
Oligo-FISH of Populus simonii Pachytene Chromosomes Improves Karyotyping and Genome Assembly
Abstract
:1. Introduction
2. Results
2.1. Karyotyping of Metaphase Chromosomes
2.2. Comparing the Three Methods for Pachytene Chromosomes Preparation
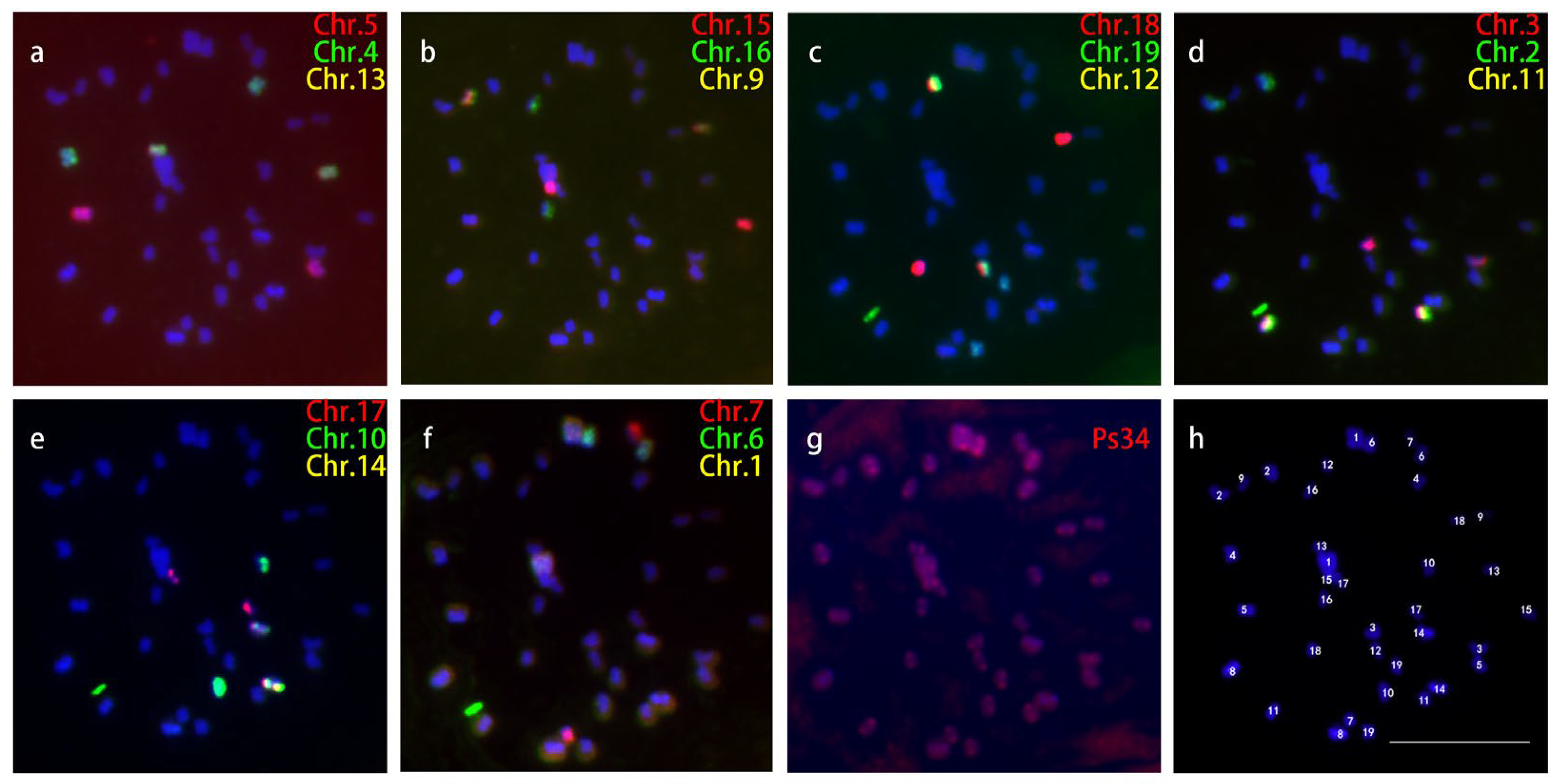
2.3. Karyotyping of Pachytene Chromosomes
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Mitotic Metaphase Chromosome Preparation
4.3. Meiotic Pachytene Chromosome Preparation
4.4. Probes
4.5. Fluorescence In Situ Hybridization
4.6. Cytological Measurements and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Xin, H.; Zhang, T.; Wu, Y.; Zhang, W.; Zhang, P.; Xi, M.; Jiang, J. An extraordinarily stable karyotype of the woody Populus species revealed by chromosome painting. Plant J. 2020, 101, 253–264. [Google Scholar] [CrossRef]
- Koo, D.; Jiang, J. Super-stretched pachytene chromosomes for fluorescence in situ hybridization mapping and immunodetection of DNA methylation. Plant J. 2009, 59, 509–516. [Google Scholar] [CrossRef]
- Cheng, Z.; Buell, C.R.; Wing, R.A.; Gu, M.; Jiang, J. Toward a Cytological Characterization of the Rice Genome. Genome Res. 2001, 11, 2133–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Zhang, T.; Thammapichai, P.; Weng, Y.; Jiang, J. Chromosome-specific painting in Cucumis species using bulked oligonucleotides. Genetics 2015, 200, 771–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braz, G.T.; He, L.; Zhao, H.; Rouillard, J.; Zhang, T.; Semrau, K.; Torres, G.A.; Jiang, J. Comparative oligo-FISH mapping: An efficient and powerful methodology to reveal karyotypic and chromosomal evolution. Genetics 2018, 208, 513–523. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Gill, B.S. Current status and the future of fluorescence in situ hybridization (FISH) in plant genome research. Genome 2006, 49, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Albert, P.S.; Zhang, T.; Semrau, K.; Rouillard, J.; Kao, Y.; Wang, C.R.; Danilova, T.V.; Jiang, J.; Birchler, J.A. Whole-chromosome paints in maize reveal rearrangements, nuclear domains, and chromosomal relationships. Proc. Natl. Acad. Sci. USA 2019, 116, 1679–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suna, J.; Sua, D.; Ma, R.; Chena, L.; Caob, Q.; Lia, Z.; Hana, Y. Chromosome painting reveals the genomic structure of three polyploid species of Ipomoea. Genome 2022, 65, 331–339. [Google Scholar] [CrossRef]
- Xin, H.; Zhang, T.; Han, Y.; Wu, Y.; Shi, J.; Xi, M.; Jiang, J. Chromosome painting and comparative physical mapping of the sex chromosomes in Populus tomentosa and Populus deltoides. Chromosoma 2018, 127, 313–321. [Google Scholar] [CrossRef]
- Zhu, J.; Tian, J.; Wang, J.; Nie, S. Variation of traits on seeds and germination derived from the hybridization between the sections Tacamahaca and Aigeiros of the genus Populus. Forests 2018, 9, 516. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Yao, D.; Chen, Y.; Yang, W.; Zhao, W.; Gao, H.; Tong, C. De novo genome assembly of Populus simonii further supports that Populus simonii and Populus trichocarpa belong to different sections. G3 2020, 10, 455–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, Y.; Xin, H.; Zhao, Y.; Sun, Z.; Dai, X.; Xie, Z.; Xi, M. Study on meiosis of microspore mother cells and germination rate of pollen grains in Populus simonii Carr. J. Nanjing For. Univ. Nat. Sci. Ed. 2017, 41, 177–180. [Google Scholar]
- Muakrong, N.; Kikuchi, S.; Fukuhara, S.; Tanya, P.; Srinives, P. Two jatropha karyotypes constructed from meiotic pachytene chromosomes: Pericentric distribution of heterochromatin and variation in repetitive DNAs. PLoS ONE 2018, 13, e0208549. [Google Scholar] [CrossRef] [PubMed]
- Menke, M.; Fuchs, J.; Schubert, I. A comparison of sequence resolution on plant chromosomes PRINS versus FISH. Theor. Appl. Genet. 1998, 97, 1314–1320. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, S.; Li, X.; Han, S.; Song, W.; Qi, L. A comparative study among sections on the karyotypes of Populus. Guihaia 2005, 4, 338–340. [Google Scholar]
- Li, M.; Chen, R. A suggestion on the standardization of karyotype analysis in plants. J. Wuhan Bot. Res. 1985, 3, 297–302. [Google Scholar]
- Li, S.; Guo, Y.; Li, J.; Zhang, D.; Wang, B.; Li, N.; Deng, C.; Gao, W. The landscape of transposable elements and satellite DNAs in the genome of a dioecious plant spinach (Spinacia oleracea L.). Mob. DNA 2019, 10, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Xin, H.; Lan, Y.; Shi, J.; Ma, Y.; Xi, M. Pachytene chromosome preparation in Populus deltoides marsh. Curr. Protoc. Plant Biol. 2016, 1, 566–573. [Google Scholar] [CrossRef]
- Zhang, W.; Lee, H.-R.; Koo, D.-H.; Jiang, J. Epigenetic modification of centromeric chromatin: Hypomethylation of DNA sequences in the CENH3-associated chromatin in Arabidopsis thaliana and maize. Plant Cell 2008, 20, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Liang, C. Assembly of chromosome-scale contigs by efficiently resolving repetitive sequences with long reads. Nat. Commun. 2019, 10, 5360–5369. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Yang, X.; Jia, Y.; Xu, Y.; Jia, P.; Dang, N.; Wang, S.; Xu, T.; Zhao, X.; Gao, S.; et al. High-quality Arabidopsis thaliana genome assembly with Nanopore and HiFi long reads. Genomics Proteom. Bioinform. 2022, 20, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Zhao, Q.; Yan, W.; Li, M.; Liu, Y.; Cheng, C.; Zhang, L.; Yu, X.; Li, J.; Qian, C.; et al. Flexible chromosome painting based on multiplex PCR of oligonucleotides and its application for comparative chromosome analyses in Cucumis. Plant J. 2020, 102, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Torres, G.A.; Novák, P.; Macas, J.; Gong, Z.; Iovene, M.; Hirsch, C.D.; Buel, C.R.; Jiang, J. Organization and evolution of subtelomeric satellite repeats in the potato genome. G3 2011, 1, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Lou, Q.; Meng, Y.; Wang, P.; Qin, X.; Cheng, C.; Zhou, J.; Yu, X.; Jahn, M.; Chen, J. Reconstruction of ancestral karyotype illuminates chromosome evolution in the genus Cucumis. Plant J. 2021, 107, 1243–1259. [Google Scholar] [CrossRef]
- Comai, L.; Amundson, K.R.; Ordonez, B.; Zhao, X.; Braz, G.T.; Jiang, J.; Henry, I.M. LD-CNV: Rapid and simple discovery of chromosomal translocations using linkage disequilibrium between copy number variable loci. Genetics 2021, 219, iyab137. [Google Scholar] [CrossRef]
- Pakull, B.; Groppe, K.; Meyer, M.; Markussen, T.; Fladung, M. Genetic linkage mapping in aspen (Populus tremula L. and Populus tremuloides Michx.). Tree Genet. Genomes 2009, 5, 505–515. [Google Scholar] [CrossRef]
- Paolucci, I.; Gaudet, M.; Jorge, V.; Beritognolo, I.; Terzoli, S.; Kuzminsky, E.; Muleo, R.; Mugnozza, G.S.; Sabatti, M. Genetic linkage maps of Populus alba L. and comparative mapping analysis of sex determination across Populus species. Tree Genet. Genomes 2010, 6, 863–875. [Google Scholar] [CrossRef]
- Gaudet, M.; Jorge, V.; Paolucci, I.; Beritognolo, I.; Mugnozza, G.S.; Sabatti, M. Genetic linkage maps of Populus nigra L. including AFLPs, SSRs, SNPs, and sex trait. Tree Genet. Genomes 2007, 4, 25–36. [Google Scholar] [CrossRef]
- Yin, T.; DiFazio, S.P.; Gunter, L.E.; Zhang, X.; Sewell, M.M.; Woolbright, S.A.; Allan, G.J.; Kelleher, C.T.; Douglas, C.J.; Scott, A.W.; et al. Genome structure and emerging evidence of an incipient sex chromosome in Populus. Genome Res. 2008, 18, 422–430. [Google Scholar] [CrossRef]
- Kersten, B.; Pakull, B.; Fladung, M. Genomics of sex determination in dioecious trees and woody plants. Trees 2017, 31, 1113–1125. [Google Scholar] [CrossRef]
- Geraldes, A.; Hefer, C.A.; Capron, A.; Kolosova, N.; Martinez-Nunez, F.; Soolanayakanahally, R.Y.; Stanton, B.; Guy, R.D.; Mansfield, S.D.; Douglas, C.J.; et al. Recent Y chromosome divergence despite ancient origin of dioecy in poplars (Populus). Mol. Ecol. 2015, 24, 3243–3256. [Google Scholar] [CrossRef] [PubMed]
- Kersten, B.; Pakull, B.; Groppe, K.; Lueneburg, J.; Fladung, M. The sex-linked region in Populus tremuloides Turesson 141 corresponds to a pericentromeric region of about two million base pairs on P. trichocarpa chromosome 19. Plant Biol. 2014, 16, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Pakull, B.; Groppe, K.; Mecucci, F.; Gaudet, M.; Sabatti, M.; Fladung, M. Genetic mapping of linkage group XIX and identification of sex-linked SSR markers in a Populus tremula × Populus tremuloides cross. Can. J. For. Res. 2011, 41, 245–253. [Google Scholar] [CrossRef]
- Andras, S.C.; Hartman, T.P.; Marshall, J.A.; Marchant, R.; Power, J.B.; Cocking, E.C.; Davey, M.R. A drop-spreading technique to produce cytoplasm-free mitotic preparations from plants with small chromosomes. Chromosome Res. 1999, 7, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Tuskan, G.A.; Difazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chr. No. | Long Arm (µm) | Short Arm (µm) | Arm Ratio c | |||
|---|---|---|---|---|---|---|
| Metaphase a | Pachytene b | Metaphase a | Pachytene b | Metaphase | Pachytene | |
| 1 | 0.76 ± 0.11 | 31.46 ± 9.21 | 0.52 ± 0.10 | 21.46 ± 6.62 | 1.46 ± 0.17 | 1.48 ± 0.11 |
| 2 | 0.53 ± 0.08 | 23.69 ± 4.45 | 0.37 ± 0.04 | 6.42 ± 1.17 | 1.46 ± 0.16 | 3.69 ± 0.18 |
| 3 | 0.56 ± 0.09 | 9.66 ± 0.69 | 0.30 ± 0.03 | 3.01 ± 0.31 | 1.90 ± 0.31 | 3.23 ± 0.28 |
| 4 | 0.47 ± 0.02 | 10.02 ± 2.260 | 0.33 ± 0.03 | 8.40 ± 1.67 | 1.43 ± 0.16 | 1.19 ± 0.07 |
| 5 | 0.48 ± 0.05 | 7.62 ± 0.69 | 0.36 ± 0.04 | 5.83 ± 0.45 | 1.35 ± 0.14 | 1.31 ± 0.09 |
| 6 | 0.48 ± 0.05 | 15.85 ± 2.80 | 0.37 ± 0.05 | 12.27 ± 2.13 | 1.33 ± 0.19 | 1.29 ± 0.07 |
| 7 | 0.39 ± 0.05 | 4.95 ± 0.44 | 0.30 ± 0.04 | 4.03 ± 0.38 | 1.28 ± 0.09 | 1.23 ± 0.08 |
| 8 d | 0.55 ± 0.07 | 16.37 ± 2.33 | 0.24 ± 0.04 | 3.33 ± 0.49 | 2.34 ± 0.27 | 4.93 ± 0.32 |
| 9 | 0.54 ± 0.05 | 9.05 ± 0.94 | 0.18 ± 0.03 | 0.52 ± 0.12 | 3.09 ± 0.59 | 18.21 ± 5.04 |
| 10 | 0.58 ± 0.09 | 19.53 ± 5.86 | 0.32 ± 0.06 | 3.83 ± 0.81 | 1.85 ± 0.33 | 5.06 ± 0.87 |
| 11 | 0.53 ± 0.09 | 5.49 ± 0.70 | 0.37 ± 0.04 | 5.00 ± 0.61 | 1.48 ± 0.20 | 1.10 ± 0.04 |
| 12 | 0.41 ± 0.04 | 4.38 ± 0.50 | 0.31 ± 0.04 | 3.61 ± 0.38 | 1.31 ± 0.14 | 1.21 ± 0.08 |
| 13 | 0.47 ± 0.03 | 5.06 ± 0.49 | 0.34 ± 0.06 | 3.60 ± 0.42 | 1.41 ± 0.26 | 1.41 ± 0.11 |
| 14 d | 0.63 ± 0.07 | 15.92 ± 2.00 | 0.23 ± 0.04 | 1.14 ± 0.29 | 2.75 ± 0.45 | 14.81 ± 3.85 |
| 15 | 0.46 ± 0.03 | 6.57 ± 0.96 | 0.30 ± 0.04 | 4.35 ± 0.67 | 1.54 ± 0.24 | 1.52 ± 0.08 |
| 16 | 0.49 ± 0.06 | 8.00 ± 1.51 | 0.29 ± 0.05 | 4.70 ± 0.91 | 1.74 ± 0.31 | 1.70 ± 0.06 |
| 17 | 0.44 ± 0.07 | 4.70 ± 0.39 | 0.31 ± 0.03 | 3.28 ± 0.311 | 1.39 ± 0.20 | 1.43 ± 0.06 |
| 18 | 0.46 ± 0.04 | 7.76 ± 1.16 | 0.34 ± 0.03 | 5.24 ± 0.74 | 1.39 ± 0.18 | 1.48 ± 0.09 |
| 19 | 0.42 ± 0.06 | 5.06 ± 0.62 | 0.35 ± 0.05 | 4.27 ± 0.51 | 1.22 ± 0.14 | 1.19 ± 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Liu, G.; Wang, Z.; Ning, Y.; Ni, R.; Xi, M. Oligo-FISH of Populus simonii Pachytene Chromosomes Improves Karyotyping and Genome Assembly. Int. J. Mol. Sci. 2023, 24, 9950. https://doi.org/10.3390/ijms24129950
Zhao Y, Liu G, Wang Z, Ning Y, Ni R, Xi M. Oligo-FISH of Populus simonii Pachytene Chromosomes Improves Karyotyping and Genome Assembly. International Journal of Molecular Sciences. 2023; 24(12):9950. https://doi.org/10.3390/ijms24129950
Chicago/Turabian StyleZhao, Yilian, Guangxin Liu, Ziyue Wang, Yihang Ning, Runxin Ni, and Mengli Xi. 2023. "Oligo-FISH of Populus simonii Pachytene Chromosomes Improves Karyotyping and Genome Assembly" International Journal of Molecular Sciences 24, no. 12: 9950. https://doi.org/10.3390/ijms24129950