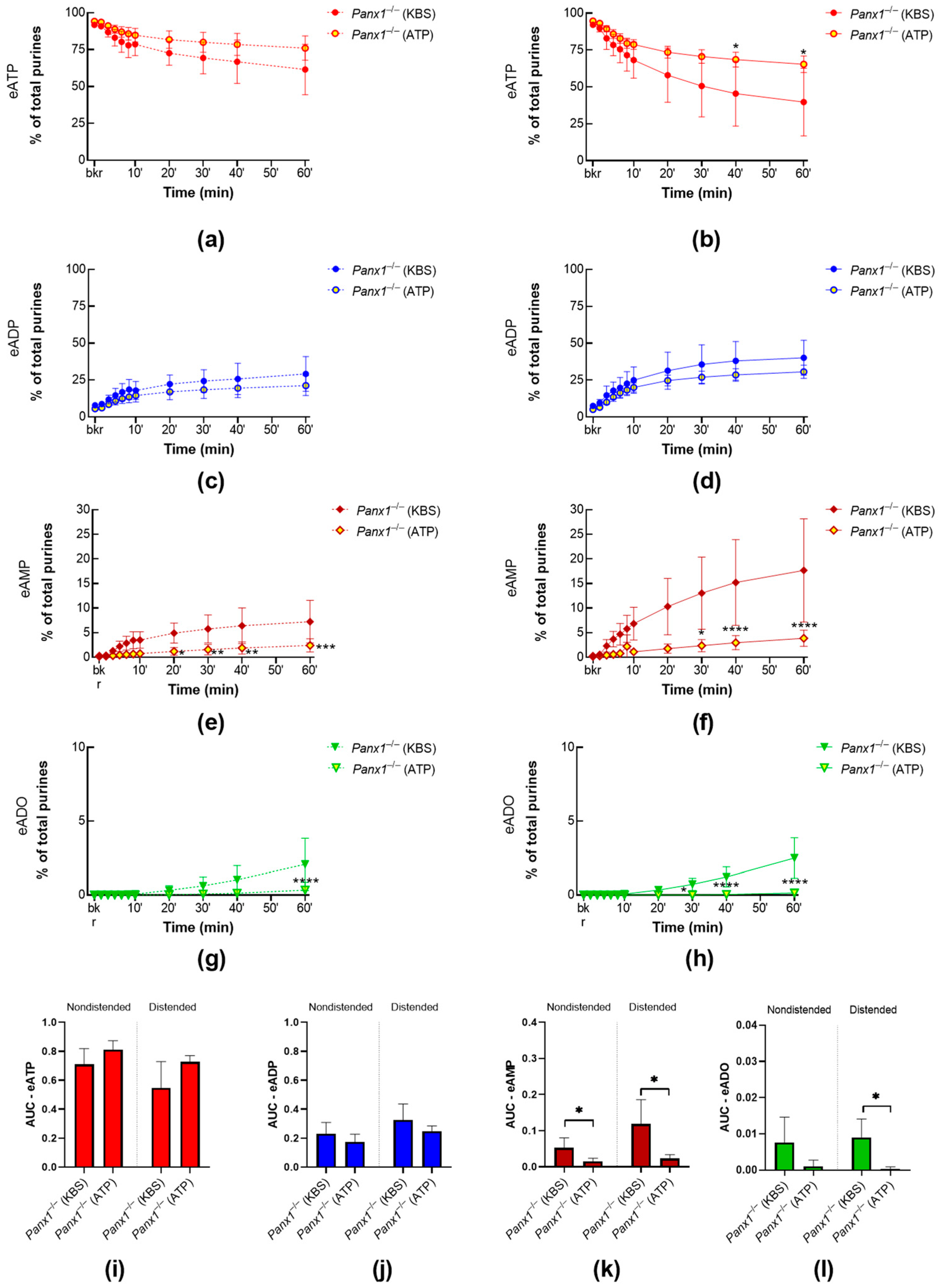
2.1.1. Panx1 Deletion Increases the Distension-Induced Release of s-ENTDs in the LP
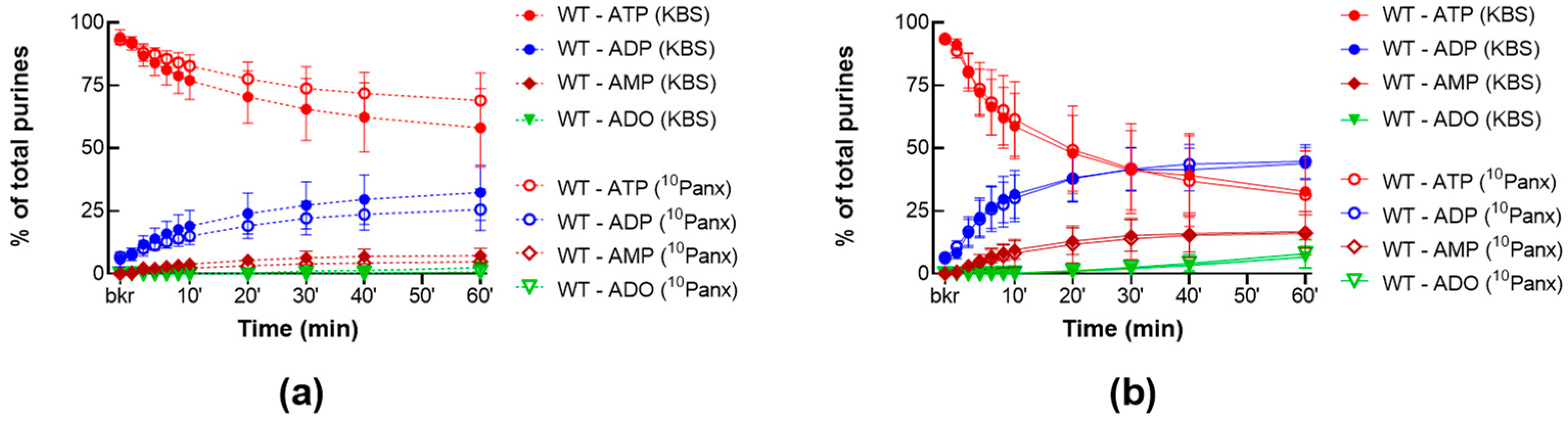
2.1.2. 10Panx Has no Effect on the Release of s-ENTDs in the LP
2.2. Role of P2X7R in the Release of s-ENTDs in LP
2.2.1. P2X7R Inhibition Does Not Alter the Constitutive nor the Distention-Induced Release of s-ENTDs in the LP of WT Denuded Bladders
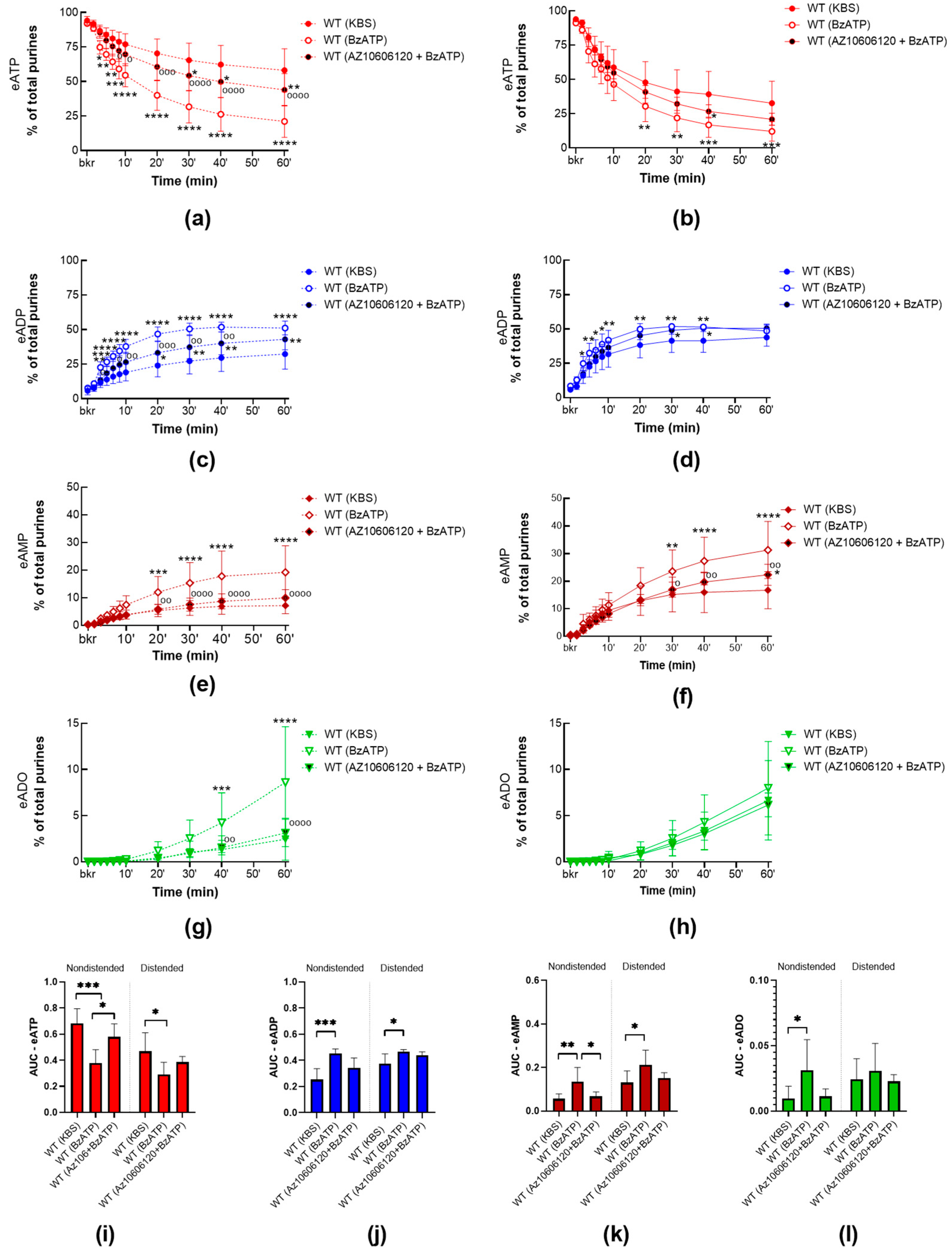
2.2.2. P2X7R Activation with BzATP Increases the Release of s-ENTDs in the LP of WT Bladders and Eliminates the Mechanosensitive Pattern of s-ENTDs Release
2.2.3. High Concentrations of ATP Facilitate the Release of s-ENTDs in the LP of WT Bladders
2.3. Interdependence of PANX1 and P2X7R in Mediating s-ENTDs Release in LP
2.3.1. BzATP Has no Effect on the Release of s-ENTDs in the LP of Panx1−/− Detrusor-Free Bladders
2.3.2. BzATP-Induced Release of s-ENTDS Is Blocked in the Presence of 10Panx
2.3.3. ATP-Induced Release of s-ENTDs during Distension Is Diminished in Panx1−/− Bladders
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Ethical Approval
4.3. RNA Isolation, Reverse Transcription, and RT-PCR
4.4. Detrusor-Free Bladder Preparation
4.5. Soluble/Releasable Nucleotidase Activity in the Lamina Propria of Detrusor-Free Bladder Preparations of WT and Panx1−/− Mice
4.6. Effect of Pharmacological Activation or Inhibition of P2X7R and/or PANX1 on the Release of Soluble Nucleotidases in the LP
4.7. Preparation of 1,N6-Etheno-Nucleotides
4.8. HPLC Analysis of 1,N6-Etheno-Nucleotides
4.9. Drugs and Reagents
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lewis, S.A.; Lewis, J.R. Kinetics of Urothelial ATP Release. Am. J. Physiol.-Ren. Physiol. 2006, 291, F332–F340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durnin, L.; Kwok, B.; Kukadia, P.; McAvera, R.; Corrigan, R.D.; Ward, S.M.; Zhang, Y.; Chen, Q.; Koh, S.D.; Sanders, K.M.; et al. An Ex Vivo Bladder Model with Detrusor Smooth Muscle Removed to Analyse Biologically Active Mediators Released from the Suburothelium. J. Physiol. 2019, 597, 1467–1485. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, D.R.; Kennedy, I.; Burton, T.J. ATP Is Released from Rabbit Urinary Bladder Epithelial Cells by Hydrostatic Pressure Changes-Possible Sensory Mechanism? J. Physiol. 1997, 505, 503–511. [Google Scholar] [CrossRef]
- Burnstock, G. Purinergic Signalling in the Urinary Tract in Health and Disease. Purinergic Signal. 2014, 10, 103–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aresta Branco, M.S.L.; Gutierrez Cruz, A.; Dayton, J.; Perrino, B.A.; Mutafova-Yambolieva, V.N. Mechanosensitive Hydrolysis of ATP and ADP in Lamina Propria of the Murine Bladder by Membrane-Bound and Soluble Nucleotidases. Front. Physiol. 2022, 13, 1185. [Google Scholar] [CrossRef]
- Todorov, L.D.; Mihaylova-Todorova, S.; Westfall, T.D.; Sneddon, P.; Kennedy, C.; Bjur, R.A.; Westfall, D.P. Neuronal Release of Soluble Nucleotidases and Their Role in Neurotransmitter Inactivation. Nature 1997, 387, 76–79. [Google Scholar] [CrossRef]
- Zimmermann, H.; Zebisch, M.; Sträter, N. Cellular Function and Molecular Structure of Ecto-Nucleotidases. Purinergic Signal. 2012, 8, 437. [Google Scholar] [CrossRef] [Green Version]
- Negoro, H.; Urban-Maldonado, M.; Liou, L.S.; Spray, D.C.; Thi, M.M.; Suadicani, S.O. Pannexin 1 Channels Play Essential Roles in Urothelial Mechanotransduction and Intercellular Signaling. PLoS ONE 2014, 9, e106269. [Google Scholar] [CrossRef]
- Beckel, J.M.; Daugherty, S.L.; Tyagi, P.; Wolf-Johnston, A.S.; Birder, L.A.; Mitchell, C.H.; de Groat, W.C. Pannexin 1 Channels Mediate the Release of ATP into the Lumen of the Rat Urinary Bladder. J. Physiol. 2015, 593, 1857–1871. [Google Scholar] [CrossRef] [Green Version]
- Pelegrin, P.; Surprenant, A. Pannexin-1 Mediates Large Pore Formation and Interleukin-1β Release by the ATP-Gated P2X7 Receptor. EMBO J. 2006, 25, 5071–5082. [Google Scholar] [CrossRef] [Green Version]
- Silverman, W.R.; de Rivero Vaccari, J.P.; Locovei, S.; Qiu, F.; Carlsson, S.K.; Scemes, E.; Keane, R.W.; Dahl, G. The Pannexin 1 Channel Activates the Inflammasome in Neurons and Astrocytes. J. Biol. Chem. 2009, 284, 18143–18151. [Google Scholar] [CrossRef] [Green Version]
- Poornima, V.; Madhupriya, M.; Kootar, S.; Sujatha, G.; Kumar, A.; Bera, A.K. P2X7 Receptor–Pannexin 1 Hemichannel Association: Effect of Extracellular Calcium on Membrane Permeabilization. J. Mol. Neurosci. 2012, 46, 585–594. [Google Scholar] [CrossRef]
- Xu, X.J.; Boumechache, M.; Robinson, L.E.; Marschall, V.; Gorecki, D.C.; Masin, M.; Murrell-Lagnado, R. Splice-Variants of the P2X7 Receptor Reveal Differential Agonist-Dependence and Functional Coupling with Pannexin-1. J. Cell Sci. 2012, 125, 3776–3789. [Google Scholar] [CrossRef] [Green Version]
- Boyce, A.K.J.; Swayne, L.A. P2X7 Receptor Cross-Talk Regulates ATP-Induced Pannexin 1 Internalization. Biochem. J. 2017, 474, 2133–2144. [Google Scholar] [CrossRef]
- Rhodes, G.; Segars, K.L.; Lee, Y.K.; Hutcheon, A.E.K.; Rich, C.B.; Trinkaus-Randall, V. Pannexin1: Role as a Sensor to Injury Is Attenuated in Pretype 2 Corneal Diabetic Epithelium. Anal. Cell. Pathol. 2021, 2021, 4793338. [Google Scholar] [CrossRef]
- Bravo, D.; Zepeda-Morales, K.; Maturana, C.J.; Retamal, J.S.; Hernández, A.; Pelissier, T.; Barra, R.; Sáez-Briones, P.; Burgos, H.; Constandil, L. NMDA and P2X7 Receptors Require Pannexin 1 Activation to Initiate and Maintain Nociceptive Signaling in the Spinal Cord of Neuropathic Rats. Int. J. Mol. Sci. 2022, 23, 6705. [Google Scholar] [CrossRef]
- Sosinsky, G.E.; Boassa, D.; Dermietzel, R.; Duffy, H.S.; Laird, D.W.; MacVicar, B.; Naus, C.C.; Penuela, S.; Scemes, E.; Spray, D.C.; et al. Pannexin Channels Are Not Gap Junction Hemichannels. Channels 2011, 5, 193–197. [Google Scholar] [CrossRef]
- Bao, L.; Locovei, S.; Dahl, G. Pannexin Membrane Channels Are Mechanosensitive Conduits for ATP. FEBS Lett. 2004, 572, 65–68. [Google Scholar] [CrossRef] [Green Version]
- Iglesias, R.; Locovei, S.; Roque, A.; Alberto, A.P.; Dahl, G.; Spray, D.C.; Scemes, E. P2X 7 Receptor-Pannexin1 Complex: Pharmacology and Signaling. Am. J. Physiol.-Cell Physiol. 2008, 295, C752–C760. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Compan, V.; Zheng, W.; Martin, E.; North, R.A.; Verkhratsky, A.; Surprenant, A. Pannexin 1 Forms an Anion-Selective Channel. Pflugers Arch. 2012, 463, 585–592. [Google Scholar] [CrossRef]
- Isakson, B.E.; Thompson, R.J. Pannexin-1 as a Potentiator of Ligand-Gated Receptor Signaling. Channels 2014, 8, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Locovei, S.; Wang, J.; Dahl, G. Activation of Pannexin 1 Channels by ATP through P2Y Receptors and by Cytoplasmic Calcium. FEBS Lett. 2006, 580, 239–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murali, S.; Zhang, M.; Nurse, C.A. Angiotensin II Mobilizes Intracellular Calcium and Activates Pannexin-1 Channels in Rat Carotid Body Type II Cells via AT1 Receptors. J. Physiol. 2014, 592, 4747–4762. [Google Scholar] [CrossRef] [PubMed]
- Retamal, M.A. Connexin and Pannexin Hemichannels Are Regulated by Redox Potential. Front. Physiol. 2014, 5, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, R.J.; Zhou, N.; MacVicar, B.A. Ischemia Opens Neuronal Gap Junction Hemichannels. Science 2006, 312, 924–927. [Google Scholar] [CrossRef]
- Locovei, S.; Bao, L.; Dahl, G. Pannexin 1 in Erythrocytes: Function without a Gap. Proc. Natl. Acad. Sci. USA 2006, 103, 7655–7659. [Google Scholar] [CrossRef] [Green Version]
- Suadicani, S.O.; Iglesias, R.; Wang, J.; Dahl, G.; Spray, D.C.; Scemes, E. ATP Signaling Is Deficient in Cultured Pannexin1-Null Mouse Astrocytes. Glia 2012, 60, 1106–1116. [Google Scholar] [CrossRef] [Green Version]
- DeLalio, L.J.; Billaud, M.; Ruddiman, C.A.; Johnstone, S.R.; Butcher, J.T.; Wolpe, A.G.; Jin, X.; Keller, T.C.S.; Keller, A.S.; Rivière, T.; et al. Constitutive SRC-Mediated Phosphorylation of Pannexin 1 at Tyrosine 198 Occurs at the Plasma Membrane. J. Biol. Chem. 2019, 294, 6940–6956. [Google Scholar] [CrossRef]
- Weilinger, N.L.; Lohman, A.W.; Rakai, B.D.; Ma, E.M.M.; Bialecki, J.; Maslieieva, V.; Rilea, T.; Bandet, M.V.; Ikuta, N.T.; Scott, L.; et al. Metabotropic NMDA Receptor Signaling Couples Src Family Kinases to Pannexin-1 during Excitotoxicity. Nat. Neurosci. 2016, 19, 432–442. [Google Scholar] [CrossRef]
- Chiu, Y.-H.; Jin, X.; Medina, C.B.; Leonhardt, S.A.; Kiessling, V.; Bennett, B.C.; Shu, S.; Tamm, L.K.; Yeager, M.; Ravichandran, K.S.; et al. A Quantized Mechanism for Activation of Pannexin Channels. Nat. Commun. 2017, 8, 14324. [Google Scholar] [CrossRef] [Green Version]
- Chekeni, F.B.; Elliott, M.R.; Sandilos, J.K.; Walk, S.F.; Kinchen, J.M.; Lazarowski, E.R.; Armstrong, A.J.; Penuela, S.; Laird, D.W.; Salvesen, G.S.; et al. Pannexin 1 Channels Mediate ‘Find-Me’ Signal Release and Membrane Permeability during Apoptosis. Nature 2010, 467, 863–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Dahl, G. Pannexin1: A Multifunction and Multiconductance and/or Permeability Membrane Channel. Am. J. Physiol.-Cell Physiol. 2018, 315, C290–C299. [Google Scholar] [CrossRef]
- Mim, C.; Perkins, G.; Dahl, G. Structure versus Function: Are New Conformations of Pannexin 1 yet to Be Resolved? J. Gen. Physiol. 2021, 153, e202012754. [Google Scholar] [CrossRef]
- Crespo Yanguas, S.; Willebrords, J.; Johnstone, S.R.; Maes, M.; Decrock, E.; De Bock, M.; Leybaert, L.; Cogliati, B.; Vinken, M. Pannexin1 as Mediator of Inflammation and Cell Death. Biochim. Biophys. Acta 2017, 1864, 51. [Google Scholar] [CrossRef]
- Willebrords, J.; Maes, M.; Pereira, I.V.A.; da Silva, T.C.; Govoni, V.M.; Lopes, V.V.; Crespo Yanguas, S.; Shestopalov, V.I.; Nogueira, M.S.; de Castro, I.A.; et al. Protective Effect of Genetic Deletion of Pannexin1 in Experimental Mouse Models of Acute and Chronic Liver Disease. Biochim. Biophys. Acta 2018, 1864, 819. [Google Scholar] [CrossRef]
- Leroy, K.; Vilas-Boas, V.; Gijbels, E.; Vanderborght, B.; Devisscher, L.; Cogliati, B.; Van Den Bossche, B.; Colle, I.; Vinken, M. Expression of Connexins and Pannexins in Diseased Human Liver. EXCLI J. 2022, 21, 1111. [Google Scholar] [CrossRef]
- Burnstock, G.; Knight, G.E. Cellular Distribution and Functions of P2 Receptor Subtypes in Different Systems. In International Review of Cytology; Academic Press: Cambridge, MA, USA, 2004; Volume 240, pp. 31–304. [Google Scholar]
- Surprenant, A.; Rassendren, F.; Kawashima, E.; North, R.A.; Buell, G. The Cytolytic P2Z Receptor for Extracellular ATP Identified as a P 2X Receptor (P2X7). Science 1996, 272, 735–738. [Google Scholar] [CrossRef] [PubMed]
- Klapperstück, M.; Büttner, C.; Schmalzing, G.; Markwardt, F. Functional Evidence of Distinct ATP Activation Sites at the Human P2X 7 Receptor. J. Physiol. 2001, 534, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Donnelly-Roberts, D.L.; Namovic, M.T.; Han, P.; Jarvis, M.F. Mammalian P2X7 Receptor Pharmacology: Comparison of Recombinant Mouse, Rat and Human P2X7 Receptors. Br. J. Pharmacol. 2009, 157, 1203–1214. [Google Scholar] [CrossRef] [Green Version]
- Gribble, F.M.; Loussouarn, G.; Tucker, S.J.; Zhao, C.; Nichols, C.G.; Ashcroft, F.M. A Novel Method for Measurement of Submembrane ATP Concentration. J. Biol. Chem. 2000, 275, 30046–30049. [Google Scholar] [CrossRef] [Green Version]
- Wang, E.C.Y.; Lee, J.-M.; Ruiz, W.G.; Balestreire, E.M.; von Bodungen, M.; Barrick, S.; Cockayne, D.A.; Birder, L.A.; Apodaca, G. ATP and Purinergic Receptor–Dependent Membrane Traffic in Bladder Umbrella Cells. J. Clin. Investig. 2005, 115, 2412–2422. [Google Scholar] [CrossRef] [Green Version]
- Adinolfi, E.; Giuliani, A.L.; De Marchi, E.; Pegoraro, A.; Orioli, E.; Di Virgilio, F. The P2X7 Receptor: A Main Player in Inflammation. Biochem. Pharmacol. 2018, 151, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Di Virgilio, F.; Sarti, A.C.; Falzoni, S.; De Marchi, E.; Adinolfi, E. Extracellular ATP and P2 Purinergic Signalling in the Tumour Microenvironment. Nat. Rev. Cancer 2018, 18, 601–618. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.; Silva, R.; Coutinho-Silva, R.; Takiya, C.; Battastini, A.; Morrone, F.; Campos, M. The Role of P2X7 Purinergic Receptors in Inflammatory and Nociceptive Changes Accompanying Cyclophosphamide-Induced Haemorrhagic Cystitis in Mice. Br. J. Pharmacol. 2012, 165, 183–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taidi, Z.; Zhou, T.; Moore, K.H.; Mansfield, K.J.; Liu, L. P2X7 Receptor Blockade Protects Against Acrolein-Induced Bladder Damage: A Potential New Therapeutic Approach for the Treatment of Bladder Inflammatory Diseases. Front. Pharmacol. 2021, 12, 682520. [Google Scholar] [CrossRef]
- Pellegatti, P.; Falzoni, S.; Pinton, P.; Rizzuto, R.; Di Virgilio, F. A Novel Recombinant Plasma Membrane-Targeted Luciferase Reveals a New Pathway for ATP Secretion. Mol. Biol. Cell 2005, 16, 3659–3665. [Google Scholar] [CrossRef] [Green Version]
- Peverini, L.; Beudez, J.; Dunning, K.; Chataigneau, T.; Grutter, T. New Insights Into Permeation of Large Cations Through ATP-Gated P2X Receptors. Front. Mol. Neurosci. 2018, 11, 265. [Google Scholar] [CrossRef] [Green Version]
- Di Virgilio, F.; Schmalzing, G.; Markwardt, F. The Elusive P2X7 Macropore. Trends Cell Biol. 2018, 28, 392–404. [Google Scholar] [CrossRef]
- Ma, W.; Hui, H.; Pelegrin, P.; Surprenant, A. Pharmacological Characterization of Pannexin-1 Currents Expressed in Mammalian Cells. J. Pharmacol. Exp. Ther. 2009, 328, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Qiu, F.; Dahl, G. A Permeant Regulating Its Permeation Pore: Inhibition of Pannexin 1 Channels by ATP. Am. J. Physiol.-Cell Physiol. 2009, 296, C250–C255. [Google Scholar] [CrossRef]
- Boyce, A.K.J.; Kim, M.S.; Wicki-Stordeur, L.E.; Swayne, L.A. ATP Stimulates Pannexin 1 Internalization to Endosomal Compartments. Biochem. J. 2015, 470, 319–330. [Google Scholar] [CrossRef]
- Purohit, R.; Bera, A.K. Pannexin 1 Plays a Pro-Survival Role by Attenuating P2X7 Receptor-Mediated Ca2+ Influx. Cell Calcium 2021, 99, 102458. [Google Scholar] [CrossRef]
- Allsopp, R.C.; Dayl, S.; Schmid, R.; Evans, R.J. Unique Residues in the ATP Gated Human P2X7 Receptor Define a Novel Allosteric Binding Pocket for the Selective Antagonist AZ10606120. Sci. Rep. 2017, 7, 725. [Google Scholar] [CrossRef] [Green Version]
- Fowler, C.J.; Griffiths, D.; de Groat, W.C. The Neural Control of Micturition. Nat. Rev. Neurosci. 2008, 9, 453–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aresta Branco, M.S.L.; Gutierrez Cruz, A.; Borhani Peikani, M.; Mutafova-Yambolieva, V.N. Sensory Neurons, PIEZO Channels and PAC1 Receptors Regulate the Mechanosensitive Release of Soluble Ectonucleotidases in the Murine Urinary Bladder Lamina Propria. Int. J. Mol. Sci. 2023, 24, 7322. [Google Scholar] [CrossRef]
- Gutierrez Cruz, A.; Aresta Branco, M.S.L.; Perrino, B.A.; Sanders, K.M.; Mutafova-Yambolieva, V.N. Urinary ATP Levels Are Controlled by Nucleotidases Released from the Urothelium in a Regulated Manner. Metabolites 2022, 13, 30. [Google Scholar] [CrossRef]
- Negoro, H.; Lutz, S.E.; Liou, L.S.; Kanematsu, A.; Ogawa, O.; Scemes, E.; Suadicani, S.O. Pannexin 1 Involvement in Bladder Dysfunction in a Multiple Sclerosis Model. Sci. Rep. 2013, 3, 2152. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.W.G.; Sui, G.; Wu, R.; Rong, W.; Wildman, S.; Montgomery, B.; Ali, A.; Langley, S.; Ruggieri, M.R.; Wu, C. TRPV4 Receptor as a Functional Sensory Molecule in Bladder Urothelium: Stretch-independent, Tissue-specific Actions and Pathological Implications. FASEB J. 2020, 34, 263–286. [Google Scholar] [CrossRef] [Green Version]
- Diem, K.; Fauler, M.; Fois, G.; Hellmann, A.; Winokurow, N.; Schumacher, S.; Kranz, C.; Frick, M. Mechanical Stretch Activates Piezo1 in Caveolae of Alveolar Type I Cells to Trigger ATP Release and Paracrine Stimulation of Surfactant Secretion from Alveolar Type II Cells. FASEB J. 2020, 34, 12785–12804. [Google Scholar] [CrossRef]
- Vial, C.; Evans, R.J. P2X Receptor Expression in Mouse Urinary Bladder and the Requirement of P2X 1 Receptors for Functional P2X Receptor Responses in the Mouse Urinary Bladder Smooth Muscle. Br. J. Pharmacol. 2000, 131, 1489–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svennersten, K.; Hallén-Grufman, K.; de Verdier, P.J.; Wiklund, N.P.; Poljakovic, M. Localization of P2X Receptor Subtypes 2, 3 and 7 in Human Urinary Bladder. BMC Urol. 2015, 15, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenertz, L.Y.; Gavala, M.L.; Zhu, Y.; Bertics, P.J. Transcriptional Control Mechanisms Associated with the Nucleotide Receptor P2X7, a Critical Regulator of Immunologic, Osteogenic, and Neurologic Functions. Immunol. Res. 2011, 50, 22–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Chai, T.C. Augmented Extracellular ATP Signaling in Bladder Urothelial Cells from Patients with Interstitial Cystitis. Am. J. Physiol.-Cell Physiol. 2006, 290, C27–C34. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Chapple, C.R.; Surprenant, A.M.; Chess-Williams, R. Enhanced Adenosine Triphosphate Release From the Urothelium of Patients With Painful Bladder Syndrome: A Possible Pathophysiological Explanation. J. Urol. 2007, 178, 1533–1536. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Chapple, C.R.; Rosario, D.; Tophill, P.R.; Chess-Williams, R. In Vitro Release of Adenosine Triphosphate from the Urothelium of Human Bladders with Detrusor Overactivity, Both Neurogenic and Idiopathic. Eur. Urol. 2010, 57, 1087–1092. [Google Scholar] [CrossRef]
- Sun, Y.; MaLossi, J.; Jacobs, S.C.; Chai, T.C. Effect of Doxazosin on Stretch-Activated Adenosine Triphosphate Release in Bladder Urothelial Cells from Patients with Benign Prostatic Hyperplasia. Urology 2002, 60, 351–356. [Google Scholar] [CrossRef]
- Birder, L.A.; Barrick, S.R.; Roppolo, J.R.; Kanai, A.J.; de Groat, W.C.; Kiss, S.; Buffington, C.A. Feline Interstitial Cystitis Results in Mechanical Hypersensitivity and Altered ATP Release from Bladder Urothelium. Am. J. Physiol.-Ren. Physiol. 2003, 285, F423–F429. [Google Scholar] [CrossRef] [Green Version]
- Munoz, A.; Smith, C.P.; Boone, T.B.; Somogyi, G.T. Overactive and Underactive Bladder Dysfunction Is Reflected by Alterations in Urothelial ATP and NO Release. Neurochem. Int. 2011, 58, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Salas, N.A.; Somogyi, G.T.; Gangitano, D.A.; Boone, T.B.; Smith, C.P. Receptor Activated Bladder and Spinal ATP Release in Neurally Intact and Chronic Spinal Cord Injured Rats. Neurochem. Int. 2007, 50, 345–350. [Google Scholar] [CrossRef] [Green Version]
- Khera, M.; Somogyi, G.T.; Kiss, S.; Boone, T.B.; Smith, C.P. Botulinum Toxin A Inhibits ATP Release from Bladder Urothelium after Chronic Spinal Cord Injury. Neurochem. Int. 2004, 45, 987–993. [Google Scholar] [CrossRef]
- Smith, C.P.; Vemulakonda, V.M.; Kiss, S.; Boone, T.B.; Somogyi, G.T. Enhanced ATP Release from Rat Bladder Urothelium during Chronic Bladder Inflammation: Effect of Botulinum Toxin A. Neurochem. Int. 2005, 47, 291–297. [Google Scholar] [CrossRef]
- MacKenzie, A.; Wilson, H.L.; Kiss-Toth, E.; Dower, S.K.; North, R.A.; Surprenant, A. Rapid Secretion of Interleukin-1β by Microvesicle Shedding. Immunity 2001, 15, 825–835. [Google Scholar] [CrossRef] [PubMed]
- Virginio, C.; MacKenzie, A.; North, R.A.; Surprenant, A. Kinetics of Cell Lysis, Dye Uptake and Permeability Changes in Cells Expressing the Rat P2X 7 Receptor. J. Physiol. 1999, 519, 335–346. [Google Scholar] [CrossRef]
- Hanley, P.J.; Kronlage, M.; Kirschning, C.; del Rey, A.; Di Virgilio, F.; Leipziger, J.; Chessell, I.P.; Sargin, S.; Filippov, M.A.; Lindemann, O.; et al. Transient P2X7 Receptor Activation Triggers Macrophage Death Independent of Toll-like Receptors 2 and 4, Caspase-1, and Pannexin-1 Proteins. J. Biol. Chem. 2012, 287, 10650–10663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banz, Y.; Beldi, G.; Wu, Y.; Atkinson, B.; Usheva, A.; Robson, S.C. CD39 Is Incorporated into Plasma Microparticles Where It Maintains Functional Properties and Impacts Endothelial Activation. Br. J. Haematol. 2008, 142, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Clayton, A.; Al-Taei, S.; Webber, J.; Mason, M.D.; Tabi, Z. Cancer Exosomes Express CD39 and CD73, Which Suppress T Cells through Adenosine Production. J. Immunol. 2011, 187, 676–683. [Google Scholar] [CrossRef] [Green Version]
- Trams, E.G.; Lauter, C.J.; Salem, N., Jr.; Heine, U. Exfoliation of Membrane Ecto-Enzymes in the Form of Micro-Vesicles. Biochim. Et Biophys. Acta (BBA)-Biomembr. 1981, 645, 63–70. [Google Scholar] [CrossRef]
- Pelegrin, P.; Surprenant, A. The P2X7 Receptor–Pannexin Connection to Dye Uptake and IL-1β Release. Purinergic Signal. 2009, 5, 129–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, W.-J.; Illes, P. Involvement of P2X7 Receptors in Chronic Pain Disorders. Purinergic Signal. 2022, 18, 83–92. [Google Scholar] [CrossRef]
- Ren, W.; Rubini, P.; Tang, Y.; Engel, T.; Illes, P. Inherent P2X7 Receptors Regulate Macrophage Functions during Inflammatory Diseases. Int. J. Mol. Sci. 2021, 23, 232. [Google Scholar] [CrossRef]
- Atkinson, L.; Batten, T.F.C.; Moores, T.S.; Varoqui, H.; Erickson, J.D.; Deuchars, J. Differential Co-Localisation of the P2X7 Receptor Subunit with Vesicular Glutamate Transporters VGLUT1 and VGLUT2 in Rat CNS. Neuroscience 2004, 123, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Deuchars, S.A.; Atkinson, L.; Brooke, R.E.; Musa, H.; Milligan, C.J.; Batten, T.F.C.; Buckley, N.J.; Parson, S.H.; Deuchars, J. Neuronal P2X 7 Receptors Are Targeted to Presynaptic Terminals in the Central and Peripheral Nervous Systems. J. Neurosci. 2001, 21, 7143–7152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durnin, L.; Corrigan, R.D.; Sanders, K.M.; Mutafova-Yambolieva, V.N. A Decentralized (Ex Vivo) Murine Bladder Model with the Detrusor Muscle Removed for Direct Access to the Suburothelium during Bladder Filling. J. Vis. Exp. 2019, 2019, e60344. [Google Scholar] [CrossRef]
- McCarthy, A.E.; Yoshioka, C.; Mansoor, S.E. Full-Length P2X7 Structures Reveal How Palmitoylation Prevents Channel Desensitization. Cell 2019, 179, 659–670. [Google Scholar] [CrossRef]
- Levitt, B.; Head, R.J.; Westfall, D.P. High-Pressure Liquid Chromatographic-Fluorometric Detection of Adenosine and Adenine Nucleotides: Application to Endogenous Content and Electrically Induced Release of Adenyl Purines in Guinea Pig Vas Deferens. Anal. Biochem. 1984, 137, 93–100. [Google Scholar] [CrossRef]
- Bobalova, J.; Bobal, P.; Mutafova-Yambolieva, V.N. High-Performance Liquid Chromatographic Technique for Detection of a Fluorescent Analogue of ADP-Ribose in Isolated Blood Vessel Preparations. Anal. Biochem. 2002, 305, 269–276. [Google Scholar] [CrossRef]
- Durnin, L.; Hayoz, S.; Corrigan, R.D.; Yanez, A.; Koh, S.D.; Mutafova-Yambolieva, V.N. Urothelial Purine Release during Filling of Murine and Primate Bladders. Am. J. Physiol.-Ren. Physiol. 2016, 311, F708–F716. [Google Scholar] [CrossRef] [Green Version]
- Vollert, J.; Macleod, M.; Dirnagl, U.; Kas, M.J.; Michel, M.C.; Potschka, H.; Riedel, G.; Wever, K.E.; Würbel, H.; Steckler, T.; et al. The EQIPD Framework for Rigor in the Design, Conduct, Analysis and Documentation of Animal Experiments. Nat. Methods 2022, 19, 1334–1337. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aresta Branco, M.S.L.; Gutierrez Cruz, A.; Peri, L.E.; Mutafova-Yambolieva, V.N. The Pannexin 1 Channel and the P2X7 Receptor Are in Complex Interplay to Regulate the Release of Soluble Ectonucleotidases in the Murine Bladder Lamina Propria. Int. J. Mol. Sci. 2023, 24, 9964. https://doi.org/10.3390/ijms24129964
Aresta Branco MSL, Gutierrez Cruz A, Peri LE, Mutafova-Yambolieva VN. The Pannexin 1 Channel and the P2X7 Receptor Are in Complex Interplay to Regulate the Release of Soluble Ectonucleotidases in the Murine Bladder Lamina Propria. International Journal of Molecular Sciences. 2023; 24(12):9964. https://doi.org/10.3390/ijms24129964
Chicago/Turabian StyleAresta Branco, Mafalda S. L., Alejandro Gutierrez Cruz, Lauren E. Peri, and Violeta N. Mutafova-Yambolieva. 2023. "The Pannexin 1 Channel and the P2X7 Receptor Are in Complex Interplay to Regulate the Release of Soluble Ectonucleotidases in the Murine Bladder Lamina Propria" International Journal of Molecular Sciences 24, no. 12: 9964. https://doi.org/10.3390/ijms24129964