
Partner, Neighbor, Housekeeper and Dimension: 3D versus 2D Glomerular Co-Cultures Reveal Drawbacks of Currently Used Cell Culture Models
 , ,
, ,
Abstract
:1. Introduction
2. Results
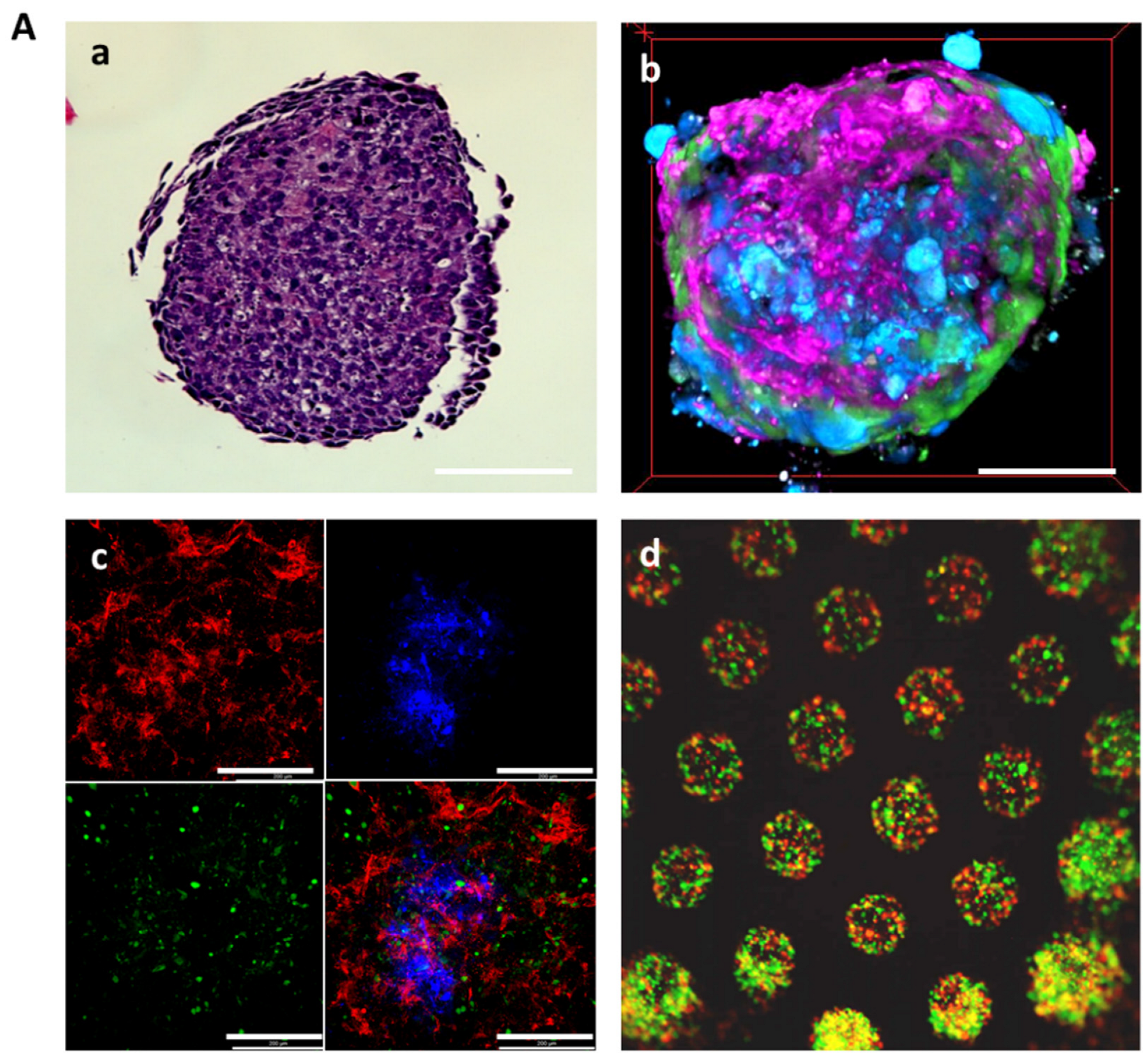
2.1. 3D Co-Culture of Human Glomerular Cell Types Improves Cell Survival
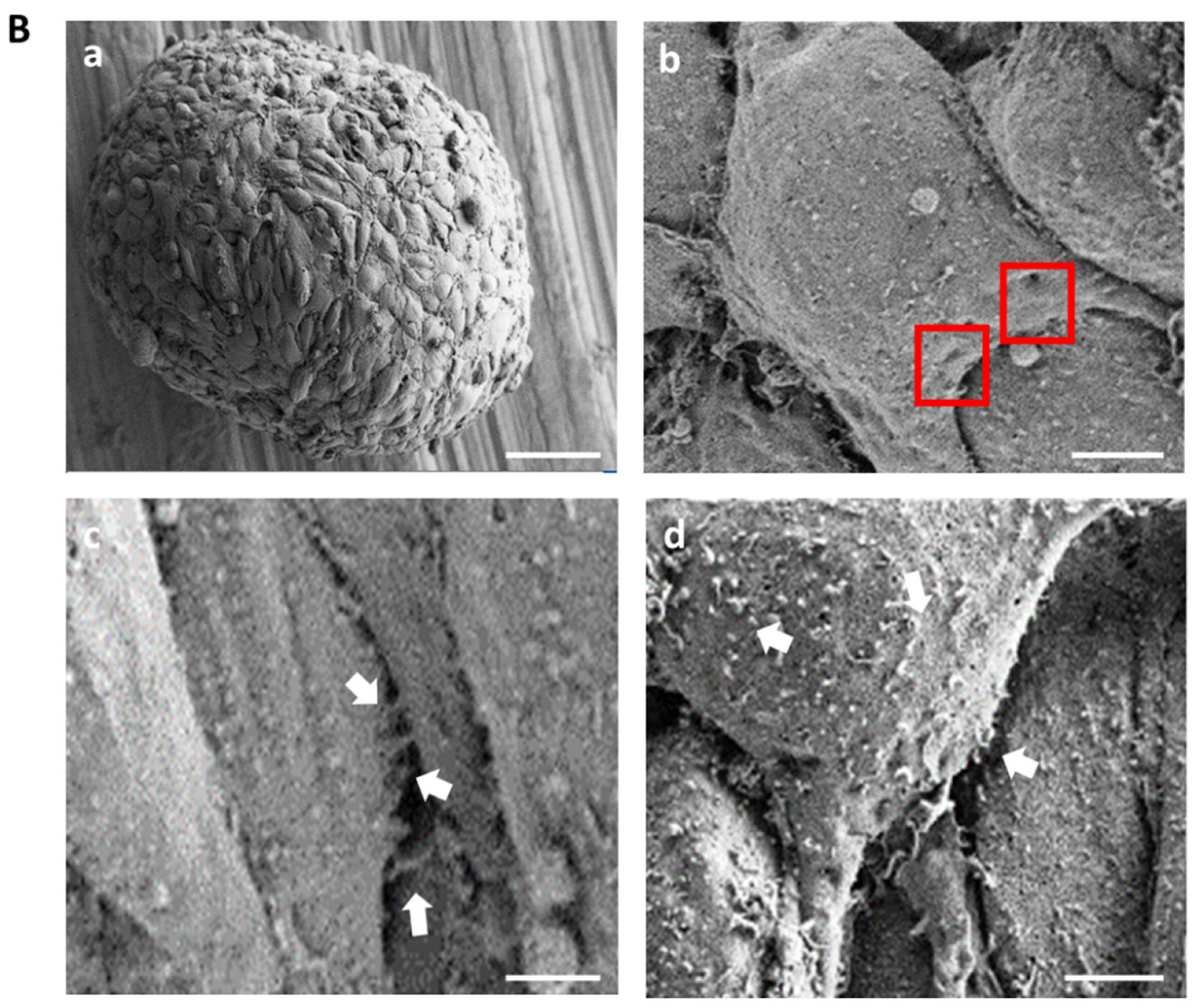
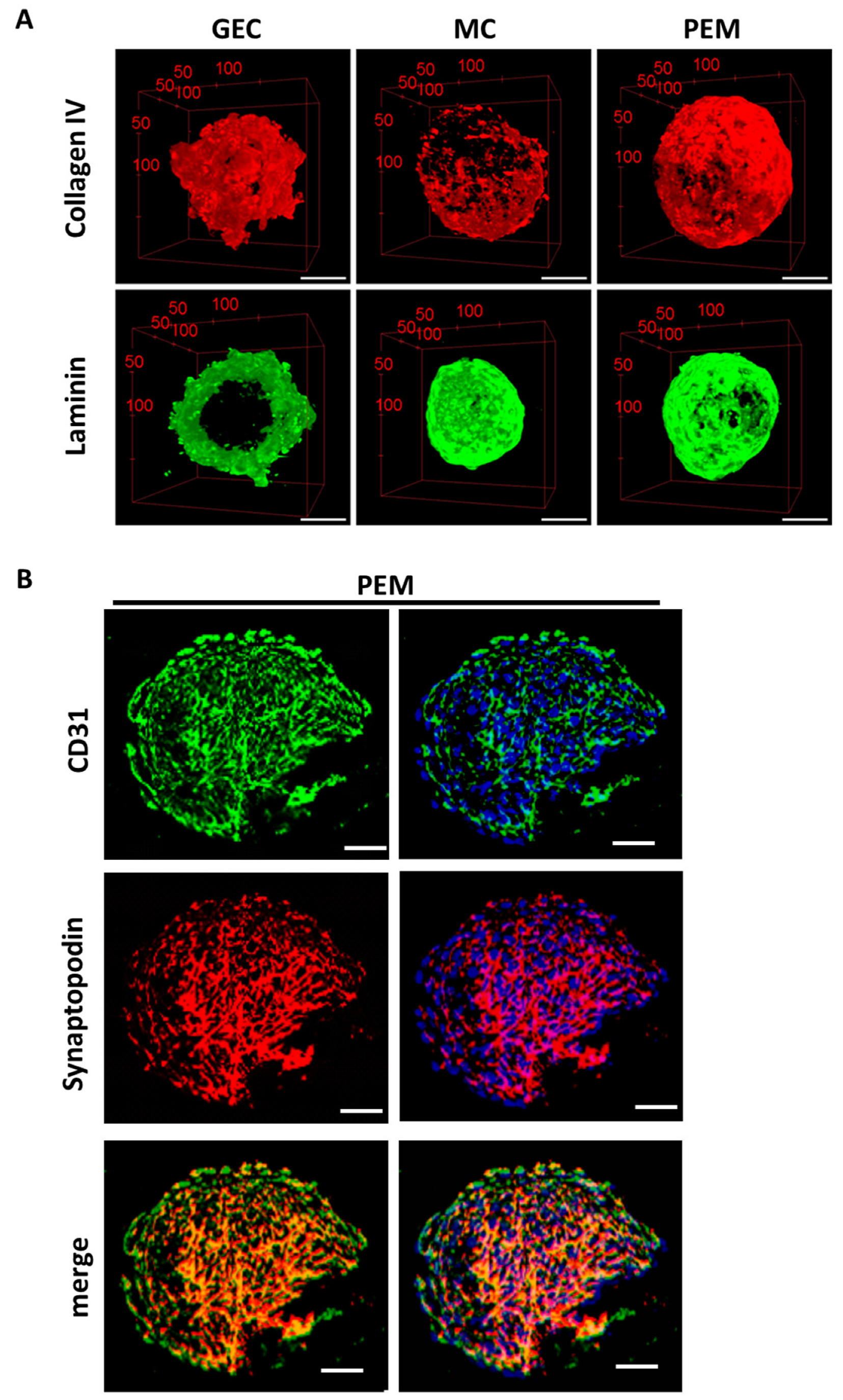
2.2. 3D Glomerular Co-Cultures Self-Arrange in a Specific Manner and Develop Fenestrae-like Structures of GEC and Interdigitating Cell Processes of Podocytes
2.3. Bulk-RNA Sequencing of 3D Glomerular Co-Culture Indicates Gene Expression Similar to In Vivo Condition
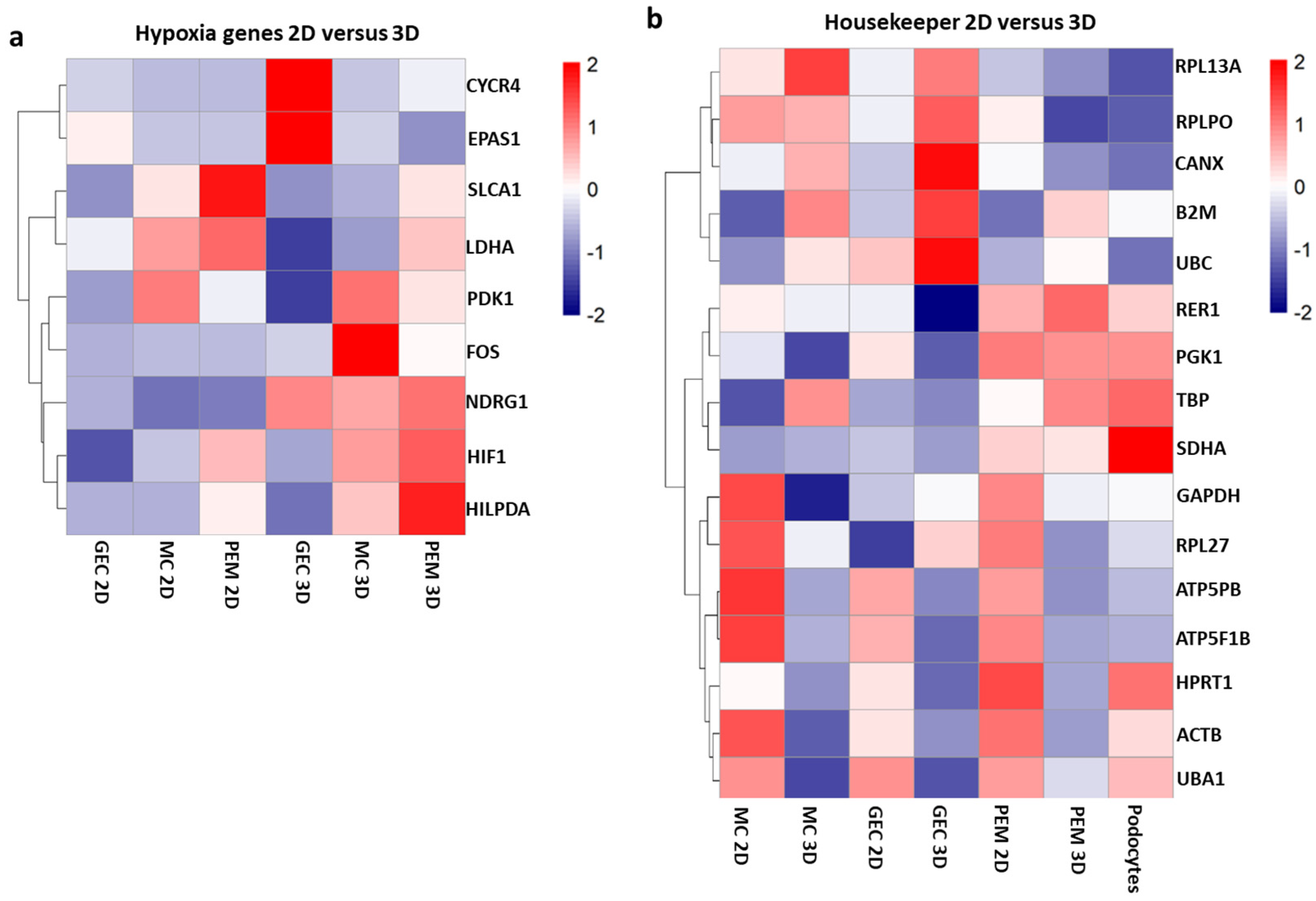
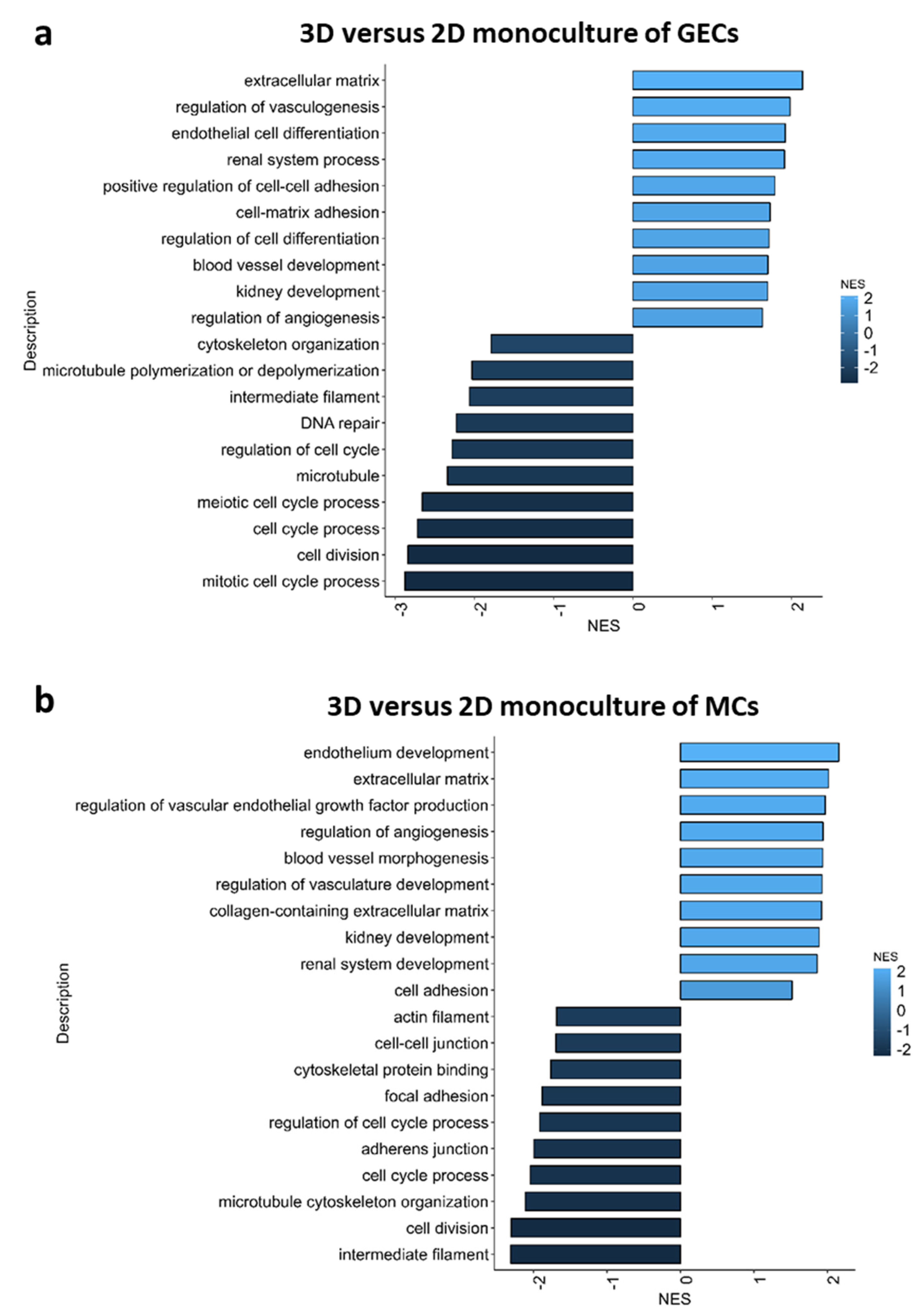
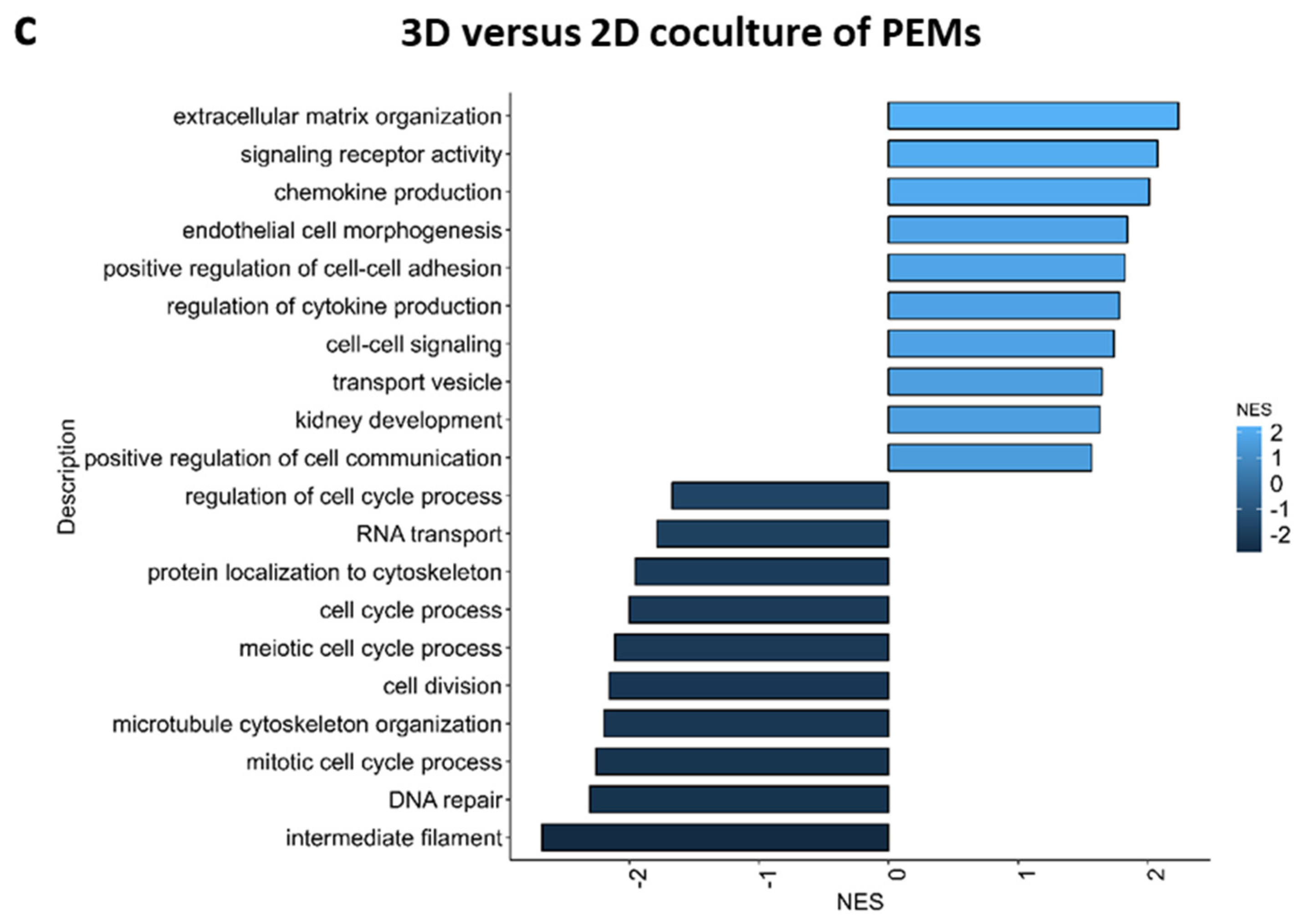
2.4. Pathway Analysis of 3D versus 2D Cultured Glomerular Cells
2.5. Validation of Bulk-RNA Sequencing Results by qPCR
2.6. Comparison of Protein Levels between 2D and 3D-Cultured Glomerular Cells
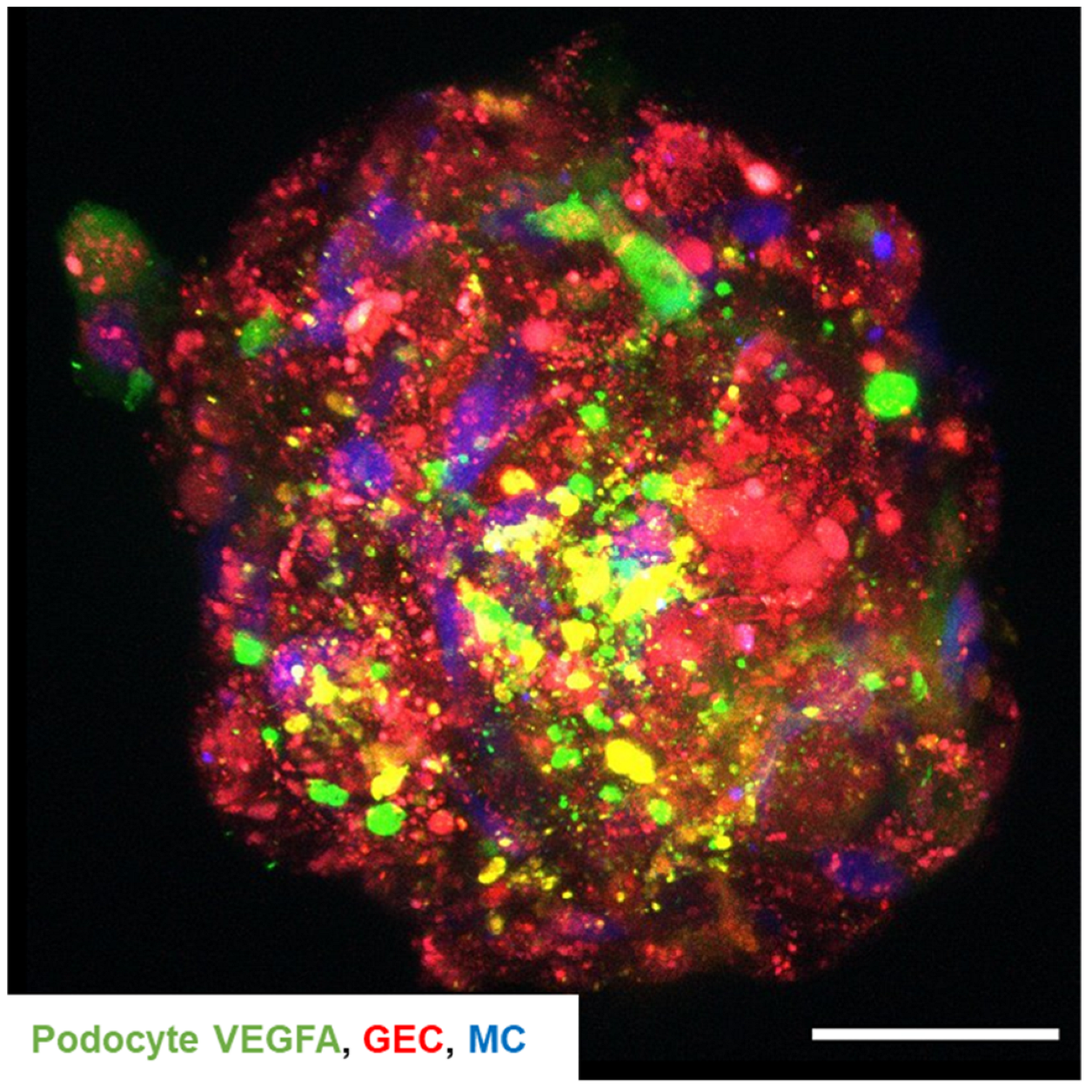
2.7. Cell–Cell Communication within the 3D Glomerular Spheroids
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Glomerular Endothelial Cell Reporter Cell Line
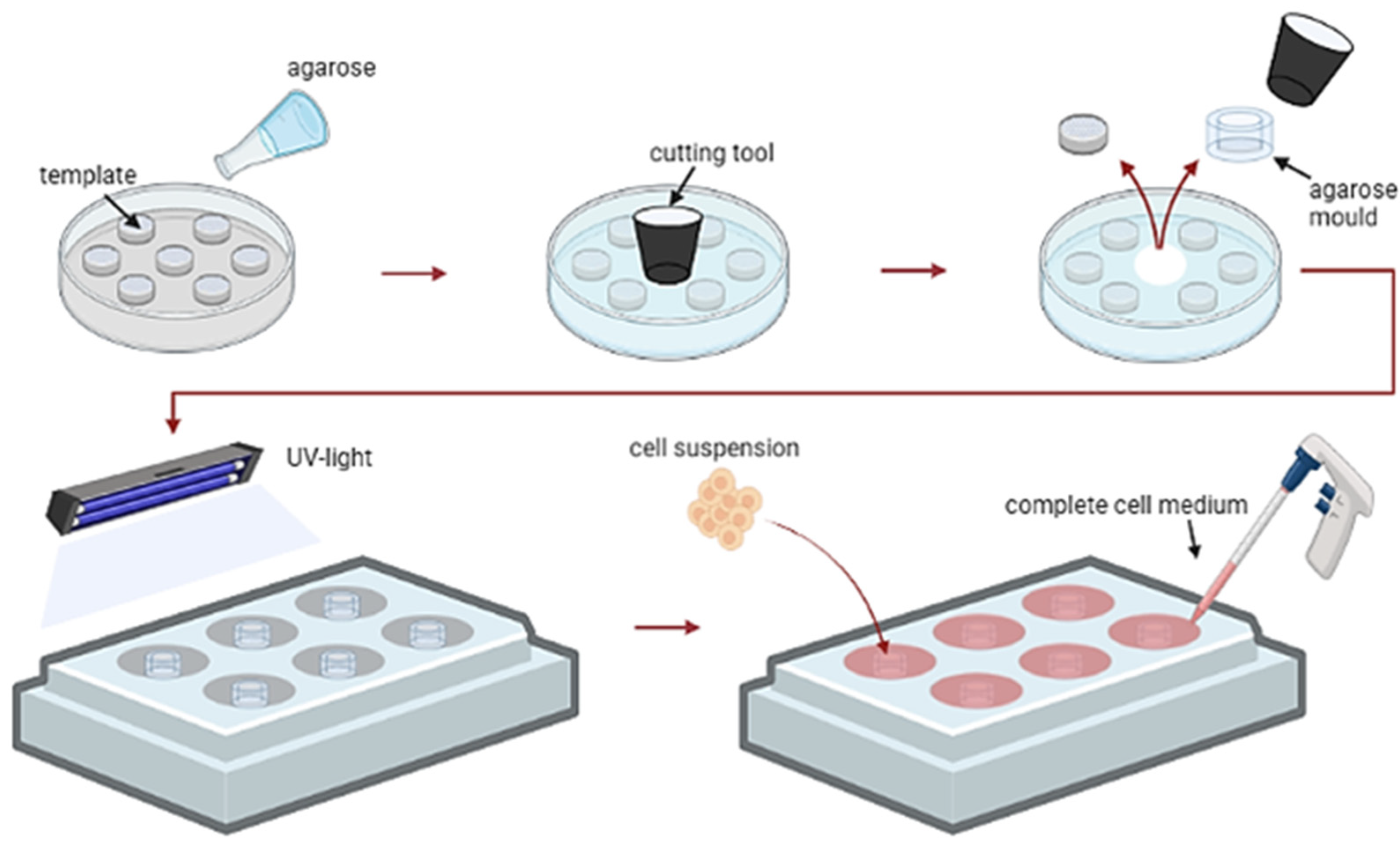
4.3. 3D Cell Culture
4.4. Live/Dead Assay
4.5. RNA Isolation
4.6. Bulk-RNA Sequencing
4.7. Immunocytochemistry-Staining
4.8. Labeling Living Cells with In Vivo Dyes
4.9. Multiphoton Microscopy
4.10. Design of Human VEGFA-GFP Plasmid and Transfection of Podocytes
4.11. Confocal Microscopy
4.12. Scanning Electron Microscopy
4.13. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Daehn, I.S.; Duffield, J.S. The glomerular filtration barrier: A structural target for novel kidney therapies. Nat. Rev. Drug Discov. 2021, 20, 770–788. [Google Scholar] [CrossRef]
- Pollak, M.R.; Quaggin, S.E.; Hoenig, M.P.; Dworkin, L.D. The glomerulus: The sphere of influence. Clin. J. Am. Soc. Nephrol. 2014, 9, 1461–1469. [Google Scholar] [CrossRef] [Green Version]
- Scott, R.P.; Quaggin, S.E. Review series: The cell biology of renal filtration. J. Cell Biol. 2015, 209, 199–210. [Google Scholar] [CrossRef]
- Müller-Deile, J.; Schröder, P.; Beverly-Staggs, L.; Hiss, R.; Fiedler, J.; Nyström, J.; Thum, T.; Haller, H.; Schiffer, M. Overexpression of preeclampsia induced microRNA-26a-5p leads to proteinuria in zebrafish. Sci. Rep. 2018, 8, 3621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller-Deile, J.; Dannenberg, J.; Schroder, P.; Lin, M.H.; Miner, J.H.; Chen, R.; Bräsen, J.H.; Thum, T.; Nyström, J.; Staggs, L.B.; et al. Podocytes regulate the glomerular basement membrane protein nephronectin by means of miR-378a-3p in glomerular diseases. Kidney Int. 2017, 92, 836–849. [Google Scholar] [CrossRef] [PubMed]
- Müller-Deile, J.; Sopel, N.; Ohs, A.; Rose, V.; Gröner, M.; Wrede, C.; Hegermann, J.; Daniel, C.; Amann, K.; Zahner, G.; et al. Glomerular Endothelial Cell-Derived microRNA-192 Regulates Nephronectin Expression in Idiopathic Membranous Glomerulonephritis. J. Am. Soc. Nephrol. 2021, 32, 2777–2794. [Google Scholar] [CrossRef] [PubMed]
- Gnudi, L.; Benedetti, S.; Woolf, A.S.; Long, D.A. Vascular growth factors play critical roles in kidney glomeruli. Clin. Sci. 2015, 129, 1225–1236. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.A.; O’Hare, M.J.; Reiser, J.; Coward, R.J.; Inward, C.D.; Farren, T.; Xing, C.Y.; Ni, L.; Mathieson, P.W.; Mundel, P. A conditionally immortalized human podocyte cell line demonstrating nephrin and podocin expression. J. Am. Soc. Nephrol. 2002, 13, 630–638. [Google Scholar] [CrossRef]
- Chittiprol, S.; Chen, P.; Petrovic-Djergovic, D.; Eichler, T.; Ransom, R.F. Marker expression, behaviors, and responses vary in different lines of conditionally immortalized cultured podocytes. Am. J. Physiol. Ren. Physiol. 2011, 301, F660–F671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valverde, M.G.; Mille, L.S.; Figler, K.P.; Cervantes, E.; Li, V.Y.; Bonventre, J.V.; Masereeuw, R.; Zhang, Y.S. Biomimetic models of the glomerulus. Nat. Rev. Nephrol. 2022, 18, 241–257. [Google Scholar] [CrossRef]
- Zhou, M.; Zhang, X.; Wen, X.; Wu, T.; Wang, W.; Yang, M.; Wang, J.; Fang, M.; Lin, B.; Lin, H. Development of a Functional Glomerulus at the Organ Level on a Chip to Mimic Hypertensive Nephropathy. Sci. Rep. 2016, 6, 31771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrosyan, A.; Cravedi, P.; Villani, V.; Angeletti, A.; Manrique, J.; Renieri, A.; De Filippo, R.E.; Perin, L.; Da Sacco, S. A glomerulus-on-a-chip to recapitulate the human glomerular filtration barrier. Nat. Commun. 2019, 10, 3656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roye, Y.; Musah, S. Isogenic Kidney Glomerulus Chip Engineered from Human Induced Pluripotent Stem Cells. J. Vis. Exp. 2022, 189, e63821. [Google Scholar] [CrossRef]
- Musah, S.; Mammoto, A.; Ferrante, T.C.; Jeanty, S.S.F.; Hirano-Kobayashi, M.; Mammoto, T.; Roberts, K.; Chung, S.; Novak, R.; Ingram, M.; et al. Mature induced-pluripotent-stem-cell-derived human podocytes reconstitute kidney glomerular-capillary-wall function on a chip. Nat. Biomed. Eng. 2017, 1, 0069. [Google Scholar] [CrossRef] [Green Version]
- Slater, S.C.; Beachley, V.; Hayes, T.; Zhang, D.; Welsh, G.I.; Saleem, M.A.; Mathieson, P.W.; Wen, X.; Su, B.; Satchell, S.C. An in vitro model of the glomerular capillary wall using electrospun collagen nanofibres in a bioartificial composite basement membrane. PLoS ONE 2011, 6, e20802. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Alfieri, C.M.; Morello, W.; Cellesi, F.; Armelloni, S.; Mattinzoli, D.; Montini, G.; Messa, P. Assessment of increased glomerular permeability associated with recurrent focal segmental glomerulosclerosis using an in vitro model of the glomerular filtration barrier. J. Nephrol. 2020, 33, 747–755. [Google Scholar] [CrossRef]
- Li, M.; Corbelli, A.; Watanabe, S.; Armelloni, S.; Ikehata, M.; Parazzi, V.; Pignatari, C.; Giardino, L.; Mattinzoli, D.; Lazzari, L.; et al. Three-dimensional podocyte-endothelial cell co-cultures: Assembly, validation, and application to drug testing and intercellular signaling studies. Eur. J. Pharm. Sci. 2016, 86, 1–12. [Google Scholar] [CrossRef]
- Hale, L.J.; Howden, S.E.; Phipson, B.; Lonsdale, A.; Er, P.X.; Ghobrial, I.; Hosawi, S.; Wilson, S.; Lawlor, K.T.; Khan, S.; et al. 3D organoid-derived human glomeruli for personalised podocyte disease modelling and drug screening. Nat. Commun. 2018, 9, 5167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Berg, C.W.; Ritsma, L.; Avramut, M.C.; Wiersma, L.E.; van den Berg, B.M.; Leuning, D.G.; Lievers, E.; Koning, M.; Vanslambrouck, J.M.; Koster, A.J.; et al. Renal Subcapsular Transplantation of PSC-Derived Kidney Organoids Induces Neo-vasculogenesis and Significant Glomerular and Tubular Maturation In Vivo. Stem Cell Rep. 2018, 10, 751–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takasato, M.; Er, P.X.; Chiu, H.S.; Maier, B.; Baillie, G.J.; Ferguson, C.; Parton, R.G.; Wolvetang, E.J.; Roost, M.S.; Chuva de Sousa Lopes, S.M.; et al. Kidney organoids from human iPS cells contain multiple lineages and model human nephrogenesis. Nature 2015, 526, 564–568. [Google Scholar] [CrossRef]
- Tuffin, J.; Burke, M.; Richardson, T.; Johnson, T.; Saleem, M.A.; Satchell, S.; Welsh, G.I.; Perriman, A. A Composite Hydrogel Scaffold Permits Self-Organization and Matrix Deposition by Cocultured Human Glomerular Cells. Adv. Healthc. Mater. 2019, 8, e1900698. [Google Scholar] [CrossRef] [PubMed]
- Ryma, M.; Genç, H.; Nadernezhad, A.; Paulus, I.; Schneidereit, D.; Friedrich, O.; Andelovic, K.; Lyer, S.; Alexiou, C.; Cicha, I.; et al. A Print-and-Fuse Strategy for Sacrificial Filaments Enables Biomimetically Structured Perfusable Microvascular Networks with Functional Endothelium Inside 3D Hydrogels. Adv. Mater. 2022, 34, e2200653. [Google Scholar] [CrossRef] [PubMed]
- Homan, K.A.; Kolesky, D.B.; Skylar-Scott, M.A.; Herrmann, J.; Obuobi, H.; Moisan, A.; Lewis, J.A. Bioprinting of 3D Convoluted Renal Proximal Tubules on Perfusable Chips. Sci. Rep. 2016, 6, 34845. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Kim, E.M.; Yamamoto, M.; Park, H.; Shin, H. Engineering Multi-Cellular Spheroids for Tissue Engineering and Regenerative Medicine. Adv. Healthc. Mater. 2020, 9, e2000608. [Google Scholar] [CrossRef]
- Potapova, I.A.; Gaudette, G.R.; Brink, P.R.; Robinson, R.B.; Rosen, M.R.; Cohen, I.S.; Doronin, S.V. Mesenchymal stem cells support migration, extracellular matrix invasion, proliferation, and survival of endothelial cells in vitro. Stem Cells 2007, 25, 1761–1768. [Google Scholar] [CrossRef]
- Sun, J.; He, H.; Xiong, Y.; Lu, S.; Shen, J.; Cheng, A.; Chang, W.-C.; Hou, M.-F.; Lancaster, J.M.; Kim, M.; et al. Fascin Protein Is Critical for Transforming Growth Factor β Protein-induced Invasion and Filopodia Formation in Spindle-shaped Tumor Cells. J. Biol. Chem. 2011, 286, 38865–38875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, H.Y.; Liu, B.H.; Sieber, M.; Hsu, S.H. Substrate-dependent gene regulation of self-assembled human MSC spheroids on chitosan membranes. BMC Genom. 2014, 15, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartosh, T.J.; Ylöstalo, J.H.; Mohammadipoor, A.; Bazhanov, N.; Coble, K.; Claypool, K.; Lee, R.H.; Choi, H.; Prockop, D.J. Aggregation of human mesenchymal stromal cells (MSCs) into 3D spheroids enhances their antiinflammatory properties. Proc. Natl. Acad. Sci. USA 2010, 107, 13724–13729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frith, J.E.; Thomson, B.; Genever, P.G. Dynamic three-dimensional culture methods enhance mesenchymal stem cell properties and increase therapeutic potential. Tissue Eng. Part C Methods 2010, 16, 735–749. [Google Scholar] [CrossRef]
- Tuffin, J.; Chesor, M.; Kuzmuk, V.; Johnson, T.; Satchell, S.C.; Welsh, G.I.; Saleem, M.A. GlomSpheres as a 3D co-culture spheroid model of the kidney glomerulus for rapid drug-screening. Commun. Biol. 2021, 4, 1351. [Google Scholar] [CrossRef]
- Yang, W.Y.; Chen, L.C.; Jhuang, Y.T.; Lin, Y.J.; Hung, P.Y.; Ko, Y.C.; Tsai, M.Y.; Lee, Y.W.; Hsu, L.W.; Yeh, C.K.; et al. Injection of hybrid 3D spheroids composed of podocytes, mesenchymal stem cells, and vascular endothelial cells into the renal cortex improves kidney function and replenishes glomerular podocytes. Bioeng. Transl. Med. 2021, 6, e10212. [Google Scholar] [CrossRef]
- St John, P.L.; Abrahamson, D.R. Glomerular endothelial cells and podocytes jointly synthesize laminin-1 and -11 chains. Kidney Int. 2001, 60, 1037–1046. [Google Scholar] [CrossRef] [Green Version]
- Yan, K.; Khoshnoodi, J.; Ruotsalainen, V.; Tryggvason, K. N-linked glycosylation is critical for the plasma membrane localization of nephrin. J. Am. Soc. Nephrol. 2002, 13, 1385–1389. [Google Scholar] [CrossRef] [Green Version]
- Nishinakamura, R. Human kidney organoids: Progress and remaining challenges. Nat. Rev. Nephrol. 2019, 15, 613–624. [Google Scholar] [CrossRef]
- Gupta, N.; Morizane, R. Kidney development to kidney organoids and back again. Semin. Cell Dev. Biol. 2022, 127, 68–76. [Google Scholar] [CrossRef]
- Chambers, B.E.; Weaver, N.E.; Wingert, R.A. The “3Ds” of Growing Kidney Organoids: Advances in Nephron Development, Disease Modeling, and Drug Screening. Cells 2023, 12, 549. [Google Scholar] [CrossRef]
- Leung, B.; Lesher-Perez, S.C.; Matsuoka, T.; Moraes, C.; Takayama, S. Media additives to promote spheroid circularity and compactness in hanging drop platform. Biomater. Sci. 2015, 3, 336–344. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, P.; Hellström, M.; Kalén, M.; Karlsson, L.; Pekny, M.; Pekna, M.; Soriano, P.; Betsholtz, C. Paracrine PDGF-B/PDGF-Rbeta signaling controls mesangial cell development in kidney glomeruli. Development 1998, 125, 3313–3322. [Google Scholar] [CrossRef] [PubMed]
- Eremina, V.; Jefferson, J.A.; Kowalewska, J.; Hochster, H.; Haas, M.; Weisstuch, J.; Richardson, C.; Kopp, J.B.; Kabir, M.G.; Backx, P.H.; et al. VEGF inhibition and renal thrombotic microangiopathy. N. Engl. J. Med. 2008, 358, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Miner, J.H. Organogenesis of the kidney glomerulus: Focus on the glomerular basement membrane. Organogenesis 2011, 7, 75–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miner, J.H. Developmental biology of glomerular basement membrane components. Curr. Opin. Nephrol. Hypertens. 1998, 7, 13–19. [Google Scholar] [CrossRef]
- Abrahamson, D.R.; Hudson, B.G.; Stroganova, L.; Borza, D.B.; St John, P.L. Cellular origins of type IV collagen networks in developing glomeruli. J. Am. Soc. Nephrol. 2009, 20, 1471–1479. [Google Scholar] [CrossRef] [Green Version]
- Kopp, J.B.; Anders, H.J.; Susztak, K.; Podestà, M.A.; Remuzzi, G.; Hildebrandt, F.; Romagnani, P. Podocytopathies. Nat. Rev. Dis. Prim. 2020, 6, 68. [Google Scholar] [CrossRef]
- Schloñdorff, D.; Banas, B. The mesangial cell revisited: No cell is an island. J. Am. Soc. Nephrol. 2009, 20, 1179–1187. [Google Scholar]
- Byron, A.; Randles, M.J.; Humphries, J.D.; Mironov, A.; Hamidi, H.; Harris, S.; Mathieson, P.W.; Saleem, M.A.; Satchell, S.C.; Zent, R.; et al. Glomerular Cell Cross-Talk Influences Composition and Assembly of Extracellular Matrix. J. Am. Soc. Nephrol. 2014, 25, 953–966. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.-D.; Xie, K.-F.; Luo, X.-J. Hypoxia-induced elevated NDRG1 mediates apoptosis through reprograming mitochondrial fission in HCC. Gene 2020, 741, 144552. [Google Scholar] [CrossRef] [PubMed]
- Sibold, S.; Roh, V.; Keogh, A.; Studer, P.; Tiffon, C.; Angst, E.; Vorburger, S.A.; Weimann, R.; Candinas, D.; Stroka, D. Hypoxia increases cytoplasmic expression of NDRG1, but is insufficient for its membrane localization in human hepatocellular carcinoma. FEBS Lett. 2007, 581, 989–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radonić, A.; Thulke, S.; Mackay, I.M.; Landt, O.; Siegert, W.; Nitsche, A. Guideline to reference gene selection for quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2004, 313, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Deindl, E.; Boengler, K.; van Royen, N.; Schaper, W. Differential expression of GAPDH and beta3-actin in growing collateral arteries. Mol. Cell Biochem. 2002, 236, 139–146. [Google Scholar] [CrossRef]
- Silver, N.; Cotroneo, E.; Proctor, G.; Osailan, S.; Paterson, K.L.; Carpenter, G.H. Selection of housekeeping genes for gene expression studies in the adult rat submandibular gland under normal, inflamed, atrophic and regenerative states. BMC Mol. Biol. 2008, 9, 64. [Google Scholar] [CrossRef] [Green Version]
- Leedale, J.A.; Lucendo-Villarin, B.; Meseguer-Ripolles, J.; Kasarinaite, A.; Webb, S.D.; Hay, D.C. Mathematical modelling of oxygen gradients in stem cell-derived liver tissue. PLoS ONE 2021, 16, e0244070. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, T.; Byun, H.; Lee, J.; Madhurakat Perikamana, S.K.; Shin, Y.M.; Kim, E.M.; Shin, H. Stem cell spheroids incorporating fibers coated with adenosine and polydopamine as a modular building blocks for bone tissue engineering. Biomaterials 2020, 230, 119652. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Encoding for | Sequence (5′ -> 3′) |
|---|---|---|
| hHPRT | Hypoxanthine phosphoribosyltransferase 1 | GACCAGTCAACAGGGGACAT AACACTTCGTGGGGTCCTTTTC |
| hGAPDH | Glyceraldehyde-3-phosphate dehydrogenase | CAAGATCATCAGCAATGCCTCC ATGATGTTCTGGAGAGCCCC |
| hRPS18 | RPS18 rRNA | GTTGATTAAGTCCCTGCCCTTTGT CGATCCGAGGGCCTCACTA |
| hWT1 | Wilms tumor protein | TTATTGCAGCCTGGGTAAGC TCAGAGGCATTCAGGATGTG |
| hSYNPO | Synaptopodin | TAAGCAACC TTCTGGGCTAAAGCTAAC |
| hNPHS1 | Nephrin | TCACATCTCCATGTCCAACC GGGCGGGATATTTTACGTTC |
| hVEGFA | Vascular endothelial growth factor A | CAACAAATGTGAATGCAGACCAAA CCCTTTCCCTTTCCTCGAACT |
| hPECAM1 | CD31 | TGGAAAGCAGATACTCTAGAACGG GGGATGTGCATCTGGCCTT |
| hKDR | Kinase insert domain receptor, VEGFR2 | TGGTTGTGTATGTCCCACCC GGAGGAATGGCATAGACCGT |
| hFLT1 | Fms-related receptor tyrosine kinase 1, VEGFR1 | TGTGAAAATGCTGAAAGAGGGGG AGATGGTGGCCAATGTGGGT |
| hPDGFB | Platelet derived growth factor subunit B | GTTTATCATGGGCCTCGGGGA TCATCAAAGGAGCGGATCGAG |
| hLAMA5 | Laminin 5 | GGAGAACGGAGAGATCGTGG CAGCGGCGAGTAGGAGAAAT |
| hCOL4A1 | Collagen IVα1 | GTCTGGCTGCTGCTGCT CACAGCCACCCTTCGCA |
| hCOL4A4 | Collagen 4α4 | GCTTCTTGCACTCACAACGG GCTCCTGTAACAGCCAACCA |
| hCOL4A5 | Collagen 4α5 | GGCCCCAAGGTCCTCCT TCCACTGGGTCCTTTCATGC |
| LDHA1 | Lactat dehydrogenase A | CATAGCTGTTCCACTTAAGGCCC TGCCATATTGGACTTGGAACC |
| hHIG2 | Hypoxia-inducible gene 2 | AACACATGCTTCATGGCTGAAAGG TCTGCGCTGGTGCTTAGTAACC |
| hGLUT 1 | Glucose transporter 1 | ACCATTGGCTCCGGTATCG GCTCGCTCCACCACAAACA |
| hMIF | Macrophage migration inhibitory factor | CGGGTTCCTCTCCGAGCT CCGATCTTGCCGATGCTGT |
| hHIF1a | Hypoxia-Inducible Factor 1-Alpha | GAAGTGGCAACTGATGAGCA GCGCGAACGACAAGAAA |
| hHIF2a | Hypoxia-Inducible Factor 2-Alpha | GTCTGAACGTCTCAAAGGGC CTTCTCCTTCCTCCTCTCCG |
| hACTB | beta actin | ACCGAGCGTGGCTACAGCTTCACC AGCACCCGTGGCCATCTCTTTCTCG |
| Antigen (Clone) | Host | Company, Cat. No. | Dilution |
|---|---|---|---|
| Collagen IV | rabbit | Abcam, Cambridge, UK, ab6586 | 1:100 |
| Laminin 1/2 | rabbit | Novus Bio, Littleton, CO, USA, NB300-144SS | 1:100 |
| CD31 | mouse | ThermoFisher Scientific, Waltham, MA, USA, MA5-13188 | 1:100 |
| Synaptopodin | rabbit | Proteintech, Rosemont, IL, USA, 21064-1-AP | 1:100 |
| PEM (Figure 3) | PEM (Figure 8) | GEC (Figure 8) | MC (Figure 8) | ||
|---|---|---|---|---|---|
| Dichroic mirrors | 495 nm | 560 nm | 560 nm | 560 nm | |
| 560 nm | n.a. | n.a. | n.a. | ||
| Fluorophores | tdTomato | 572/35 nm | n.a. | n.a. | n.a. |
| CFSE | 525/50 nm | n.a. | n.a. | n.a. | |
| E450 | 450/30 nm | n.a. | n.a. | n.a. | |
| Alexa Fluor 647 | n.a. | 675/67 nm | 675/67 nm | 675/67 nm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rederer, A.; Rose, V.; Krüger, R.; Schmittutz, L.; Swierzy, I.; Fischer, L.; Thievessen, I.; Bauer, J.; Friedrich, O.; Schiffer, M.; et al. Partner, Neighbor, Housekeeper and Dimension: 3D versus 2D Glomerular Co-Cultures Reveal Drawbacks of Currently Used Cell Culture Models. Int. J. Mol. Sci. 2023, 24, 10384. https://doi.org/10.3390/ijms241210384
Rederer A, Rose V, Krüger R, Schmittutz L, Swierzy I, Fischer L, Thievessen I, Bauer J, Friedrich O, Schiffer M, et al. Partner, Neighbor, Housekeeper and Dimension: 3D versus 2D Glomerular Co-Cultures Reveal Drawbacks of Currently Used Cell Culture Models. International Journal of Molecular Sciences. 2023; 24(12):10384. https://doi.org/10.3390/ijms241210384
Chicago/Turabian StyleRederer, Anna, Victoria Rose, René Krüger, Linda Schmittutz, Izabela Swierzy, Lena Fischer, Ingo Thievessen, Julian Bauer, Oliver Friedrich, Mario Schiffer, and et al. 2023. "Partner, Neighbor, Housekeeper and Dimension: 3D versus 2D Glomerular Co-Cultures Reveal Drawbacks of Currently Used Cell Culture Models" International Journal of Molecular Sciences 24, no. 12: 10384. https://doi.org/10.3390/ijms241210384