
Dimethyl Fumarate and Intestine: From Main Suspect to Potential Ally against Gut Disorders
 and
and
Abstract
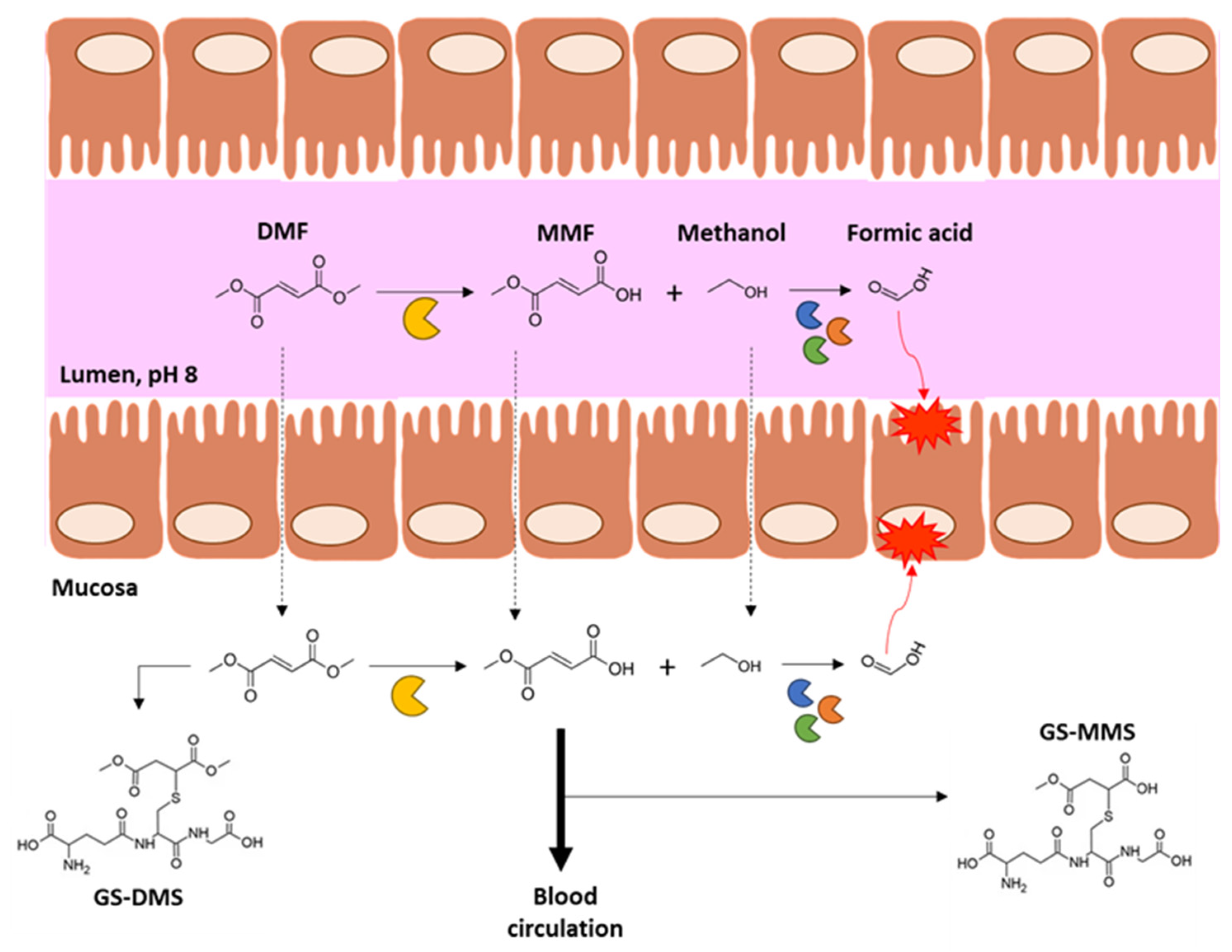
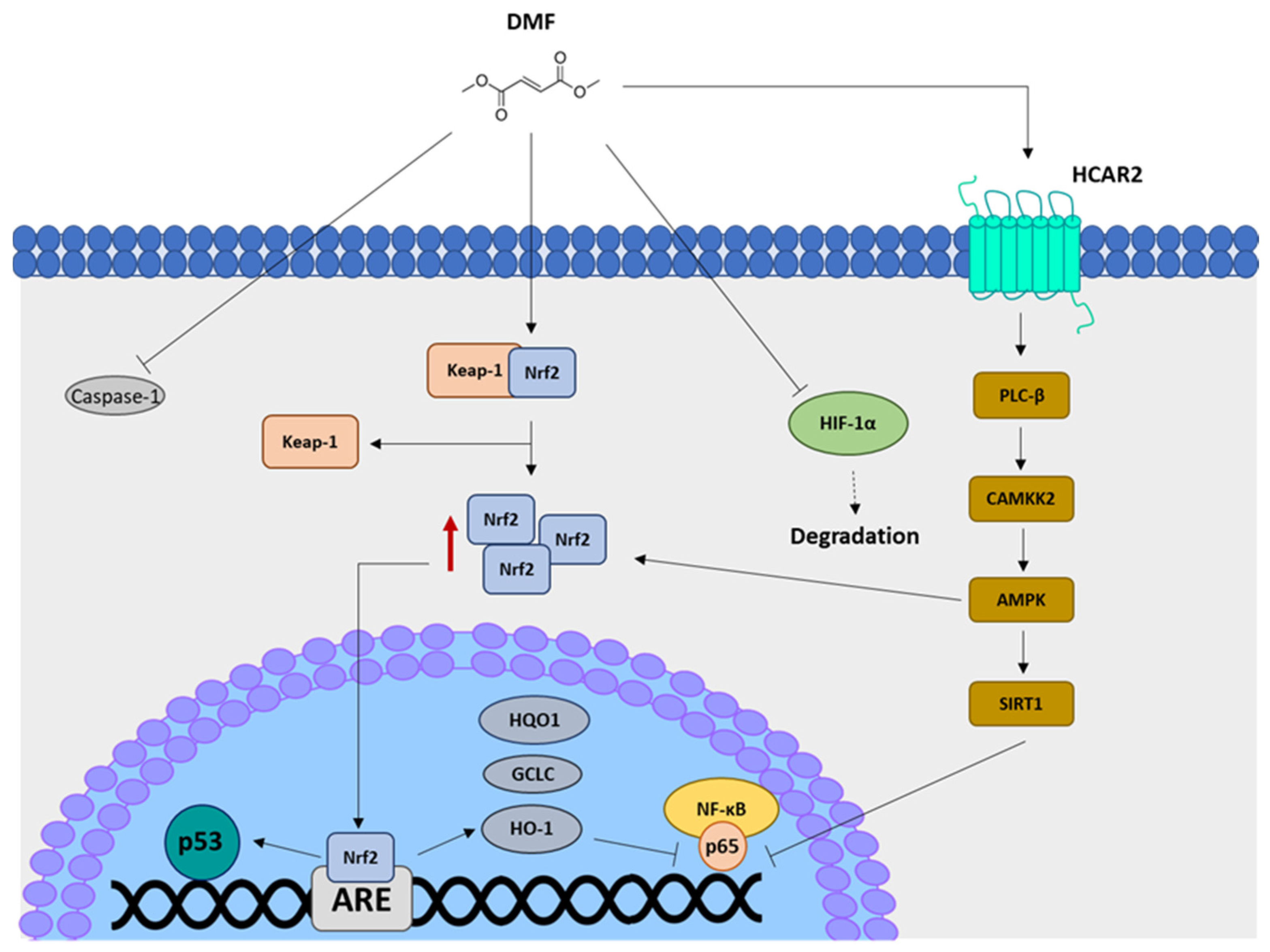
:1. Dimethyl Fumarate Pharmacokinetics and Mechanisms of Action
2. Dimethyl Fumarate and Intestine: State of the Art
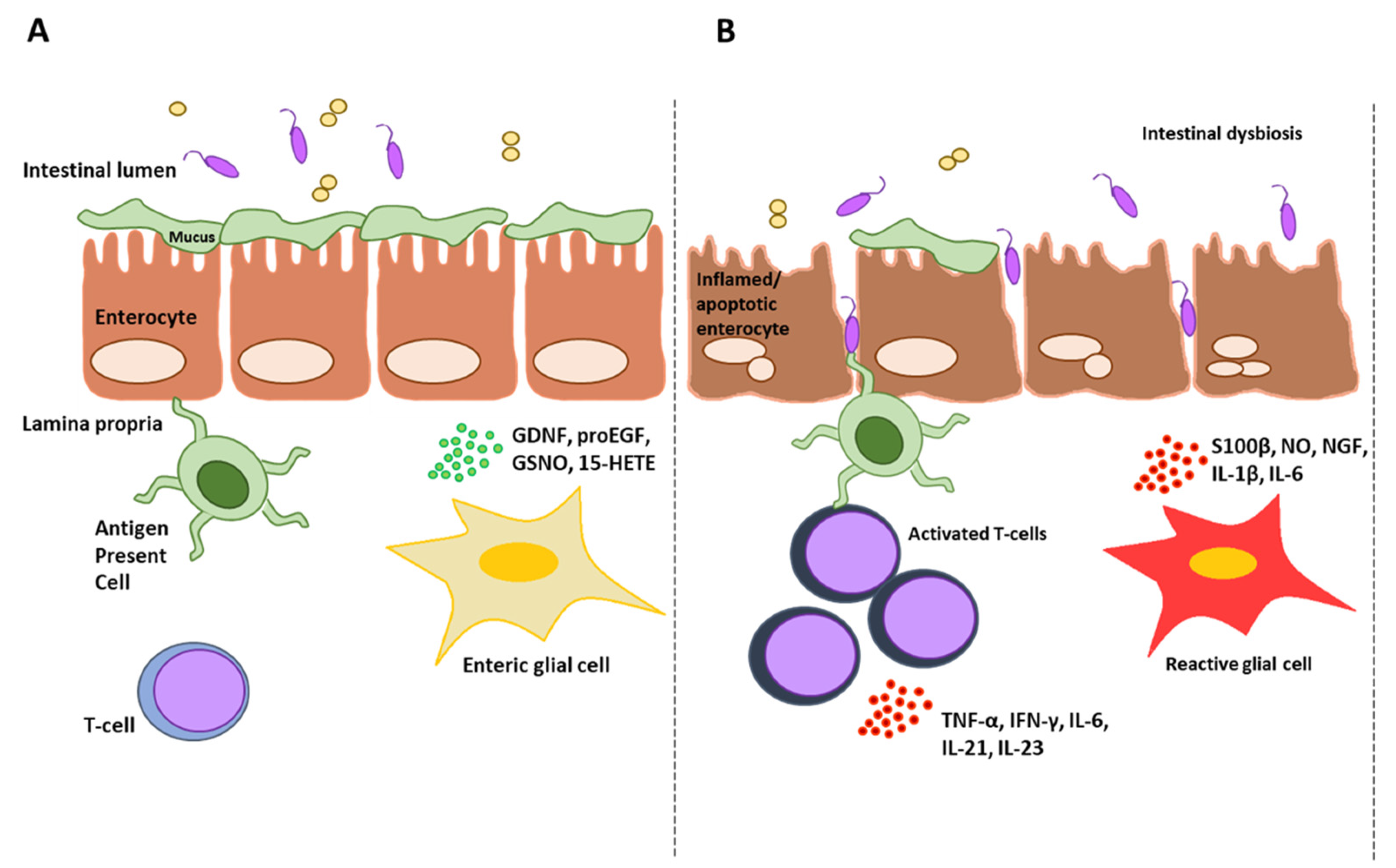
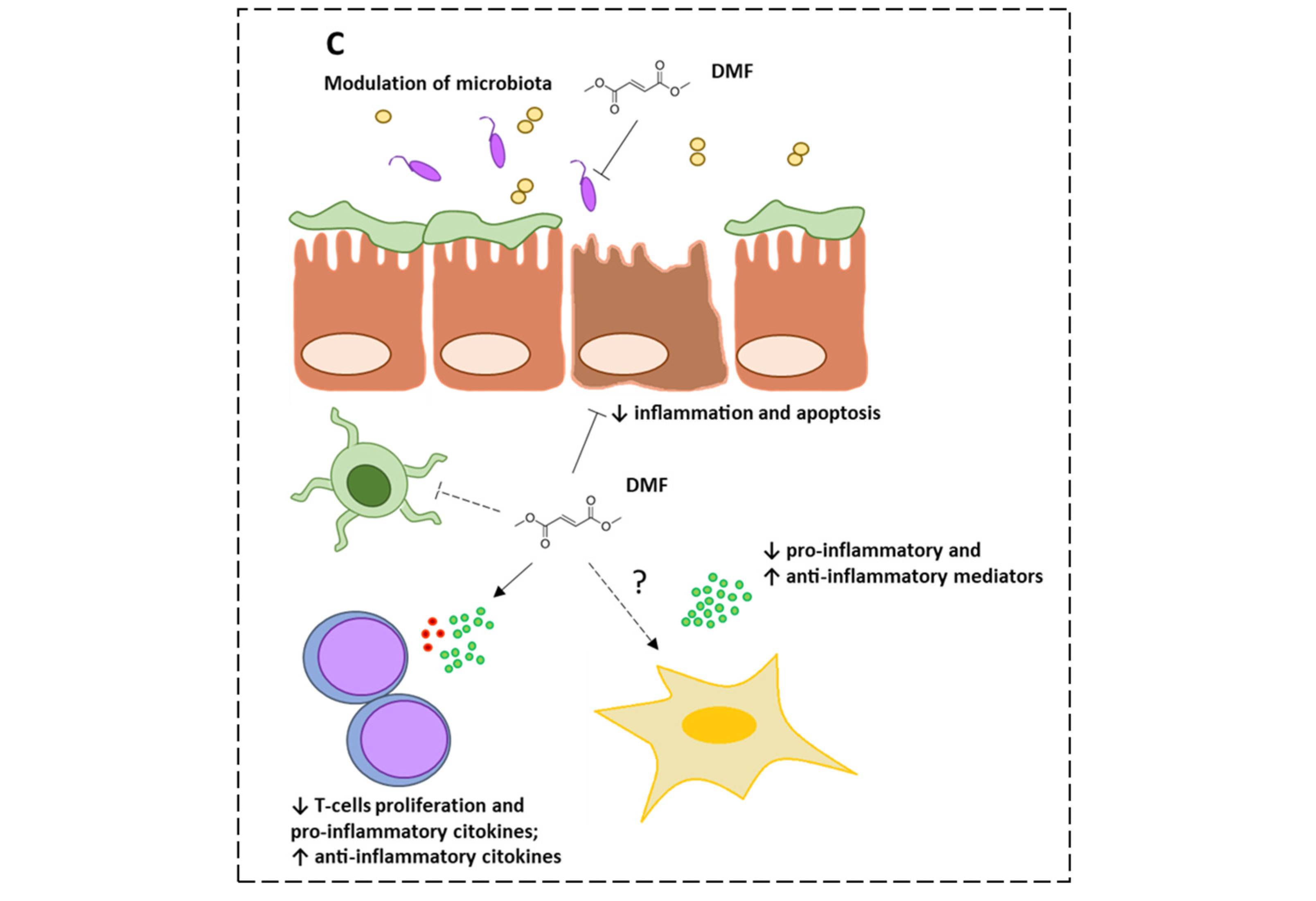
2.1. Dimethyl Fumarate’s Effects on Intestinal Mucosa
2.2. Dimethyl Fumarate’s Effects on Gut Microbiota
3. Intestinal Disorders as Targets of DMF
3.1. Inflammatory Bowel Diseases (IBDs)
3.2. Celiac Disease
4. Rationale for DMF Repurposing in Intestinal Pathologies
5. Other Considerations for DMF-Based Approaches in Intestinal Disorders
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saidu, N.E.B.; Kavian, N.; Leroy, K.; Jacob, C.; Nicco, C.; Batteux, F.; Alexandre, J. Dimethyl fumarate, a two-edged drug: Current status and future directions. Med. Res. Rev. 2019, 39, 1923–1952. [Google Scholar] [CrossRef] [PubMed]
- Eleni, K.; Michalis, S.C.; Patrickios, C.S. Group transfer polymerization of biobased monomers. Eur. Polym. J. 2013, 49, 761–767. [Google Scholar]
- Love, B.E.; Bennett, L.J. Two approaches to the synthesis of dimethyl fumarate that demonstrate fundamental principles of organic chemistry. J. Chem. Educ. 2017, 94, 1543–1546. [Google Scholar] [CrossRef]
- Kees, F. Dimethyl fumarate: A Janus-faced substance? Expert. Opin. Pharm. 2013, 14, 1559–1567. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Topphoff, U.; Varrin-Doyer, M.; Pekarek, K.; Spencer, C.M.; Shetty, A.; Sagan, S.A.; Cree, B.A.; Sobel, R.A.; Wipke, B.T.; Steinman, L.; et al. Dimethyl fumarate treatment induces adaptive and innate immune modulation independent of Nrf2. Proc. Natl. Acad. Sci. USA 2016, 113, 4777–4782. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Li, H.; Sheehy, A.; Cullen, P.; Allaire, N.; Scannevin, R.H. Dimethyl fumarate alters microglia phenotype and protects neurons against proinflammatory toxic microenvironments. J. Neuroimmunol. 2016, 299, 35–44. [Google Scholar] [CrossRef]
- Zhovtis Ryerson, L.; Green, R.; Confident, G.; Pandey, K.; Richter, B.; Bacon, T.; Sammarco, C.; Laing, L.; Kalina, J.; Kister, I. Efficacy and tolerability of dimethyl fumarate in White-, African- and Hispanic- Americans with multiple sclerosis. Ther. Adv. Neurol. Disord. 2016, 9, 454–461. [Google Scholar] [CrossRef] [Green Version]
- Ochi, H.; Niino, M.; Onizuka, Y.; Hiramatsu, K.; Hase, M.; Yun, J.; Matta, A.; Torii, S. 72-Week Safety and Tolerability of Dimethyl Fumarate in Japanese Patients with Relapsing-remitting Multiple Sclerosis: Analysis of the Randomised, Double Blind, Placebo-Controlled, Phase III APEX Study and its Open-Label Extension. Adv. Ther. 2018, 35, 1598–1611. [Google Scholar] [CrossRef] [Green Version]
- Sabin, J.; Urtiaga, S.; Pilo, B.; Thuissard, I.; Galan, V.; Sainz de la Maza, S.; Costa-Frossard, L.; Gómez-Moreno, M.; Díaz-Díaz, J.; Oreja-Guevara, C.; et al. Tolerability and safety of dimethyl fumarate in relapsing multiple sclerosis: A prospective observational multicenter study in a real-life Spanish population. J. Neurol. 2020, 267, 2362–2371. [Google Scholar] [CrossRef]
- Mallucci, G.; Annovazzi, P.; Miante, S.; Torri-Clerici, V.; Matta, M.; La Gioia, S.; Cavarretta, R.; Mantero, V.; Costantini, G.; D’Ambrosio, V.; et al. Two-year real-life efficacy, tolerability and safety of dimethyl fumarate in an Italian multicentre study. J. Neurol. 2018, 265, 1850–1859. [Google Scholar] [CrossRef]
- Litjens, N.H.; Burggraaf, J.; van Strijen, E.; van Gulpen, C.; Mattie, H.; Schoemaker, R.C.; van Dissel, J.T.; Thio, H.B.; Nibbering, P.H. Pharmacokinetics of oral fumarates in healthy subjects. Br. J. Clin. Pharm. 2004, 58, 429–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rostami-Yazdi, M.; Clement, B.; Schmidt, T.J.; Schinor, D.; Mrowietz, U. Detection of metabolites of fumaric acid esters in human urine: Implications for their mode of action. J. Investig. Derm. 2009, 129, 231–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rostami-Yazdi, M.; Clement, B.; Mrowietz, U. Pharmacokinetics of anti-psoriatic fumaric acid esters in psoriasis patients. Arch. Derm. Res. 2010, 302, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Werdenberg, D.; Joshi, R.; Wolffram, S.; Merkle, H.P.; Langguth, P. Presystemic metabolism and intestinal absorption of antipsoriatic fumaric acid esters. Biopharm. Drug. Dispos. 2003, 24, 259–273. [Google Scholar] [CrossRef]
- Palte, M.J.; Wehr, A.; Tawa, M.; Perkin, K.; Leigh-Pemberton, R.; Hanna, J.; Miller, C.; Penner, N. Improving the Gastrointestinal Tolerability of Fumaric Acid Esters: Early Findings on Gastrointestinal Events with Diroximel Fumarate in Patients with Relapsing-Remitting Multiple Sclerosis from the Phase 3, Open-Label EVOLVE-MS-1 Study. Adv. Ther. 2019, 36, 3154–3165. [Google Scholar] [CrossRef] [Green Version]
- Litjens, N.H.; van Strijen, E.; van Gulpen, C.; Mattie, H.; van Dissel, J.T.; Thio, H.B.; Nibbering, P.H. In vitro pharmacokinetics of anti-psoriatic fumaric acid esters. BMC Pharm. 2004, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Dibbert, S.; Clement, B.; Skak-Nielsen, T.; Mrowietz, U.; Rostami-Yazdi, M. Detection of fumarate-glutathione adducts in the portal vein blood of rats: Evidence for rapid dimethylfumarate metabolism. Arc. Derm. Res. 2013, 305, 447–451. [Google Scholar] [CrossRef]
- Canning, P.; Sorrell, F.J.; Bullock, A.N. Structural basis of Keap1 interactions with Nrf2. Free Radic. Biol. Med. 2015, 88 Pt B, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Takaya, K.; Suzuki, T.; Motohashi, H.; Onodera, K.; Satomi, S.; Kensler, T.W.; Yamamoto, M. Validation of the multiple sensor mechanism of the Keap1-Nrf2 system. Free Radic. Biol. Med. 2012, 53, 817–827. [Google Scholar] [CrossRef] [Green Version]
- Unni, S.; Deshmukh, P.; Krishnappa, G.; Kommu, P.; Padmanabhan, B. Structural insights into the multiple binding modes of Dimethyl Fumarate (DMF) and its analogs to the Kelch domain of Keap1. FEBS J. 2021, 288, 1599–1613. [Google Scholar] [CrossRef]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharm. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghunath, A.; Sundarraj, K.; Nagarajan, R.; Arfuso, F.; Bian, J.; Kumar, A.P.; Sethi, G.; Perumal, E. Antioxidant response elements: Discovery, classes, regulation and potential applications. Redox Biol. 2018, 17, 297–314. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.B.; Pandita, R.K.; Eskiocak, U.; Ly, P.; Kaisani, A.; Kumar, R.; Cornelius, C.; Wright, W.E.; Pandita, T.K.; Shay, J.W. Targeting of Nrf2 induces DNA damage signaling and protects colonic epithelial cells from ionizing radiation. Proc. Natl. Acad. Sci. USA 2012, 109, E2949–E2955. [Google Scholar] [CrossRef] [Green Version]
- Jin, S. p53, Autophagy and tumor suppression. Autophagy 2005, 1, 171–173. [Google Scholar] [CrossRef] [Green Version]
- Ni, H.M.; Boggess, N.; McGill, M.R.; Lebofsky, M.; Borude, P.; Apte, U.; Jaeschke, H.; Ding, W.X. Liver-specific loss of Atg5 causes persistent activation of Nrf2 and protects against acetaminophen-induced liver injury. Toxicol. Sci. 2012, 127, 438–450. [Google Scholar] [CrossRef]
- Copple, I.M.; Lister, A.; Obeng, A.D.; Kitteringham, N.R.; Jenkins, R.E.; Layfield, R.; Foster, B.J.; Goldring, C.E.; Park, B.K. Physical and functional interaction of sequestosome 1 with Keap1 regulates the Keap1-Nrf2 cell defense pathway. J. Biol. Chem. 2010, 285, 16782–16788. [Google Scholar] [CrossRef] [Green Version]
- Bae, S.H.; Sung, S.H.; Oh, S.Y.; Lim, J.M.; Lee, S.K.; Park, Y.N.; Lee, H.E.; Kang, D.; Rhee, S.G. Sestrins activate Nrf2 by promoting p62-dependent autophagic degradation of Keap1 and prevent oxidative liver damage. Cell. Metab. 2013, 17, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Jain, A.; Lamark, T.; Sjøttem, E.; Larsen, K.B.; Awuh, J.A.; Øvervatn, A.; McMahon, M.; Hayes, J.D.; Johansen, T. p62/SQSTM1 is a target gene for transcription factor NRF2 and creates a positive feedback loop by inducing antioxidant response element-driven gene transcription. J. Biol. Chem. 2010, 285, 22576–22591. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.; Wang, Z.; Fu, Z.; Ma, H.; Jiang, M.; Xu, A.; Zhang, W. p62/SQSTM1 protects against cisplatin-induced oxidative stress in kidneys by mediating the cross talk between autophagy and the Keap1-Nrf2 signalling pathway. Free Radic. Res. 2019, 53, 800–814. [Google Scholar] [CrossRef]
- Kapuy, O.; Papp, D.; Vellai, T.; Bánhegyi, G.; Korcsmáros, T. Systems-Level Feedbacks of NRF2 Controlling Autophagy upon Oxidative Stress Response. Antioxidants 2018, 7, 39. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Yi, Y.; Xia, G.; Zhao, Y.; Yu, Y.; Li, L.; Hua, C.; He, B.; Yang, B.; Yu, C.; et al. Nrf2-miR-129-3p-mTOR Axis Controls an miRNA Regulatory Network Involved in HDACi-Induced Autophagy. Mol. Ther. 2019, 27, 1039–1050. [Google Scholar] [CrossRef] [PubMed]
- Joo, M.S.; Kim, W.D.; Lee, K.Y.; Kim, J.H.; Koo, J.H.; Kim, S.G. AMPK Facilitates Nuclear Accumulation of Nrf2 by Phosphorylating at Serine 550. Mol. Cell. Biol. 2016, 36, 1931–1942. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, K.; Baldinger, J.; Mayerhofer, B.; Atanasov, A.G.; Dirsch, V.M.; Heiss, E.H. Activated AMPK boosts the Nrf2/HO-1 signaling axis--A role for the unfolded protein response. Free Radic. Biol. Med. 2015, 88 Pt B, 417–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornberg, M.D.; Bhargava, P.; Kim, P.M.; Putluri, V.; Snowman, A.M.; Putluri, N.; Calabresi, P.A.; Snyder, S.H. Dimethyl fumarate targets GAPDH and aerobic glycolysis to modulate immunity. Science 2018, 360, 449–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillard, G.O.; Collette, B.; Anderson, J.; Chao, J.; Scannevin, R.H.; Huss, D.J.; Fontenot, J.D. DMF, but not other fumarates, inhibits NF-κB activity in vitro in an Nrf2-independent manner. J. Neuroimmunol. 2015, 283, 74–85. [Google Scholar] [CrossRef] [Green Version]
- Loewe, R.; Holnthoner, W.; Gröger, M.; Pillinger, M.; Gruber, F.; Mechtcheriakova, D.; Hofer, E.; Wolff, K.; Petzelbauer, P. Dimethylfumarate inhibits TNF-induced nuclear entry of NF-kappa B/p65 in human endothelial cells. J. Immunol. 2002, 168, 4781–4787. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Assmann, J.C.; Krenz, A.; Rahman, M.; Grimm, M.; Karsten, C.M.; Köhl, J.; Offermanns, S.; Wettschureck, N.; Schwaninger, M. Hydroxycarboxylic acid receptor 2 mediates dimethyl fumarate’s protective effect in EAE. J. Clin. Investig. 2014, 124, 2188–2192. [Google Scholar] [CrossRef]
- Parodi, B.; Rossi, S.; Morando, S.; Cordano, C.; Bragoni, A.; Motta, C.; Usai, C.; Wipke, B.T.; Scannevin, R.H.; Mancardi, G.L.; et al. Fumarates modulate microglia activation through a novel HCAR2 signaling pathway and rescue synaptic dysregulation in inflamed CNS. Acta Neuropathol. 2015, 130, 279–295. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Liu, Y.; Fang, J.; Chen, Y.; Li, H.; Gao, K. Dimethyl fumarate inhibits the expression and function of hypoxia-inducible factor-1α (HIF-1α). Biochem. Biophys. Res. Commun. 2014, 448, 303–307. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, W.; Zhang, X.; Lu, P.; Du, Q.; Tao, L.; Ding, Y.; Wang, Y.; Hu, R. Dimethyl fumarate ameliorates dextran sulfate sodium-induced murine experimental colitis by activating Nrf2 and suppressing NLRP3 inflammasome activation. Biochem. Pharm. 2016, 112, 37–49. [Google Scholar] [CrossRef]
- Tastan, B.; Arioz, B.I.; Tufekci, K.U.; Tarakcioglu, E.; Gonul, C.P.; Genc, K.; Genc, S. Dimethyl Fumarate Alleviates NLRP3 Inflammasome Activation in Microglia and Sickness Behavior in LPS-Challenged Mice. Front. Immunol. 2021, 12, 737065. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Gönczi, L.; Lakatos, P.L.; Burisch, J. The Burden of Inflammatory Bowel Disease in Europe in 2020. J. Crohns Colitis 2021, 15, 1573–1587. [Google Scholar] [CrossRef] [PubMed]
- Makharia, G.K.; Singh, P.; Catassi, C.; Sanders, D.S.; Leffler, D.; Ali, R.A.R.; Bai, J.C. The global burden of coeliac disease: Opportunities and challenges. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 313–327. [Google Scholar] [CrossRef] [PubMed]
- Booth, L.; Malkin, M.; Dent, P. Repurposing Tecfidera for cancer. Aging 2016, 8, 1289–1290. [Google Scholar] [CrossRef] [Green Version]
- Lastres-Becker, I.; García-Yagüe, A.J.; Scannevin, R.H.; Casarejos, M.J.; Kügler, S.; Rábano, A.; Cuadrado, A. Repurposing the NRF2 Activator Dimethyl Fumarate as Therapy Against Synucleinopathy in Parkinson’s Disease. Antioxid. Redox Signal. 2016, 25, 61–77. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi-Beni, R.; Najafi, A.; Savar, S.M.; Mohebbi, N.; Khoshnevisan, A. Role of dimethyl fumarate in the treatment of glioblastoma multiforme: A review article. Iran. J. Neurol. 2019, 18, 127–133. [Google Scholar] [CrossRef]
- Garcia-Mesa, Y.; Xu, H.N.; Vance, P.; Gruenewald, A.L.; Garza, R.; Midkiff, C.; Alvarez-Hernandez, X.; Irwin, D.J.; Gill, A.J.; Kolson, D.L. Dimethyl Fumarate, an Approved Multiple Sclerosis Treatment, Reduces Brain Oxidative Stress in SIV-Infected Rhesus Macaques: Potential Therapeutic Repurposing for HIV Neuroprotection. Antioxidants 2021, 10, 416. [Google Scholar] [CrossRef]
- Thomas, S.D.; Jha, N.K.; Sadek, B.; Ojha, S. Repurposing Dimethyl Fumarate for Cardiovascular Diseases: Pharmacological Effects, Molecular Mechanisms, and Therapeutic Promise. Pharmaceuticals 2022, 15, 497. [Google Scholar] [CrossRef]
- Manai, F.; Govoni, S.; Amadio, M. The Challenge of Dimethyl Fumarate Repurposing in Eye Pathologies. Cells 2022, 11, 4061. [Google Scholar] [CrossRef]
- Majkutewicz, I. Dimethyl fumarate: A review of preclinical efficacy in models of neurodegenerative diseases. Eur. J. Pharm. 2022, 926, 175025. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.A.; Barnes, L.H.; Rassman, V.L.; Kelloff, G.V.; Steele, V.E. Use of azoxymethane-induced foci of aberrant crypts in rat colon to identify potential cancer chemopreventive agents. Carcinogenesis 1994, 15, 1049–1054. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Smith, B.L.; Munday, C.M. Effect of inducers of DT-diaphorase on the toxicity of 2-methyl- and 2-hydroxy-1,4-naphthoquinone to rats. Chem. Biol. Interact. 1999, 123, 219–237. [Google Scholar] [CrossRef] [PubMed]
- Begleiter, A.; Sivananthan, K.; Curphey, T.J.; Bird, R.P. Induction of NAD(P)H quinone: Oxidoreductase1 inhibits carcinogen-induced aberrant crypt foci in colons of Sprague-Dawley rats. Cancer Epidemiol. Biomark. Rev. 2003, 12, 566–572. [Google Scholar]
- Kaluzki, I.; Hailemariam-Jahn, T.; Doll, M.; Kaufmann, R.; Balermpas, P.; Zöller, N.; Kippenberger, S.; Meissner, M. Dimethylfumarate Inhibits Colorectal Carcinoma Cell Proliferation: Evidence for Cell Cycle Arrest, Apoptosis and Autophagy. Cells 2019, 8, 1329. [Google Scholar] [CrossRef] [Green Version]
- Kirlin, W.G.; Cai, J.; DeLong, M.J.; Patten, E.J.; Jones, D.P. Dietary compounds that induce cancer preventive phase 2 enzymes activate apoptosis at comparable doses in HT29 colon carcinoma cells. J. Nutr. 1999, 129, 1827–1835. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Zhao, Y.; Ma, C.Y.; Xu, X.M.; Zhang, Y.Q.; Wang, C.G.; Jin, J.; Shen, X.; Gao, J.L.; Li, N.; et al. Dimethyl fumarate induces necroptosis in colon cancer cells through GSH depletion/ROS increase/MAPKs activation pathway. Br. J. Pharm. 2015, 172, 3929–3943. [Google Scholar] [CrossRef] [Green Version]
- Casili, G.; Cordaro, M.; Impellizzeri, D.; Bruschetta, G.; Paterniti, I.; Cuzzocrea, S.; Esposito, E. Dimethyl Fumarate Reduces Inflammatory Responses in Experimental Colitis. J. Crohns Colitis 2016, 10, 472–483. [Google Scholar] [CrossRef] [Green Version]
- Pitarokoili, K.; Bachir, H.; Sgodzai, M.; Grüter, T.; Haupeltshofer, S.; Duscha, A.; Pedreiturria, X.; Motte, J.; Gold, R. Induction of Regulatory Properties in the Intestinal Immune System by Dimethyl Fumarate in Lewis Rat Experimental Autoimmune Neuritis. Front. Immunol. 2019, 10, 2132. [Google Scholar] [CrossRef]
- Gendy, A.; Soubh, A.; Al-Mokaddem, A.; Kotb El-Sayed, M. Dimethyl fumarate protects against intestinal ischemia/reperfusion lesion: Participation of Nrf2/HO-1, GSK-3β and Wnt/β-catenin pathway. Biomed. Pharm. 2021, 134, 111130. [Google Scholar] [CrossRef]
- Mi, Y.; Xie, X.; Bao, Z.; Xiong, X.; Wang, X.; Zhang, H. Dimethyl fumarate protects against intestine damage in necrotizing enterocolitis by inhibiting the Toll-like receptor (TLR) inflammatory signaling pathway. Tissue Cell 2023, 81, 102003. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Takasu, C.; Lau, H.; Robles, L.; Vo, K.; Farzaneh, T.; Vaziri, N.D.; Stamos, M.J.; Ichii, H. Dimethyl Fumarate Alleviates Dextran Sulfate Sodium-Induced Colitis, through the Activation of Nrf2-Mediated Antioxidant and Anti-inflammatory Pathways. Antioxidants 2020, 9, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dingenen, J.; Pieters, L.; Van Nuffel, E.; Lefebvre, R.A. Hemin reduces postoperative ileus in a heme oxygenase 1-dependent manner while dimethyl fumarate does without heme oxygenase 1-induction. Neurogastroenterol. Motil. 2020, 32, e13624. [Google Scholar] [CrossRef] [PubMed]
- Davé, S.H.; Tilstra, J.S.; Matsuoka, K.; Li, F.; DeMarco, R.A.; Beer-Stolz, D.; Sepulveda, A.R.; Fink, M.P.; Lotze, M.T.; Plevy, S.E. Ethyl pyruvate decreases HMGB1 release and ameliorates murine colitis. J. Leukoc. Biol. 2009, 86, 633–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djedović, N.; Stanisavljevic, S.; Jevtić, B.; Momčilović, M.; Lavrnja, I.; Miljković, D. Anti-encephalitogenic effects of ethyl pyruvate are reflected in the central nervous system and the gut. Biomed. Pharm. 2017, 96, 78–85. [Google Scholar] [CrossRef]
- Ma, N.; Wu, Y.; Xie, F.; Du, K.; Wang, Y.; Shi, L.; Ji, L.; Liu, T.; Ma, X. Dimethyl fumarate reduces the risk of mycotoxins via improving intestinal barrier and microbiota. Oncotarget 2017, 8, 44625–44638. [Google Scholar] [CrossRef]
- Chen, H.; Wu, X.; Sun, R.; Lu, H.; Lin, R.; Gao, X.; Li, G.; Feng, Z.; Zhu, R.; Yao, Y.; et al. Dysregulation of CD177+ neutrophils on intraepithelial lymphocytes exacerbates gut inflammation via decreasing microbiota-derived DMF. Gut Microbes 2023, 15, 2172668. [Google Scholar] [CrossRef]
- Buscarinu, M.C.; Gargano, F.; Lionetto, L.; Capi, M.; Morena, E.; Fornasiero, A.; Reniè, R.; Landi, A.C.; Pellicciari, G.; Romano, C.; et al. Intestinal Permeability and Circulating CD161+CCR6+CD8+T Cells in Patients With Relapsing-Remitting Multiple Sclerosis Treated With Dimethylfumarate. Front. Neurol. 2021, 12, 683398. [Google Scholar] [CrossRef]
- Patel, V.; Joharapurkar, A.; Kshirsagar, S.; Patel, M.; Savsani, H.; Patel, A.; Ranvir, R.; Jain, M. Repurposing dimethyl fumarate for gastric ulcer and ulcerative colitis: Evidence of local efficacy without systemic side effect. Med. Drug. Discov. 2022, 16, 100142. [Google Scholar] [CrossRef]
- Sangineto, M.; Grabherr, F.; Adolph, T.E.; Grander, C.; Reider, S.; Jaschke, N.; Mayr, L.; Schwärzler, J.; Dallio, M.; Moschen, A.R.; et al. Dimethyl fumarate ameliorates hepatic inflammation in alcohol related liver disease. Liver Int. 2020, 40, 1610–1619. [Google Scholar] [CrossRef] [Green Version]
- Eppinga, H.; Thio, H.B.; Schreurs, M.W.J.; Blakaj, B.; Tahitu, R.I.; Konstantinov, S.R.; Peppelenbosch, M.P.; Fuhler, G.M. Depletion of Saccharomyces cerevisiae in psoriasis patients, restored by Dimethylfumarate therapy (DMF). PLoS ONE 2017, 12, e0176955. [Google Scholar] [CrossRef] [PubMed]
- Herbrecht, R.; Nivoix, Y. Saccharomyces cerevisiae fungemia: An adverse effect of Saccharomyces boulardii probiotic administration. Clin. Infect. Dis. 2005, 40, 1635–1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Torrado, R.; Querol, A. Opportunistic Strains of Saccharomyces cerevisiae: A Potential Risk Sold in Food Products. Front. Microbiol. 2016, 6, 1522. [Google Scholar] [CrossRef] [Green Version]
- Rannikko, J.; Holmberg, V.; Karppelin, M.; Arvola, P.; Huttunen, R.; Mattila, E.; Kerttula, N.; Puhto, T.; Tamm, Ü.; Koivula, I.; et al. Fungemia and Other Fungal Infections Associated with Use of Saccharomyces boulardii Probiotic Supplements. Emerg. Infect. Dis. 2021, 27, 2090–2096. [Google Scholar] [CrossRef] [PubMed]
- Lamps, L.W.; Lai, K.K.; Milner, D.A., Jr. Fungal infections of the gastrointestinal tract in the immunocompromised host: An update. Adv. Anat. Pathol. 2014, 21, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Rumah, K.R.; Vartanian, T.K.; Fischetti, V.A. Oral Multiple Sclerosis Drugs Inhibit the In vitro Growth of Epsilon Toxin Producing Gut Bacterium, Clostridium perfringens. Front. Cell. Infect. Microbiol. 2017, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadovnikova, I.S.; Gureev, A.P.; Ignatyeva, D.A.; Gryaznova, M.V.; Chernyshova, E.V.; Krutskikh, E.P.; Novikova, A.G.; Popov, V.N. Nrf2/ARE Activators Improve Memory in Aged Mice via Maintaining of Mitochondrial Quality Control of Brain and the Modulation of Gut Microbiome. Pharmaceuticals 2021, 14, 607. [Google Scholar] [CrossRef]
- Shah, S.; Locca, A.; Dorsett, Y.; Cantoni, C.; Ghezzi, L.; Lin, Q.; Bokoliya, S.; Panier, H.; Suther, C.; Gormley, M.; et al. Alterations of the gut mycobiome in patients with MS. EBioMedicine 2021, 71, 103557. [Google Scholar] [CrossRef]
- Katz Sand, I.; Zhu, Y.; Ntranos, A.; Clemente, J.C.; Cekanaviciute, E.; Brandstadter, R.; Crabtree-Hartman, E.; Singh, S.; Bencosme, Y.; Debelius, J.; et al. Disease-modifying therapies alter gut microbial composition in MS. Neurol. Neuroimmunol. Neuroinflamm. 2018, 6, e517. [Google Scholar] [CrossRef] [Green Version]
- Storm-Larsen, C.; Myhr, K.M.; Farbu, E.; Midgard, R.; Nyquist, K.; Broch, L.; Berg-Hansen, P.; Buness, A.; Holm, K.; Ueland, T.; et al. Gut microbiota composition during a 12-week intervention with delayed-release dimethyl fumarate in multiple sclerosis—A pilot trial. Mult. Scler. J. Exp. Transl. Clin. 2019, 5, 2055217319888767. [Google Scholar] [CrossRef] [Green Version]
- Ferri, C.; Castellazzi, M.; Merli, N.; Laudisi, M.; Baldin, E.; Baldi, E.; Mancabelli, L.; Ventura, M.; Pugliatti, M. Gut Microbiota Changes during Dimethyl Fumarate Treatment in Patients with Multiple Sclerosis. Int. J. Mol. Sci. 2023, 24, 2720. [Google Scholar] [CrossRef] [PubMed]
- Diebold, M.; Meola, M.; Purushothaman, S.; Siewert, L.K.; Pössnecker, E.; Roloff, T.; Lindberg, R.L.; Kuhle, J.; Kappos, L.; Derfuss, T.; et al. Gut microbiota composition as a candidate risk factor for dimethyl fumarate-induced lymphopenia in multiple sclerosis. Gut Microbes 2022, 14, 2147055. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2017, 390, 2769–2778. [Google Scholar] [CrossRef] [PubMed]
- Abraham, C.; Cho, J.H. Inflammatory bowel disease. N. Eng. J. Med. 2009, 361, 2066–2078. [Google Scholar] [CrossRef] [PubMed]
- Baumgart, D.C.; Sandborn, W.J. Crohn’s disease. Lancet 2012, 380, 1590–1605. [Google Scholar] [CrossRef] [Green Version]
- Ungaro, R.; Mehandru, S.; Allen, P.B.; Peyrin-Biroulet, L.; Colombel, J.F. Ulcerative colitis. Lancet 2017, 389, 1756–1770. [Google Scholar] [CrossRef]
- Khor, B.; Gardet, A.; Xavier, R.J. Genetics and pathogenesis of inflammatory bowel disease. Nature 2011, 474, 307–317. [Google Scholar] [CrossRef] [Green Version]
- Gajendran, M.; Loganathan, P.; Catinella, A.P.; Hashash, J.G. A comprehensive review and update on Crohn’s disease. Dis. Mon. 2018, 64, 20–57. [Google Scholar] [CrossRef]
- Qin, X. Why is damage limited to the mucosa in ulcerative colitis but transmural in Crohn’s disease? World J. Gastrointest. Pathophysiol. 2013, 4, 63–64. [Google Scholar] [CrossRef]
- Huang, H.; Fang, M.; Jostins, L.; Umićević Mirkov, M.; Boucher, G.; Anderson, C.A.; Andersen, V.; Cleynen, I.; Cortes, A.; Crins, F.; et al. Fine-mapping inflammatory bowel disease loci to single-variant resolution. Nature 2017, 547, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Peters, L.A.; Perrigoue, J.; Mortha, A.; Iuga, A.; Song, W.M.; Neiman, E.M.; Llewellyn, S.R.; Di Narzo, A.; Kidd, B.A.; Telesco, S.E.; et al. A functional genomics predictive network model identifies regulators of inflammatory bowel disease. Nat. Genet. 2017, 49, 1437–1449. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Cui, Y.; Zhang, Y.; Zhao, T.; Cong, J. Fecal microbiota transplantation for induction of remission in Crohn’s disease: A systematic review and meta-analysis. Int. J. Color. Dis. 2023, 38, 62. [Google Scholar] [CrossRef] [PubMed]
- van Lingen, E.E.; Baunwall, S.S.M.D.; Lieberknecht, S.S.C.; Benech, N.N.; Ianiro, G.G.; Sokol, H.H.; Gasbarrini, A.A.; Cammarota, G.G.; Eriksen, M.M.K.; van der Meulen-de Jong, A.A.E.; et al. Short- and long-term follow-up after fecal microbiota transplantation as treatment for recurrent Clostridioides difficile infection in patients with inflammatory bowel disease. Ther. Adv. Gastroenterol. 2023, 16, 17562848231156285. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.; Parsa, S.; Garza, M.; Quigley, E.M.M.; Abraham, B.P. The Role of Fecal Microbiota Transplantation in the Induction of Remission in Ulcerative Colitis. Dig. Dis. 2023. [Google Scholar] [CrossRef]
- Roda, G.; Chien Ng, S.; Kotze, P.G.; Argollo, M.; Panaccione, R.; Spinelli, A.; Kaser, A.; Peyrin-Biroulet, L.; Danese, S. Crohn’s disease. Nat. Rev. Dis. Prim. 2020, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Petagna, L.; Antonelli, A.; Ganini, C.; Bellato, V.; Campanelli, M.; Divizia, A.; Efrati, C.; Franceschilli, M.; Guida, A.M.; Ingallinella, S.; et al. Pathophysiology of Crohn’s disease inflammation and recurrence. Biol. Direct 2020, 15, 23. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Wang, S.; Li, J. Treatment of Inflammatory Bowel Disease: A Comprehensive Review. Front. Med. 2021, 8, 765474. [Google Scholar] [CrossRef]
- Di Sabatino, A.; Corazza, G.R. Coeliac disease. Lancet 2009, 373, 1480–1493. [Google Scholar] [CrossRef]
- Yan, D.; Duermeyer, L.; Leoveanu, C.; Nambara, E. The functions of the endosperm during seed germination. Plant. Cell. Physiol. 2014, 55, 1521–1533. [Google Scholar] [CrossRef]
- Drago, S.; El Asmar, R.; Di Pierro, M.; Grazia Clemente, M.; Tripathi, A.; Sapone, A.; Thakar, M.; Iacono, G.; Carroccio, A.; D’Agate, C.; et al. Gliadin, zonulin and gut permeability: Effects on celiac and non-celiac intestinal mucosa and intestinal cell lines. Scand. J. Gastroenterol. 2006, 41, 408–419. [Google Scholar] [CrossRef]
- Siegel, M.; Strnad, P.; Watts, R.E.; Choi, K.; Jabri, B.; Omary, M.B.; Khosla, C. Extracellular transglutaminase 2 is catalytically inactive, but is transiently activated upon tissue injury. PLoS ONE 2008, 3, e1861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molberg, O.; Mcadam, S.N.; Körner, R.; Quarsten, H.; Kristiansen, C.; Madsen, L.; Fugger, L.; Scott, H.; Norén, O.; Roepstorff, P.; et al. Tissue transglutaminase selectively modifies gliadin peptides that are recognized by gut-derived T cells in celiac disease. Nat. Med. 1998, 4, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, D. Human intestinal dendritic cells as controllers of mucosal immunity. Rev. Esp. Enferm. Dis. 2013, 105, 279–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escudero-Hernández, C.; Peña, A.S.; Bernardo, D. Immunogenetic Pathogenesis of Celiac Disease and Non-celiac Gluten Sensitivity. Curr. Gastroenterol. Rep. 2016, 18, 36. [Google Scholar] [CrossRef] [PubMed]
- Tjon, J.M.; van Bergen, J.; Koning, F. Celiac disease: How complicated can it get? Immunogenetics 2010, 62, 641–651. [Google Scholar] [CrossRef] [Green Version]
- Kopacz, A.; Kloska, D.; Klimczyk, D.; Kopec, M.; Jozkowicz, A.; Piechota-Polanczyk, A. Nrf2 Transcriptional Activity Governs Intestine Development. Int. J. Mol. Sci. 2022, 23, 6175. [Google Scholar] [CrossRef]
- Yagishita, Y.; McCallum, M.L.; Kensler, T.W.; Wakabayashi, N. Constitutive Activation of Nrf2 in Mice Expands Enterogenesis in Small Intestine Through Negative Regulation of Math1. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 503–524. [Google Scholar] [CrossRef]
- Piotrowska, M.; Swierczynski, M.; Fichna, J.; Piechota-Polanczyk, A. The Nrf2 in the pathophysiology of the intestine: Molecular mechanisms and therapeutic implications for inflammatory bowel diseases. Pharm. Res. 2021, 163, 105243. [Google Scholar] [CrossRef]
- Wen, Z.; Liu, W.; Li, X.; Chen, W.; Liu, Z.; Wen, J.; Liu, Z. A Protective Role of the NRF2-Keap1 Pathway in Maintaining Intestinal Barrier Function. Oxid. Med. Cell. Longev. 2019, 2019, 1759149. [Google Scholar] [CrossRef] [Green Version]
- Yanaka, A. Contribution of NRF2 in Gastrointestinal Protection from Oxidative Injury. Curr. Pharm. Des. 2018, 24, 2023–2033. [Google Scholar] [CrossRef]
- Khor, T.O.; Huang, M.T.; Kwon, K.H.; Chan, J.Y.; Reddy, B.S.; Kong, A.N. Nrf2-deficient mice have an increased susceptibility to dextran sulfate sodium-induced colitis. Cancer Res. 2006, 66, 11580–11584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerstgrasser, A.; Melhem, H.; Leonardi, I.; Atrott, K.; Schäfer, M.; Werner, S.; Rogler, G.; Frey-Wagner, I. Cell-specific Activation of the Nrf2 Antioxidant Pathway Increases Mucosal Inflammation in Acute but Not in Chronic Colitis. J. Crohns Colitis 2017, 11, 485–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Cui, C.; Li, N.; Sun, X.; Wen, L.; Gao, E.; Wang, F. Antioxidant activity and protective effect of wheat germ peptides in an in vitro celiac disease model via Keap1/Nrf2 signaling pathway. Food Res. Int. 2022, 161, 111864. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Xie, Y.; Yuan, J.; Gao, J.; Xiao, Z.; Wu, Y.; Chen, H. The Nutritional Intervention of Resveratrol Can Effectively Alleviate the Intestinal Inflammation Associated with Celiac Disease Induced by Wheat Gluten. Front. Immunol. 2022, 13, 878186. [Google Scholar] [CrossRef]
- Bergamo, P.; Palmieri, G.; Cocca, E.; Ferrandino, I.; Gogliettino, M.; Monaco, A.; Maurano, F.; Rossi, M. Adaptive response activated by dietary cis9, trans11 conjugated linoleic acid prevents distinct signs of gliadin-induced enteropathy in mice. Eur. J. Nutr. 2016, 55, 729–740. [Google Scholar] [CrossRef]
- Kang, S.; Kim, W.; Jeong, S.; Lee, Y.; Nam, J.; Lee, S.; Jung, Y. Oxidized 5-aminosalicylic acid activates Nrf2-HO-1 pathway by covalently binding to Keap1: Implication in anti-inflammatory actions of 5-aminosalicylic acid. Free Radic. Biol. Med. 2017, 108, 715–724. [Google Scholar] [CrossRef]
- Hosseini, A.; Masjedi, A.; Baradaran, B.; Hojjat-Farsangi, M.; Ghalamfarsa, G.; Anvari, E.; Jadidi-Niaragh, F. Dimethyl fumarate: Regulatory effects on the immune system in the treatment of multiple sclerosis. J. Cell. Physiol. 2019, 234, 9943–9955. [Google Scholar] [CrossRef]
- Djedovic, N.; Mansilla, M.J.; Jevtić, B.; Navarro-Barriuso, J.; Saksida, T.; Martínez-Cáceres, E.M.; Miljković, Ð. Ethyl Pyruvate Induces Tolerogenic Dendritic Cells. Front. Immunol. 2019, 10, 157. [Google Scholar] [CrossRef]
- Mazzola, M.A.; Raheja, R.; Regev, K.; Beynon, V.; von Glehn, F.; Paul, A.; Pierre, I.; Kivisakk, P.; Weiner, H.L.; Gandhi, R. Monomethyl fumarate treatment impairs maturation of human myeloid dendritic cells and their ability to activate T cells. Mult. Scler. 2019, 25, 63–71. [Google Scholar] [CrossRef]
- Neunlist, M.; Van Landeghem, L.; Mahé, M.M.; Derkinderen, P.; des Varannes, S.B.; Rolli-Derkinderen, M. The digestive neuronal-glial-epithelial unit: A new actor in gut health and disease. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 90–100. [Google Scholar] [CrossRef]
- Vergnolle, N.; Cirillo, C. Neurons and Glia in the Enteric Nervous System and Epithelial Barrier Function. Physiology 2018, 33, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Pochard, C.; Coquenlorge, S.; Freyssinet, M.; Naveilhan, P.; Bourreille, A.; Neunlist, M.; Rolli-Derkinderen, M. The multiple faces of inflammatory enteric glial cells: Is Crohn’s disease a gliopathy? Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G1–G11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, G.; Cirillo, C.; Sarnelli, G.; De Filippis, D.; D’Armiento, F.P.; Rocco, A.; Nardone, G.; Petruzzelli, R.; Grosso, M.; Izzo, P.; et al. Enteric glial-derived S100B protein stimulates nitric oxide production in celiac disease. Gastroenterology 2007, 133, 918–925. [Google Scholar] [CrossRef] [PubMed]
- Murphy, T.H.; Yu, J.; Ng, R.; Johnson, D.A.; Shen, H.; Honey, C.R.; Johnson, J.A. Preferential expression of antioxidant response element mediated gene expression in astrocytes. J. Neurochem. 2001, 76, 1670–1678. [Google Scholar] [CrossRef]
- Graber, D.J.; Hickey, W.F.; Stommel, E.W.; Harris, B.T. Anti-inflammatory efficacy of dexamethasone and Nrf2 activators in the CNS using brain slices as a model of acute injury. J. Neuroimmune Pharm. 2012, 7, 266–278. [Google Scholar] [CrossRef]
- Kalinin, S.; Polak, P.E.; Lin, S.X.; Braun, D.; Guizzetti, M.; Zhang, X.; Rubinstein, I.; Feinstein, D.L. Dimethyl fumarate regulates histone deacetylase expression in astrocytes. J. Neuroimmunol. 2013, 263, 13–19. [Google Scholar] [CrossRef]
- Paraiso, H.C.; Kuo, P.C.; Curfman, E.T.; Moon, H.J.; Sweazey, R.D.; Yen, J.H.; Chang, F.L.; Yu, I.C. Dimethyl fumarate attenuates reactive microglia and long-term memory deficits following systemic immune challenge. J. Neuroinflammation 2018, 15, 100. [Google Scholar] [CrossRef]
- Liu, L.; Vollmer, M.K.; Ahmad, A.S.; Fernandez, V.M.; Kim, H.; Doré, S. Pretreatment with Korean red ginseng or dimethyl fumarate attenuates reactive gliosis and confers sustained neuroprotection against cerebral hypoxic-ischemic damage by an Nrf2-dependent mechanism. Free Radic. Biol. Med. 2019, 131, 98–114. [Google Scholar] [CrossRef]
- Pascal, V.; Pozuelo, M.; Borruel, N.; Casellas, F.; Campos, D.; Santiago, A.; Martinez, X.; Varela, E.; Sarrabayrouse, G.; Machiels, K.; et al. A microbial signature for Crohn’s disease. Gut 2017, 66, 813–822. [Google Scholar] [CrossRef] [Green Version]
- Breton, J.; Tanes, C.; Tu, V.; Albenberg, L.; Rowley, S.; Devas, N.; Hwang, R.; Kachelries, K.; Wu, G.D.; Baldassano, R.N.; et al. A Microbial Signature for Paediatric Perianal Crohn’s Disease. J. Crohns Colitis 2022, 16, 1281–1292. [Google Scholar] [CrossRef]
- Nistal, E.; Caminero, A.; Herrán, A.R.; Arias, L.; Vivas, S.; de Morales, J.M.; Calleja, S.; de Miera, L.E.; Arroyo, P.; Casqueiro, J. Differences of small intestinal bacteria populations in adults and children with/without celiac disease: Effect of age, gluten diet, and disease. Inflamm. Bowel Dis. 2012, 18, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; De Angelis, M.; De Pasquale, I.; Ndagijimana, M.; Vernocchi, P.; Ricciuti, P.; Gagliardi, F.; Laghi, L.; Crecchio, C.; Guerzoni, M.E.; et al. Duodenal and faecal microbiota of celiac children: Molecular, phenotype and metabolome characterization. BMC Microbiol. 2011, 11, 219. [Google Scholar] [CrossRef] [Green Version]
- Schippa, S.; Iebba, V.; Barbato, M.; Di Nardo, G.; Totino, V.; Checchi, M.P.; Longhi, C.; Maiella, G.; Cucchiara, S.; Conte, M.P. A distinctive ‘microbial signature’ in celiac pediatric patients. BMC Microbiol. 2010, 10, 175. [Google Scholar] [CrossRef] [PubMed]
- Caminero, A.; Galipeau, H.J.; McCarville, J.L.; Johnston, C.W.; Bernier, S.P.; Russell, A.K.; Jury, J.; Herran, A.R.; Casqueiro, J.; Tye-Din, J.A.; et al. Duodenal Bacteria From Patients With Celiac Disease and Healthy Subjects Distinctly Affect Gluten Breakdown and Immunogenicity. Gastroenterology 2016, 151, 670–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Xie, X.R.; Wu, H.; Li, H.; Chi, J.S.; Liu, X.M.; Luo, J.; Tang, Y.; Xu, C.X. Mesenchymal Stem Cells Promote Intestinal Mucosal Repair by Positively Regulating the Nrf2/Keap1/ARE Signaling Pathway in Acute Experimental Colitis. Dig. Dis. Sci. 2023, 68, 1835–1846. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Liu, L.; Liu, X.; Chen, H.; Liu, K.; Huang, N.; Wang, Y. Epoxymicheliolide prevents dextran sulfate sodium-induced colitis in mice by inhibiting TAK1-NF-κB pathway and activating Keap1-NRF2 signaling in macrophages. Int. Immunopharmacol. 2022, 113 Pt A, 109404. [Google Scholar] [CrossRef]
- Yang, N.; Xia, Z.; Shao, N.; Li, B.; Xue, L.; Peng, Y.; Zhi, F.; Yang, Y. Carnosic acid prevents dextran sulfate sodium-induced acute colitis associated with the regulation of the Keap1/Nrf2 pathway. Sci. Rep. 2017, 7, 11036. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Ma, X.; He, Y.; Wang, Y.; Zhong, T.; Zhang, Y. Anti-inflammatory and anti-oxidant properties of Melianodiol on DSS-induced ulcerative colitis in mice. PeerJ 2022, 10, e14209. [Google Scholar] [CrossRef]
- Alsharif, I.A.; Fayed, H.M.; Abdel-Rahman, R.F.; Abd-Elsalam, R.M.; Ogaly, H.A. Miconazole Mitigates Acetic Acid-Induced Experimental Colitis in Rats: Insight into Inflammation, Oxidative Stress and Keap1/Nrf-2 Signaling Crosstalk. Biology 2022, 11, 303. [Google Scholar] [CrossRef]
- Guo, F.; Tsao, R.; Li, C.; Wang, X.; Zhang, H.; Jiang, L.; Sun, Y.; Xiong, H. Green Pea (Pisum sativum L.) Hull Polyphenol Extracts Ameliorate DSS-Induced Colitis through Keap1/Nrf2 Pathway and Gut Microbiota Modulation. Foods 2021, 10, 2765. [Google Scholar] [CrossRef]
- Wang, X.; Quan, S.; Li, J.; Liu, Y.; Sun, H.; Zhang, J.; Liu, D. Protective Effects of Grape Seed Proanthocyanidin Extract in Preventing DSS Induced Ulcerative Colitis Based on Pharmacodynamic, Pharmacokinetic and Tissue Distribution. Curr. Drug. Metab. 2022, 23, 496–505. [Google Scholar] [PubMed]
- Wang, N.; Cui, C.; Xu, C.; Ren, H.; Wang, F.; Yu, Q.; Zhang, G. Effect and mechanism of peanut skin proanthocyanidins on gliadin-induced Caco-2 celiac disease model cells. Clin. Immunol. 2022, 245, 109100. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.Y.; Wilkinson, J. A salutary case of Fumaderm potentially masking the symptoms of bowel cancer and partial bowel obstruction. Br. J. Derm. 2007, 157, 825–826. [Google Scholar] [CrossRef] [PubMed]
- Kofler, L.; Kathrein-Schneider, S.; Schweinzer, K.; Kofler, H. Fumaric acid: A possible new therapy for macular edema? Int. Ophthalmol. 2019, 39, 1627–1631. [Google Scholar] [CrossRef] [PubMed]
- Wynn, D.; Lategan, T.W.; Sprague, T.N.; Rousseau, F.S.; Fox, E.J. Monomethyl fumarate has better gastrointestinal tolerability profile compared with dimethyl fumarate. Mult. Scler. Relat. Disord. 2020, 45, 102335. [Google Scholar] [CrossRef]
- Shakya, A.; Soni, U.K.; Rai, G.; Chatterjee, S.S.; Kumar, V. Gastro-protective and Anti-stress Efficacies of Monomethyl Fumarate and a Fumaria indica Extract in Chronically Stressed Rats. Cell. Mol. Neurobiol. 2016, 36, 621–635. [Google Scholar] [CrossRef]
- Brennan, M.S.; Matos, M.F.; Li, B.; Hronowski, X.; Gao, B.; Juhasz, P.; Rhodes, K.J.; Scannevin, R.H. Dimethyl fumarate and monoethyl fumarate exhibit differential effects on KEAP1, NRF2 activation, and glutathione depletion in vitro. PLoS ONE 2015, 10, e0120254. [Google Scholar] [CrossRef]
- Wipke, B.T.; Hoepner, R.; Strassburger-Krogias, K.; Thomas, A.M.; Gianni, D.; Szak, S.; Brennan, M.S.; Pistor, M.; Gold, R.; Chan, A.; et al. Different Fumaric Acid Esters Elicit Distinct Pharmacologic Responses. Neurol. Neuroimmunol. Neuroinflamm. 2021, 8, e950. [Google Scholar] [CrossRef]
- Naismith, R.T.; Wundes, A.; Ziemssen, T.; Jasinska, E.; Freedman, M.S.; Lembo, A.J.; Selmaj, K.; Bidollari, I.; Chen, H.; Hanna, J.; et al. EVOLVE-MS-2 Study Group Diroximel Fumarate Demonstrates an Improved Gastrointestinal Tolerability Profile Compared with Dimethyl Fumarate in Patients with Relapsing-Remitting Multiple Sclerosis: Results from the Randomized, Double-Blind, Phase III EVOLVE-MS-2 Study. CNS Drugs 2020, 34, 185–196. [Google Scholar]
- Motterlini, R.; Nikam, A.; Manin, S.; Ollivier, A.; Wilson, J.L.; Djouadi, S.; Muchova, L.; Martens, T.; Rivard, M.; Foresti, R. HYCO-3, a dual CO-releaser/Nrf2 activator, reduces tissue inflammation in mice challenged with lipopolysaccharide. Redox Biol. 2019, 20, 334–348. [Google Scholar] [CrossRef]
- Aihara, E.; Engevik, K.A.; Montrose, M.H. Trefoil Factor Peptides and Gastrointestinal Function. Annu. Rev. Physiol. 2017, 79, 357–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Lin, Z.; Lin, Q.; Bei, W.; Guo, J. Pathological and therapeutic roles of bioactive peptide trefoil factor 3 in diverse diseases: Recent progress and perspective. Cell. Death Dis. 2022, 13, 62. [Google Scholar] [CrossRef] [PubMed]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayele, H.K.; Balesaria, S.; Srai, S.K. Phytoestrogens modulate hepcidin expression by Nrf2: Implications for dietary control of iron absorption. Free Radic. Biol. Med. 2015, 89, 1192–1202. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author and Year | Molecule | Type of Study | Model | Described Effects |
|---|---|---|---|---|
| Pereira et al., 1994 [52] | DMF | In vitro | Fisher rats (ACF model) | ↓ ACF |
| Munday et al., 1999 [53] | DMF | In vivo | Sprague-Dawley rats (treated with naphthoquinone) | ↑ DT-diaphorase |
| Kirlin et al., 1999 [56] | DMF | In vitro | HT-29 cells | ↑ Apoptosis |
| Begleiter et al., 2003 [54] | DMF | In vivo | Sprague-Dawley rats (ACF model) | ↑ NQO1, GST; No effects on UGT |
| Davé et al., 2009 [64] | EP | In vivo/in vitro | Murine IL-10 KO chronic colitis model, murine macrophages | ↑ HMGB1, HO-1; ↓ RAGE, IL-12p40, NO, NF-κB DNA binding |
| Xie et al., 2015 [57] | DMF | In vitro | CT-26, HT-29, HCT-116, and SGC-7901 cells | ↓ GSH, viability; ↑ JNK, p38, Erk1/2, autophagy markers |
| Liu et al., 2016 [41] | DMF | In vivo/in vitro | DSS-induced colitis model (wt or Nrf2−/− C57BL/J mice) Bone marrow cells Human THP-1 cells | ↓ Body weight loss, colon shortening, MPO, iNOS, IL-1β, TNF-α, IL-6, caspase-1, NLRP3, mROS, mtDNA release; ↑ GSH, Nrf2 |
| Casili et al., 2016 [58] | DMF | In vivo/in vitro | Experimental colitis (IL-10KO and wt mice, CD-1 mice treated with DNBS) | ↓ Body weight loss, diarrhea, colon damage, MPO, TNF-α, ICAM-1; ↑ Mn-SOD, ZO-1, HO-1 impairment of NF-κB |
| Djedović et al., 2017 [65] | EP | In vivo | EAE mouse model | ↓ T cells, APCs, Th1/Th17-related molecules in mesenteric lymph nodes and Peyer patches |
| Ma et al., 2017 [66] | DMF | In vivo | BALB/c mice | ↑ Growth and morphology of intestinal mucosa; ↓ IEB permeability, mycotoxins |
| Kaluzki et al., 2019 [55] | DMF | In vitro | HT-29 and T84 cells | ↓ Proliferation, cyclin D1, CDK4; ↑ p21, autophagy and apoptotic markers |
| Pitarokoili et al., 2019 [59] | DMF | In vivo/ex vivo | Experimental autoimmune neuritis (Lewis rats) | ↓ TLR-4 and IFN-γ mRNA; ↑ Nrf2, HO-1, and FoxP3 mRNA; regulatory T cells in Peyer patches |
| Li et al., 2020 [62] | DMF | In vivo | DSS-induced colitis model | ↓ Colon shortening, inflammation, COX-2; ↑GCLC, GPX |
| Van Dingenen et al., 2020 [63] | DMF | In vivo | Post-operative ileus model (C57BL/J mice under IM) | ↓ Delayed transit; ↓ IL-6, lymphocyte infiltration, Erk-1/2, NF-κB |
| Sangineto et al., 2020 [70] | DMF | In vivo | Ethanol-induced gastric ulcer model | Protection against gut barrier dysfunction and LPS translocation |
| Gendy et al., 2021 [60] | DMF | In vivo | Ischemia/reperfusion model (Wistar rats) | ↑ Nrf2, HO-1, Bcl-2; ↓ GSK-3β, MDA, iNOS, NF-κB, MPO, TNF-α, IL-1β, P-selectin, caspase-3 |
| Buscarinu et al., 2021 [68] | DMF | Observational study | MS patients | Variable effects on gut barrier alterations |
| Patel et al., 2022 [69] | DMF | In vivo | Ethanol-induced gastric ulcer model | ↓ Thiobarbituric acid reactive substance levels; protection against ulcers |
| Mi et al., 2023 [61] | DMF | In vivo | Necrotizing enterocolitis (C57BL/J mice treated with hypoxia and LPS) | ↓ Weight loss, diarrhea, IL-6, IL-1β, TNF-α, TLR-4, NF-κB, Bax, caspase 3/9; ↑ Bcl-2 |
| Chen et al., 2023 [67] | DMF | In vivo | DSS-induced colitis or C. rodentium infection | ↓ Intestinal inflammation, gasdermin D-induced pyroptosis of IELs |
| Author and Year | Molecule | Type of Study | Model | Described Effects |
|---|---|---|---|---|
| Eppinga et al., 2017 [71] | DMF | In vivo/in vitro | Fecal samples from psoriasis patients; S. cerevisiae | ↑ S. cerevisiae in DMF-treated patients |
| Rumah et al., 2017 [76] | DMF | In vitro | C. perfringens | ↓ Growth |
| Ma et al., 2017 [66] | DMF | In vivo | BALB/c mice | ↑ Growth and morphology of intestinal mucosa; ↓ IEB permeability, mycotoxins |
| Katz Sand et al., 2018 [79] | DMF | Observational study | MS patients | ↓ Lachnospiraceae, Veillonellaceae, Firmicutes, Fusobacteria, Clostridiales; ↑ Bacteroidetes |
| Storm-Larsen et al., 2019 [80] | DMF | Observational study | MS patients | Reduced trend of Actinobacteria, ↑ Firmicutes (Faecalibacterium) |
| Sadnovnikova et al., 2021 [77] | DMF | In vivo | C57BL/J mice | ↑ Mitochondrial biogenesis, mitophagy, Nrf2/ARE pathway; modification of gut microbiota |
| Shah et al., 2021 [78] | DMF | Observational study | MS patients | No changes in gut mycobiota |
| Diebold et al., 2022 [82] | DMF | Observational study | MS patients | ↓ Akkermansia muciniphilia and Coprococcus eutactus; ↑ Lactobacillus pentosus |
| Ferri et al., 2023 [81] | DMF | Observational study | MS patients | ↓ Clostridium; ↑ Streptococcus, Haemophilus, Clostridium, Lachnospira, Blautia, Subdoligranulum and ↓ Bacteroidetes, Barnesiella, Odoribacter, Akkermansia in patients with side effects |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manai, F.; Zanoletti, L.; Arfini, D.; Micco, S.G.D.; Gjyzeli, A.; Comincini, S.; Amadio, M. Dimethyl Fumarate and Intestine: From Main Suspect to Potential Ally against Gut Disorders. Int. J. Mol. Sci. 2023, 24, 9912. https://doi.org/10.3390/ijms24129912
Manai F, Zanoletti L, Arfini D, Micco SGD, Gjyzeli A, Comincini S, Amadio M. Dimethyl Fumarate and Intestine: From Main Suspect to Potential Ally against Gut Disorders. International Journal of Molecular Sciences. 2023; 24(12):9912. https://doi.org/10.3390/ijms24129912
Chicago/Turabian StyleManai, Federico, Lisa Zanoletti, Davide Arfini, Simone Giorgio De Micco, Arolda Gjyzeli, Sergio Comincini, and Marialaura Amadio. 2023. "Dimethyl Fumarate and Intestine: From Main Suspect to Potential Ally against Gut Disorders" International Journal of Molecular Sciences 24, no. 12: 9912. https://doi.org/10.3390/ijms24129912