
Seed Priming with MeJa Prevents Salt-Induced Growth Inhibition and Oxidative Damage in Sorghum bicolor by Inducing the Expression of Jasmonic Acid Biosynthesis Genes
 , ,
, ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
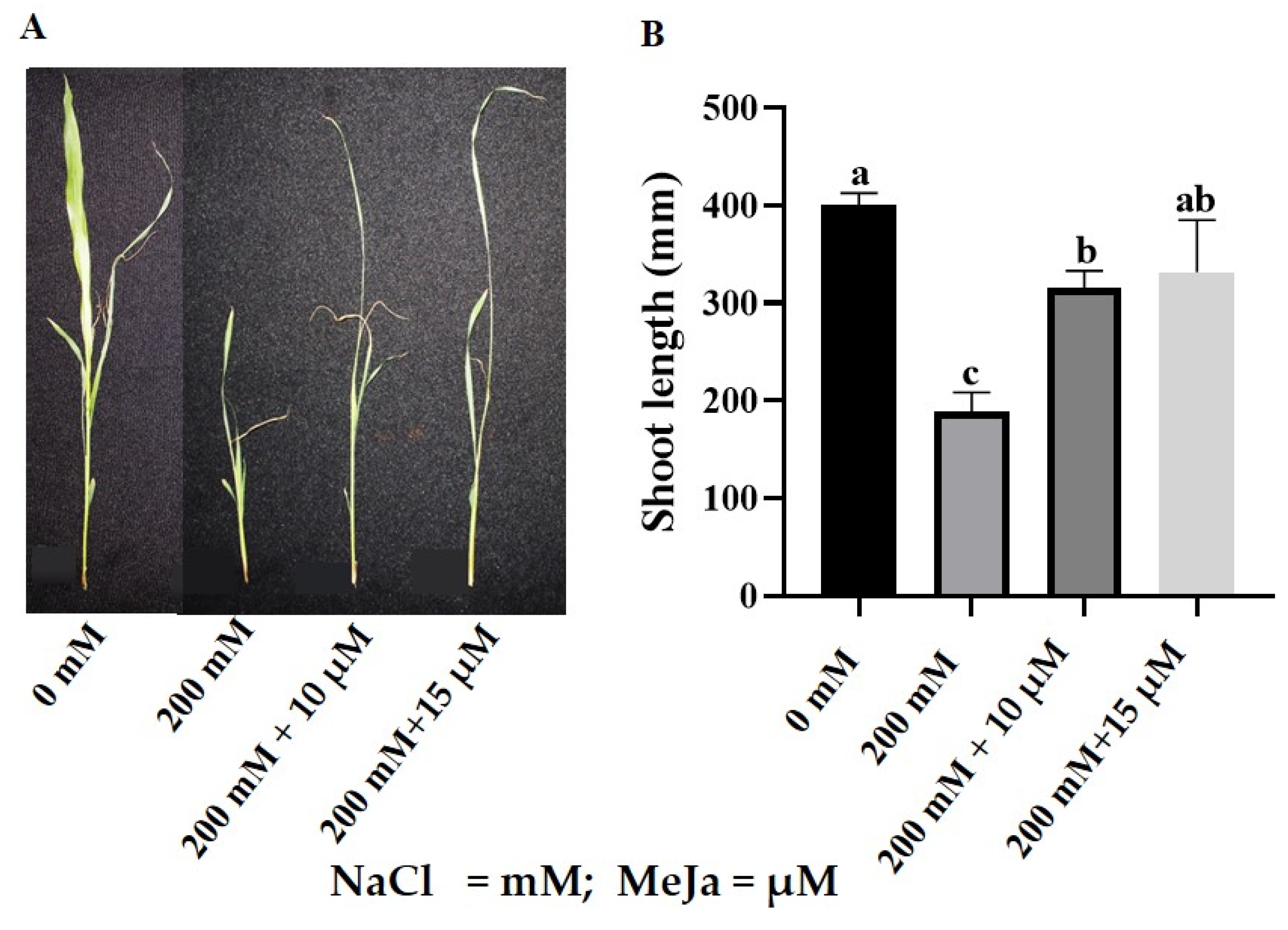
2.1. Morpho-Physiological Response of S. bicolor to MeJa under Salt Stress
2.1.1. Growth and Biomass
2.1.2. Photosynthetic Pigments
2.2. Biochemical Response of S. bicolor to MeJa under Salt Stress
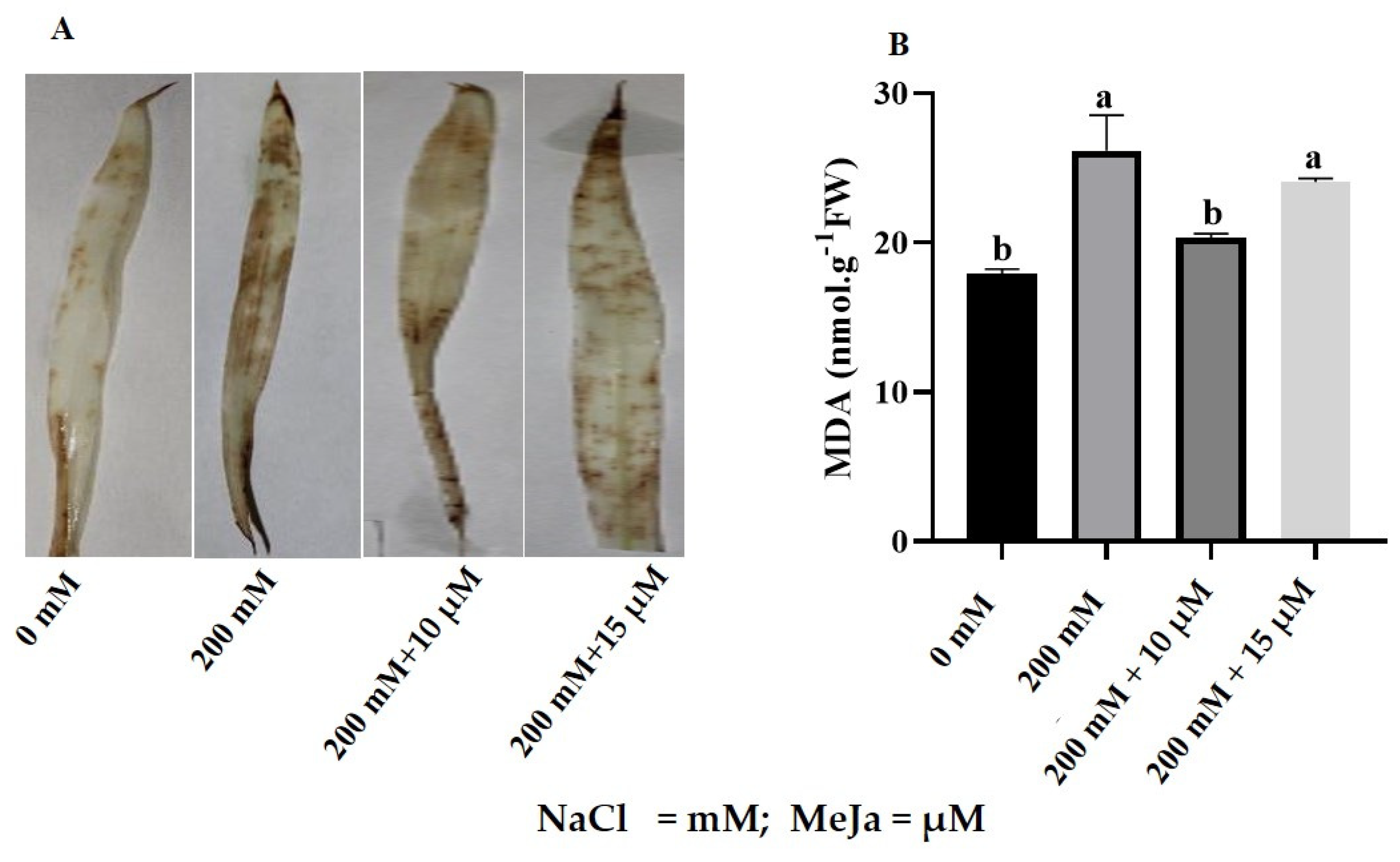
2.2.1. ROS Production
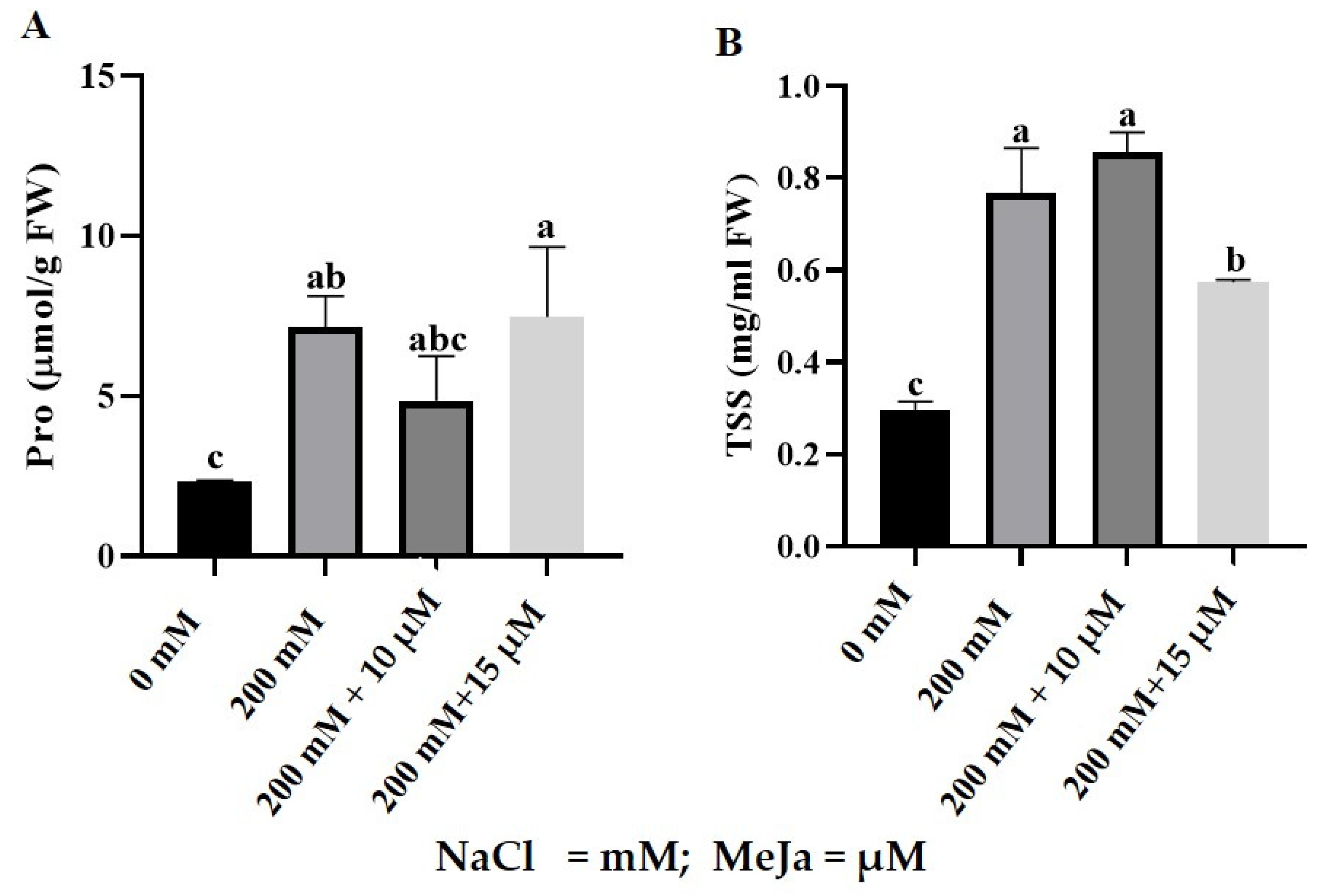
2.2.2. Proline and Total Soluble Sugars Content
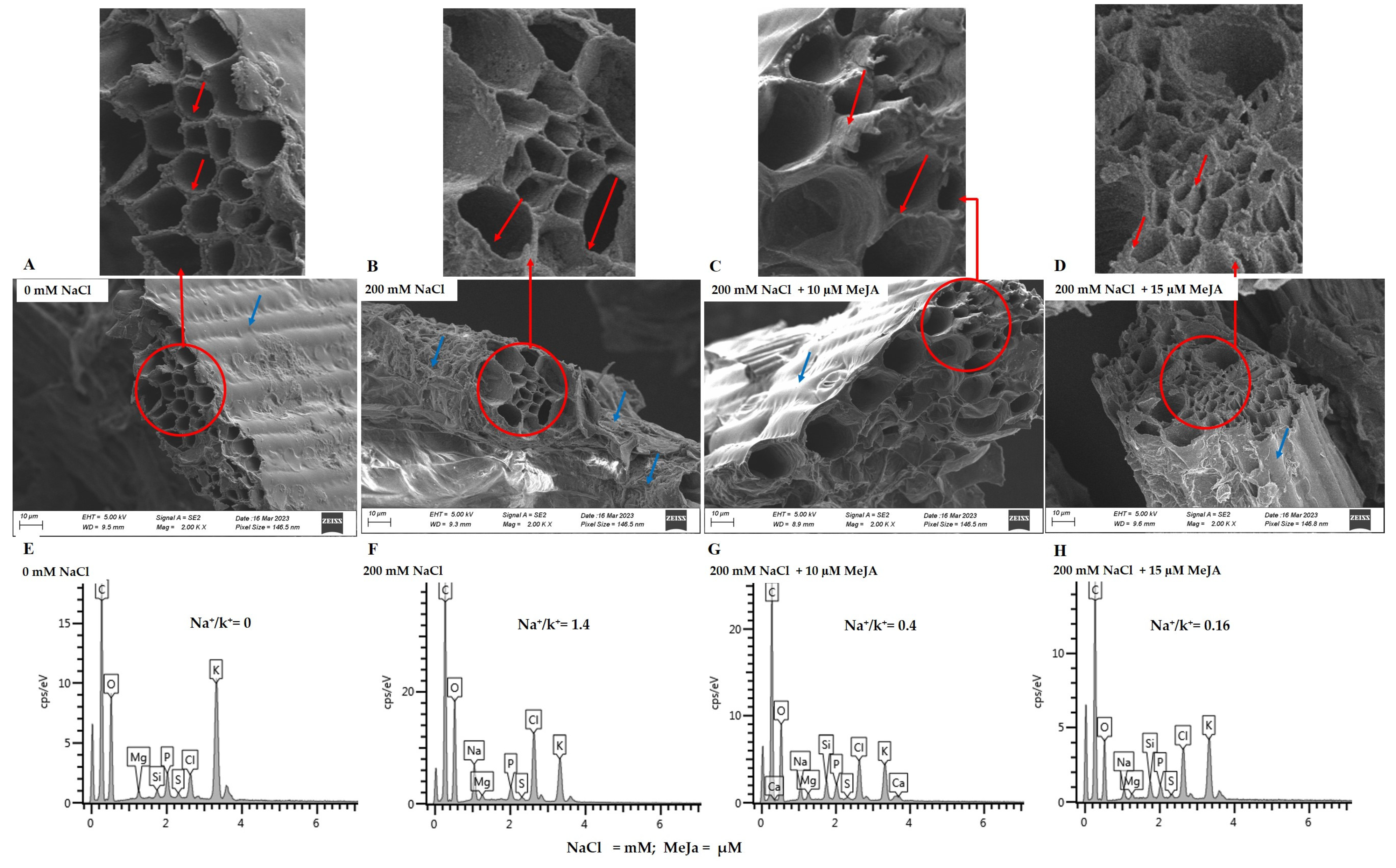
2.2.3. Anatomic Structure and Element Analysis
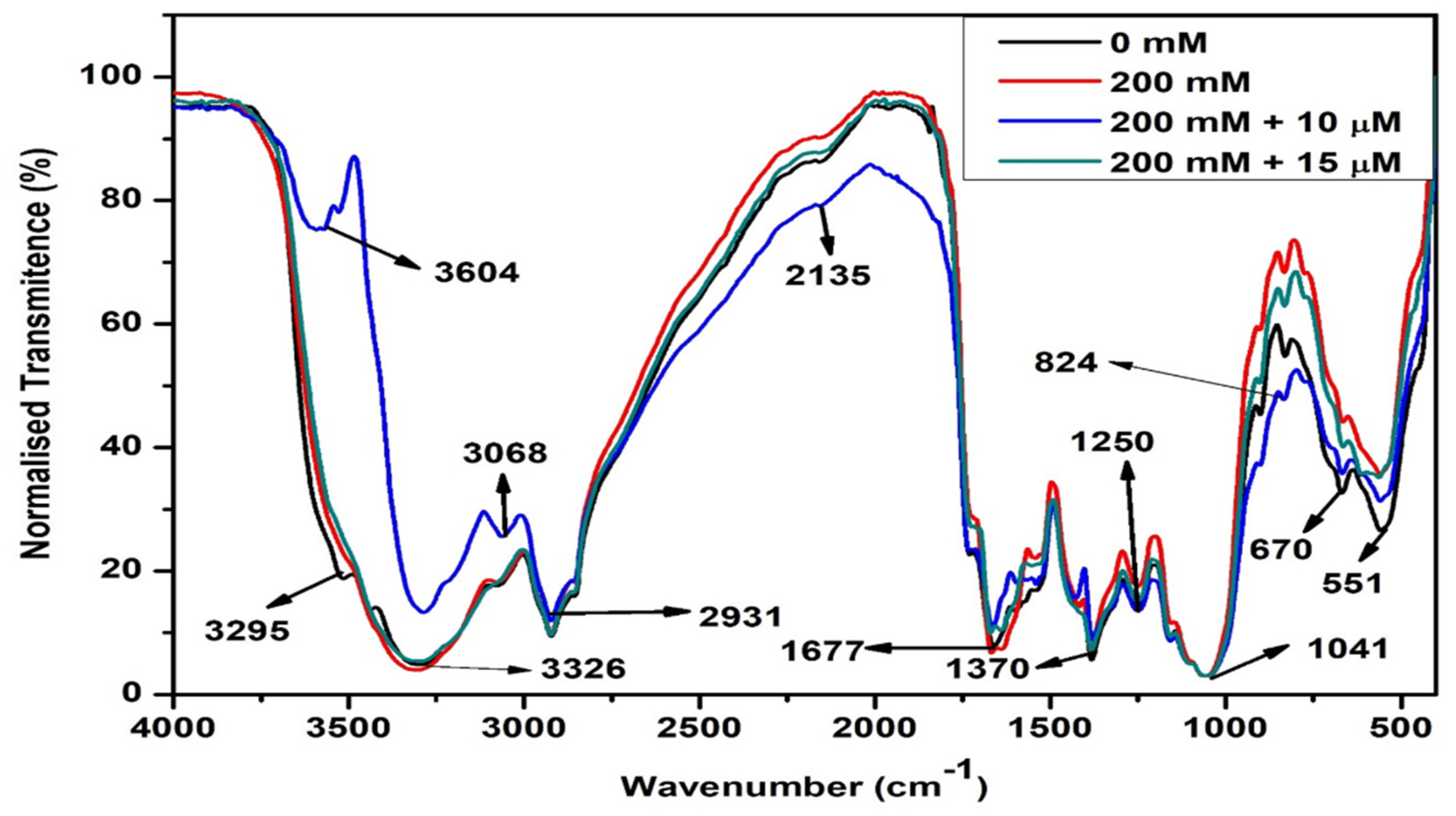
2.2.4. Changes in Biomolecules
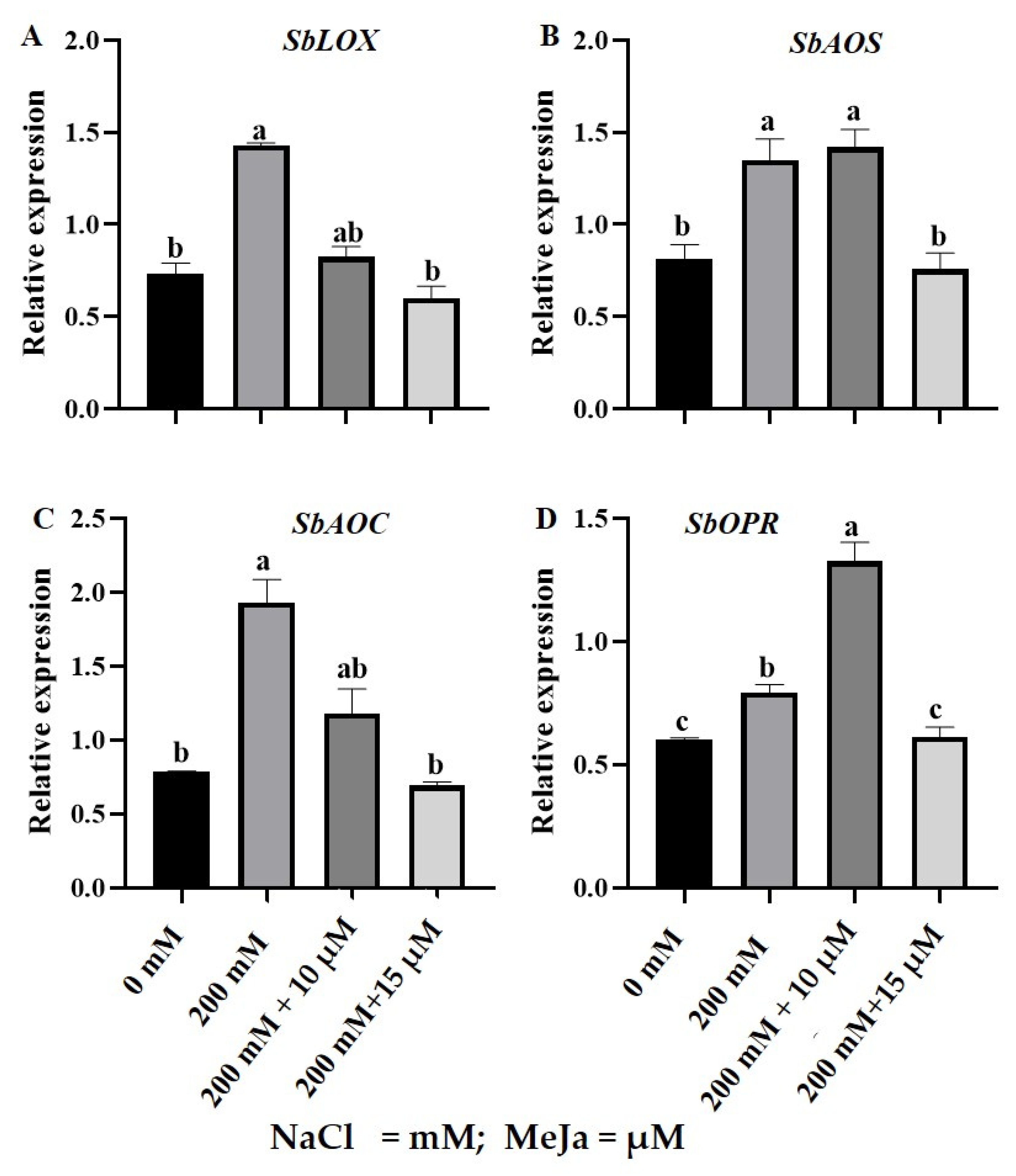
2.3. Gene Expression Analysis of Jasmonic Acid Biosynthesis Pathway Genes in S. bicolor under Salt Stress in Response to MeJa
3. Discussion
4. Materials and Methods
4.1. Sorghum Germination and Growth Condition
4.2. Treatment Application
4.3. Histochemical Detection of H2O2
4.4. Lipid Peroxidation
4.5. Chlorophyll Content
4.6. Proline Content
4.7. Total Soluble Sugars
4.8. Anatomic Analysis Using Scanning Electron Microscopy
4.9. Fourier-Transform Infrared (FTIR) Spectroscopic Analysis of Biomolecules
4.10. RNA Extraction and cDNA Preparation
4.11. Quantitative Real-Time Polymerase Chain Reaction
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, Y.; Ambrose, R.P.K. Structural characteristics of sorghum kernel: Effects of temperature. Int. J. Food Prop. 2017, 20, 2630–2638. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, M.; Kale, H.; Kardes, Y.M.; Karaman, K.; Yılmaz, M.F.; Temizgül, R.; Akar, T. Characterization of local sorghum (Sorghum bicolor L.) population grains in terms of nutritional properties and evaluation by GT biplot approach. Starch 2020, 72, 1900232. [Google Scholar] [CrossRef]
- de Morais Cardoso, L.; Pinheiro, S.S.; Martino, H.S.; Pinheiro-Sant′Ana, H.M. Sorghum (Sorghum bicolor L.): Nutrients, bioactive compounds, and potential impact on human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 372–390. [Google Scholar] [CrossRef]
- Shen, S.; Huang, R.; Li, C.; Wu, W.; Chen, H.; Shi, J.; Chen, S.; Ye, X. Phenolic compositions and antioxidant activities differ significantly among sorghum grains with different applications. Molecules 2018, 23, 1203. [Google Scholar] [CrossRef] [Green Version]
- Mullet, J.E.; Klein, R.R.; Klein, P.E. Sorghum bicolor—An important species for comparative grass genomics and a source of beneficial genes for agriculture. Curr. Opin. Plant Biol. 2002, 5, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.H.; Bowers, J.E.; Bruggmann, R.; Dubchak, I.; Grimwood, J.; Gundlach, H.; Haberer, G.; Hellsten, U.; Mitros, T.; Poliakov, A.; et al. The Sorghum bicolor genome and the diversification of grasses. Nature 2009, 457, 551–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, M. The Feeding of Nations: Redefining Food Security for the 21st Century; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Singh, R.K.; Joshi, P.K.; Sinha, V.S.P.; Kumar, M. Indicator based assessment of food security in SAARC nations under the influence of climate change scenarios. Futur. Foods 2022, 5, 100122. [Google Scholar] [CrossRef]
- Hinojosa, L.; González, J.A.; Barrios-Masias, F.H.; Fuentes, F.; Murphy, K.M. Quinoa abiotic stress responses: A review. Plants 2018, 7, 106. [Google Scholar] [CrossRef] [Green Version]
- He, M.; He, C.-Q.; Ding, N.-Z. Abiotic stresses: General defenses of land plants and chances for engineering multistress tolerance. Front. Plant Sci. 2018, 9, 1771. [Google Scholar] [CrossRef] [Green Version]
- Du, F.; Shi, H.; Zhang, X.; Xu, X. Responses of reactive oxygen scavenging enzymes, proline and malondialdehyde to water deficits among six secondary successional seral species in Loess Plateau. PLoS ONE 2014, 9, e98872. [Google Scholar] [CrossRef]
- Ndou, N.; Rakgotho, T.; Nkuna, M.; Doumbia, I.Z.; Mulaudzi, T.; Ajayi, R.F. Green Synthesis of Iron Oxide (Hematite) Nanoparticles and Their Influence on Sorghum bicolor Growth under Drought Stress. Plants 2023, 12, 1425. [Google Scholar] [CrossRef]
- Kong, W.; Liu, F.; Zhang, C.; Zhang, J.; Feng, H. Non-destructive determination of Malondialdehyde (MDA) distribution in oilseed rape leaves by laboratory scale NIR hyperspectral imaging. Sci. Rep. 2016, 6, 35393. [Google Scholar] [CrossRef] [PubMed]
- Safdar, H.; Amin, A.; Shafiq, Y.; Ali, A.; Yasin, R.; Shoukat, A.; Ul Hussan, M.; Sarwar, M.I. A review: Impact of salinity on plant growth. Nat. Sci. 2019, 17, 34–40. [Google Scholar]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef] [Green Version]
- Mulaudzi, T.; Hendricks, K.; Mabiya, T.; Muthevhuli, M.; Ajayi, R.F.; Mayedwa, N.; Gehring, C.; Iwuoha, E. Calcium improves germination and growth of Sorghum bicolor seedlings under salt stress. Plants 2020, 9, 730. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.H.; Kumar, S.A.; Ramesh, K.; Reddy, P.S.; Nagaraju, M.; Prakash, A.B.; Shah, T.; Henderson, A.; Srivastava, R.K.; Rajasheker, G.; et al. Genome-wide identification and analysis of Arabidopsis sodium proton antiporter (NHX) and human sodium proton exchanger (NHE) homologs in sorghum bicolor. Genes 2018, 9, 236. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, P.; Abass Ahanger, M.; Nasser Alyemeni, M.; Wijaya, L.; Alam, P.; Ashraf, M. Mitigation of sodium chloride toxicity in Solanum lycopersicum L. by supplementation of jasmonic acid and nitric oxide. J. Plant Interact. 2018, 13, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Yin, F.; Liu, X.; Cao, B.; Xu, K. Low pH altered salt stress in antioxidant metabolism and nitrogen assimilation in ginger (Zingiber officinale) seedlings. Physiol. Plant. 2020, 168, 648–659. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, L.; Lu, B.; Ma, T.; Jiang, D.; Li, J.; Zhang, K.; Sun, H.; Zhang, Y.; Bai, Z.; et al. Exogenous melatonin promotes seed germination and osmotic regulation under salt stress in cotton (Gossypium hirsutum L.). PLoS ONE 2020, 15, e0228241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, C.; Prakash, D. Enhancement of active constituents of medicinal plants through the use of microbes. In Plant Biotic Interactions; Springer: Berlin/Heidelberg, Germany, 2019; pp. 227–241. [Google Scholar]
- Niu, Y.; Hu, T.; Zhou, Y.; Hasi, A. Isolation and characterization of two Medicago falcate AP2/EREBP family transcription factor cDNA, MfDREB1 and MfDREB1s. Plant Physiol Biochem. 2010, 48, 971–976. [Google Scholar] [CrossRef]
- Hdira, S.; Haddoudi, L.; Hanana, M.; Romero, I.; Mahjoub, A.; Ben Jouira, H.; Ludidi, N.; Sanchez-Ballesta, M.T.; Abdelly, C.; Badri, M. Morpho-Physiological, Biochemical, and Genetic Responses to Salinity in Medicago truncatula. Plants 2021, 10, 808. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Bhardwaj, R.; Zheng, B. Phytohormones regulate accumulation of osmolytes under abiotic stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- Hannachi, S.; Van Labeke, M.-C. Salt stress affects germination, seedling growth and physiological responses differentially in eggplant cultivars (Solanum melongena L.). Sci. Hortic. 2018, 228, 56–65. [Google Scholar] [CrossRef]
- Liu, X.; Hou, X. Antagonistic regulation of ABA and GA in metabolism and signaling pathways. Front. Plant Sci. 2018, 9, 251. [Google Scholar] [CrossRef] [Green Version]
- Ku, Y.-S.; Sintaha, M.; Cheung, M.-Y.; Lam, H.-M. Plant hormone signaling crosstalks between biotic and abiotic stress responses. Int. J. Mol. Sci. 2018, 19, 3206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabagh, A.E.; Mbarki, S.; Hossain, A.; Iqbal, M.A.; Islam, M.S.; Raza, A.; Llanes, A.; Reginato, M.; Rahman, M.A.; Mahboob, W.; et al. Potential role of plant growth regulators in administering crucial processes against abiotic stresses. Front. Agron. 2021, 3, 648694. [Google Scholar] [CrossRef]
- Xu, L.; Wu, C.; Oelmüller, R.; Zhang, W. Role of phytohormones in Piriformospora indica-induced growth promotion and stress tolerance in plants: More questions than answers. Front. Microbiol. 2018, 9, 1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trifunović-Momčilov, M.; Motyka, V.; Dobrev, P.I.; Marković, M.; Milošević, S.; Jevremović, S.; Dragićević, I.; Subotić, A. Phytohormone profiles in non-transformed and AtCKX transgenic centaury (Centaurium erythraea Rafn) shoots and roots in response to salinity stress in vitro. Sci. Rep. 2021, 11, 21471. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Yan, S. MeJA is more effective than JA in inducing defense responses in Larix olgensis. Arthropod-Plant Interactions 2018, 12, 49–56. [Google Scholar] [CrossRef]
- Sujeeth, N.; Mehterov, N.; Gupta, S.; Qureshi, M.K.; Fischer, A.; Proost, S.; Omidbakhshfard, M.A.; Obata, T.; Benina, M.; Staykov, N.; et al. A novel seed plants gene regulates oxidative stress tolerance in Arabidopsis thaliana. Cell. Mol. Life Sci. 2020, 77, 705–718. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Huang, W.; Li, M.; Xu, Z.; Wang, F.; Xiong, A. Expression profiles of genes involved in jasmonic acid biosynthesis and signaling during growth and development of carrot. Acta Biochim. Et Biophys. Sin. 2016, 48, 795–803. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Duan, Y.; Hu, G.; Geng, X.; Zhang, G.; Yan, P.; Liu, Z.; Zhang, L.; Song, X. Identification of candidate genes and biosynthesis pathways related to fertility conversion by wheat KTM3315A transcriptome profiling. Front. Plant Sci. 2017, 8, 449. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Huang, Y.; Dong, X.; Wang, R.; Tang, M.; Cai, J.; Chen, J.; Zhang, X.; Nie, G. Exogenous methyl jasmonate improves heat tolerance of perennial ryegrass through alteration of osmotic adjustment, antioxidant defense, and expression of jasmonic acid-responsive genes. Front. Plant Sci. 2021, 12, 664519. [Google Scholar] [CrossRef]
- Mulaudzi, T.; Nkuna, M.; Sias, G.; Doumbia, I.Z.; Njomo, N.; Iwuoha, E. Antioxidant Capacity of Chitosan on Sorghum Plants under Salinity Stress. Agriculture 2022, 12, 1544. [Google Scholar] [CrossRef]
- Rakgotho, T.; Ndou, N.; Mulaudzi, T.; Iwuoha, E.; Mayedwa, N.; Ajayi, R.F. Green-Synthesized Zinc Oxide Nanoparticles Mitigate Salt Stress in Sorghum bicolor. Agriculture 2022, 12, 597. [Google Scholar] [CrossRef]
- Ahmadi, F.I.; Karimi, K.; Struik, P.C. Effect of exogenous application of methyl jasmonate on physiological and biochemical characteristics of Brassica napus L. cv. Talaye under salinity stress. S. Afr. J. Bot. 2018, 115, 5–11. [Google Scholar] [CrossRef]
- Limani, N.; Arraouadi, S.; Hajlaoui, H. Biochemical and Physiological Behavior against Salt Stress effect on two Quinoa accessions (Chenopodium quinoa Willd.). Int. J. Agric. Anim. Prod. 2022, 2, 9–19. [Google Scholar]
- Kanwal, S.; Ilyas, N.; Shabir, S.; Saeed, M.; Gul, R.; Zahoor, M.; Batool, N.; Mazhar, R. Application of biochar in mitigation of negative effects of salinity stress in wheat (Triticum aestivum L.). J. Plant Nutr. 2017, 41, 526–538. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; Gong, D.; Gao, Y.; Pan, R.; Hu, J.; Guan, Y. Priming with methyl jasmonate alleviates polyethylene glycol-induced osmotic stress in rice seeds by regulating the seed metabolic profile. Environ. Exp. Bot. 2018, 153, 236–248. [Google Scholar] [CrossRef]
- Mousavi, S.R.; Niknejad, Y.; Fallah, H.; Tari, D.B. Methyl jasmonate alleviates arsenic toxicity in rice. Plant Cell Rep. 2020, 39, 1041–1060. [Google Scholar] [CrossRef]
- Lalotra, S.; Hemantaranjan, A.; Srivastava, R. Alleviating role of methyl jasmonate and zinc on morpho-physiological and biochemical attributes in chickpea (Cicer arietinum L.) under salinity stress. J. Pharmacogn. Phytochem. 2019, 8, 527–532. [Google Scholar]
- Labiad, M.H.; Giménez, A.; Varol, H.; Tüzel, Y.; Egea-Gilabert, C.; Fernández, J.A.; Martínez-Ballesta, M.d.C. Effect of Exogenously Applied Methyl Jasmonate on Yield and Quality of Salt-Stressed Hydroponically Grown Sea Fennel (Crithmum maritimum L.). Agronomy 2021, 11, 1083. [Google Scholar] [CrossRef]
- Anjum, S.A.; Tanveer, M.; Hussain, S.; Tung, S.A.; Samad, R.A.; Wang, L.; Khan, I.; Rehman, N.U.; Shah, A.N.; Shahzad, B. Exogenously applied methyl jasmonate improves the drought tolerance in wheat imposed at early and late developmental stages. Acta Physiol. Plant. 2015, 38, 25. [Google Scholar] [CrossRef]
- Taïbi, K.; Taïbi, F.; Abderrahim, L.A.; Ennajah, A.; Belkhodja, M.; Mulet, J.M. Effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidant defence systems in Phaseolus vulgaris L. S. Afr. J. Bot. 2016, 105, 306–312. [Google Scholar] [CrossRef]
- Baniasadi, F.; Saffari, V.R.; Moud, A.A.M. Physiological and growth responses of Calendula officinalis L. plants to the interaction effects of polyamines and salt stress. Sci. Hortic. 2018, 234, 312–317. [Google Scholar] [CrossRef]
- Nounjan, N.; Mahakham, W.; Siangliw, J.L.; Toojinda, T.; Theerakulpisut, P. Chlorophyll Retention and High Photosynthetic Performance Contribute to Salinity Tolerance in Rice Carrying Drought Tolerance Quantitative Trait Loci (QTLs). Agriculture 2020, 10, 620. [Google Scholar] [CrossRef]
- Reddy, K.N.; Hoagland, R.E.; Zablotowicz, R.M. Effect of glyphosate on growth, chlorophyll, and nodulation in glyphosate-resistant and susceptible soybean (Glycine max) varieties. J. New Seeds 2000, 2, 37–52. [Google Scholar] [CrossRef]
- Nguyen, D.; Rieu, I.; Mariani, C.; van Dam, N.M. How plants handle multiple stresses: Hormonal interactions underlying responses to abiotic stress and insect herbivory. Plant Mol. Biol. 2016, 91, 727–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taheri, Z.; Vatankhah, E.; Jafarian, V. Methyl jasmonate improves physiological and biochemical responses of Anchusa italica under salinity stress. S. Afr. J. Bot. 2020, 130, 375–382. [Google Scholar] [CrossRef]
- Mahmoud, L.M.; Vincent, C.I.; Grosser, J.W.; Dutt, M. The response of salt-stressed Valencia sweet orange (Citrus sinensis) to salicylic acid and methyl jasmonate treatments. Plant Physiol. Rep. 2021, 26, 137–151. [Google Scholar] [CrossRef]
- Özdemir, B.; Tanyolac, Z.Ö.; Ulukapı, K.; Onus, A.N. Evaluation of salinity tolerance level of some pepper (Capsicum annuum L.) cultivars. Int. J. Agric. Innov. Res. 2016, 5, 247–251. [Google Scholar]
- Ranjbarfordoei, A.; Samson, R.; Van Damme, P. Chlorophyll fluorescence performance of sweet almond [Prunus dulcis (Miller) D. Webb] in response to salinity stress induced by NaCl. Photosynthetica 2006, 44, 513–522. [Google Scholar] [CrossRef]
- Lang, D.; Yu, X.; Jia, X.; Li, Z.; Zhang, X. Methyl jasmonate improves metabolism and growth of NaCl-stressed Glycyrrhiza uralensis seedlings. Sci. Hortic. 2020, 266, 109287. [Google Scholar] [CrossRef]
- Daudi, A.; O′Brien, J.A. Detection of Hydrogen Peroxide by DAB Staining in Arabidopsis Leaves. Bio-Protocol 2012, 2, e263. [Google Scholar] [CrossRef] [Green Version]
- Karimi, R.; Gavili-Kilaneh, K.; Khadivi, A. Methyl jasmonate promotes salinity adaptation responses in two grapevine (Vitis vinifera L.) cultivars differing in salt tolerance. Food Chem. 2022, 375, 131667. [Google Scholar] [CrossRef]
- Ahmad, F.; Kamal, A.; Singh, A.; Ashfaque, F.; Alamri, S.; Siddiqui, M.H. Salicylic Acid Modulates Antioxidant System, Defense Metabolites, and Expression of Salt Transporter Genes in Pisum sativum Under Salinity Stress. J. Plant Growth Regul. 2020, 41, 1905–1918. [Google Scholar] [CrossRef]
- Shah, T.; Latif, S.; Saeed, F.; Ali, I.; Ullah, S.; Alsahli, A.A.; Jan, S.; Ahmad, P. Seed priming with titanium dioxide nanoparticles enhances seed vigor, leaf water status, and antioxidant enzyme activities in maize (Zea mays L.) under salinity stress. J. King Saud Univ. Sci. 2021, 33, 101207. [Google Scholar] [CrossRef]
- El-Amier, Y.; Elhindi, K.; El-Hendawy, S.; Al-Rashed, S.; Abd-ElGawad, A. Antioxidant system and biomolecules alteration in Pisum sativum under heavy metal stress and possible alleviation by 5-aminolevulinic acid. Molecules 2019, 24, 4194. [Google Scholar] [CrossRef] [Green Version]
- Farsaraei, S.; Moghaddam, M.; Pirbalouti, A.G. Changes in growth and essential oil composition of sweet basil in response of salinity stress and superabsorbents application. Sci. Hortic. 2020, 271, 109465. [Google Scholar] [CrossRef]
- Hannan, A.; Hassan, L.; Hoque, N.; Arif, T.-U.; Robin, A.H.K. Increasing New Root Length Reflects Survival Mechanism of Rice (Oryza sativa L.) Genotypes under PEG-Induced Osmotic Stress. Plant Breed. Biotechnol. 2020, 8, 46–57. [Google Scholar] [CrossRef]
- Tarchoune, I.; Sgherri, C.; Izzo, R.; Lachaâl, M.; Navari-Izzo, F.; Ouerghi, Z. Changes in the antioxidative systems of Ocimum basilicum L. (cv. Fine) under different sodium salts. Acta Physiol. Plant. 2012, 34, 1873–1881. [Google Scholar] [CrossRef]
- Rui, M.; Ma, C.; Hao, Y.; Guo, J.; Rui, Y.; Tang, X.; Zhao, Q.; Fan, X.; Zhang, Z.; Hou, T.; et al. Iron Oxide Nanoparticles as a Potential Iron Fertilizer for Peanut (Arachis hypogaea). Front. Plant Sci. 2016, 7, 815. [Google Scholar] [CrossRef] [Green Version]
- Manzoor, H.; Mehwish; Bukhat, S.; Rasul, S.; Rehmani, M.I.A.; Noreen, S.; Athar, H.-U.; Zafar, Z.U.; Skalicky, M.; Soufan, W.; et al. Methyl Jasmonate Alleviated the Adverse Effects of Cadmium Stress in Pea (Pisum sativum L.): A Nexus of Photosystem II Activity and Dynamics of Redox Balance. Front. Plant Sci. 2022, 13, 860664. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Zhang, J.; Zhang, J.; Zhang, W.; Zheng, L.; Borjigin, T.; Wang, Y. Nitric oxide alleviates salt-induced stress damage by regulating the ascorbate–glutathione cycle and Na+/K+ homeostasis in Nitraria tangutorum Bobr. Plant Physiol. Biochem. 2022, 173, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Vighi, I.L.; Benitez, L.C.; Amaral, M.N.; Moraes, G.P.; Auler, P.A.; Rodrigues, G.S.; Deuner, S.; Mala, L.C.; Braga, E.J.B. Functional characterization of the antioxidant enzymes in rice plants exposed to salinity stress. Biol. Plant. 2017, 61, 540–550. [Google Scholar] [CrossRef]
- He, W.; Luo, H.; Xu, H.; Zhou, Z.; Li, D.; Bao, Y.; Fu, Q.; Song, J.; Jiao, Y.; Zhang, Z. Effect of exogenous methyl jasmonate on physiological and carotenoid composition of yellow maize sprouts under NaCl stress. Food Chem. 2021, 361, 130177. [Google Scholar] [CrossRef] [PubMed]
- Jafari, S.H.; Arani, A.M.; Esfahani, S.T. The Combined Effects of Rhizobacteria and Methyl Jasmonate on Rosmarinic Acid Production and Gene Expression Profile in Origanum Vulgare l. Under Salinity Conditions. J. Plant Growth Regul. 2022, 42, 1472–1487. [Google Scholar] [CrossRef]
- Nadeem, M.; Anwar-Ul-Haq, M.; Saqib, M.; Maqsood, M.; He, Z. Ameliorative Effect of Silicic Acid and Silicates on Oxidative, Osmotic Stress, and Specific Ion Toxicity in Spring Wheat (Triticum aestivum L.) Genotypes. J. Soil Sci. Plant Nutr. 2022, 22, 2334–2345. [Google Scholar] [CrossRef]
- Latef, A.A.; Arif, T.U.; Rhaman, M. Exogenous Auxin-Mediated Salt Stress Alleviation in Faba Bean (Vicia faba L.). Agronomy 2021, 11, 547. [Google Scholar] [CrossRef]
- Li, H.; Testerink, C.; Zhang, Y. How roots and shoots communicate through stressful times. Trends Plant Sci. 2021, 26, 940–952. [Google Scholar] [CrossRef]
- Zörb, C.; Mühling, K.H.; Kutschera, U.; Geilfus, C.-M. Salinity stiffens the epidermal cell walls of salt-stressed maize leaves: Is the epidermis growth-restricting? PLoS ONE 2015, 10, e0118406. [Google Scholar] [CrossRef] [Green Version]
- Pacheco-Silva, N.V.; Donato, A.M. Morpho-anatomy of the leaf of Myrciaria glomerata. Rev. Bras. de Farm. 2016, 26, 275–280. [Google Scholar] [CrossRef] [Green Version]
- Brookbank, B.P.; Patel, J.; Gazzarrini, S.; Nambara, E. Role of Basal ABA in Plant Growth and Development. Genes 2021, 12, 1936. [Google Scholar] [CrossRef]
- Hwang, B.G.; Ryu, J.; Lee, S.J. Vulnerability of Protoxylem and Metaxylem Vessels to Embolisms and Radial Refilling in a Vascular Bundle of Maize Leaves. Front. Plant Sci. 2016, 7, 941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kollist, H.; Zandalinas, S.I.; Sengupta, S.; Nuhkat, M.; Kangasjärvi, J.; Mittler, R. Rapid Responses to Abiotic Stress: Priming the Landscape for the Signal Transduction Network. Trends Plant Sci. 2018, 24, 25–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaz, M.A.; Zakharenko, A.M.; Zemchenko, I.V.; Haider, M.S.; Ali, M.A.; Imtiaz, M.; Chung, G.; Tsatsakis, A.; Sun, S.; Golokhvast, K.S. Phytolith Formation in Plants: From Soil to Cell. Plants 2019, 8, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meunier, J.D.; Barboni, D.; Anwar-Ul-Haq, M.; Levard, C.; Chaurand, P.; Vidal, V.; Grauby, O.; Huc, R.; Laffont-Schwob, I.; Rabier, J.; et al. Effect of phytoliths for mitigating water stress in durum wheat. New Phytol. 2017, 215, 229–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westworth, S.; Ashwath, N.; Cozzolino, D. Application of FTIR-ATR spectroscopy to detect salinity response in Beauty Leaf Tree (Calophyllum inophyllum L). Energy Procedia 2019, 160, 761–768. [Google Scholar] [CrossRef]
- Končitíková, R.; Vigouroux, A.; Kopečná, M.; Andree, T.; Bartoš, J.; Šebela, M.; Moréra, S.; Kopečný, D. Role and structural characterization of plant aldehyde dehydrogenases from family 2 and family 7. Biochem. J. 2015, 468, 109–123. [Google Scholar] [CrossRef]
- Kishor, P.B.K.; Kumari, P.H.; Sunita, M.S.L.; Sreenivasulu, N. Role of proline in cell wall synthesis and plant development and its implications in plant ontogeny. Front. Plant Sci. 2015, 6, 544. [Google Scholar] [CrossRef] [Green Version]
- Raza, A.; Charagh, S.; Zahid, Z.; Mubarik, M.S.; Javed, R.; Siddiqui, M.H.; Hasanuzzaman, M. Jasmonic acid: A key frontier in conferring abiotic stress tolerance in plants. Plant Cell Rep. 2021, 40, 1513–1541. [Google Scholar] [CrossRef] [PubMed]
- Rezai, S.; Orojloo, M.; Bidabadi, S.S.; Soleimanzadeh, M. Possible Role of Methyl Jasmonate in Protection to NaCl-Induced Salt Stress in Pepper cv. “Green Hashemi”. Int. J. Agric. Crop Sci. 2013, 6, 1235. [Google Scholar]
- Tavallali, V.; Karimi, S. Methyl jasmonate enhances salt tolerance of almond rootstocks by regulating endogenous phytohormones, antioxidant activity and gas-exchange. J. Plant Physiol. 2019, 234, 98–105. [Google Scholar] [CrossRef]
- Chini, A.; Fonseca, S.; Fernández, G.; Adie, B.; Chico, J.M.; Lorenzo, O.; García-Casado, G.; López-Vidriero, I.; Lozano, F.M.; Ponce, M.R.; et al. The JAZ family of repressors is the missing link in jasmonate signalling. Nature 2007, 448, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Thines, B.; Katsir, L.; Melotto, M.; Niu, Y.; Mandaokar, A.; Liu, G.; Nomura, K.; He, S.Y.; Howe, G.A.; Browse, J. JAZ repressor proteins are targets of the SCFCOI1 complex during jasmonate signalling. Nature 2007, 448, 661–665. [Google Scholar] [CrossRef]
- De Domenico, S.; Aurino, M.; Gallo, A.; Poltronieri, P.; Pastor, V.; Flors, V.; Santino, A. Oxylipin dynamics in Medicago truncatula in response to salt and wounding stresses. Physiol. Plant. 2019, 165, 198–208. [Google Scholar] [CrossRef]
- Kashyap, S.P.; Prasanna, H.C.; Kumari, N.; Mishra, P.; Singh, B. Understanding salt tolerance mechanism using transcriptome profiling and de novo assembly of wild tomato Solanum chilense. Sci. Rep. 2020, 10, 15835. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Q.; Zhai, H.; Li, Y.; Wang, X.; Liu, Q.; He, S. Transcript profile analysis reveals important roles of jasmonic acid signalling pathway in the response of sweet potato to salt stress. Sci. Rep. 2017, 7, 40819. [Google Scholar] [CrossRef] [Green Version]
- Qiu, X.; Xu, Y.; Xiong, B.; Dai, L.; Huang, S.; Dong, T.; Sun, G.; Liao, L.; Deng, Q.; Wang, X.; et al. Effects of exogenous methyl jasmonate on the synthesis of endogenous jasmonates and the regulation of photosynthesis in citrus. Physiol. Plant. 2020, 170, 398–414. [Google Scholar] [CrossRef]
- Lado, J.; Manzi, M. Metabolic and Hormonal Responses of Plants to Cold Stress. In Medicinal Plants and Environmental Challenges; Springer: Berlin/Heidelberg, Germany, 2017; pp. 137–158. [Google Scholar]
- Singh, P.; Arif, Y.; Siddiqui, H.; Hayat, S. Jasmonate: A versatile messenger in plants. In Jasmonates and Salicylates Signaling in Plants; Springer: Berlin/Heidelberg, Germany, 2021; pp. 129–158. [Google Scholar]
- Mulaudzi-Masuku, T.; Mutepe, R.D.; Mkhoro, O.C.; Faro, A.; Ndimba, B. Identification and characterization of a heat-inducible Hsp70 gene from Sorghum bicolor which confers tolerance to thermal stress. Cell Stress Chaperones 2015, 20, 793–804. [Google Scholar] [CrossRef] [Green Version]
- Rahman, A.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Calcium Supplementation Improves Na+/K+ Ratio, Antioxidant Defense and Glyoxalase Systems in Salt-Stressed Rice Seedlings. Front. Plant Sci. 2016, 7, 609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilcacundo, R.; Barrio, D.A.; Piñuel, L.; Boeri, P.; Tombari, A.; Pinto, A.; Welbaum, J.; Hernández-Ledesma, B.; Carrillo, W. Inhibition of Lipid Peroxidation of Kiwicha (Amaranthus caudatus) Hydrolyzed Protein Using Zebrafish Larvae and Embryos. Plants 2018, 7, 69. [Google Scholar] [CrossRef] [Green Version]
- Heath, R.L.; Packer, L. Reprint of: Photoperoxidation in Isolated Chloroplasts I. Kinetics and Stoichiometry of Fatty Acid Peroxidation. Arch. Biochem. Biophys. 2022, 726, 109248. [Google Scholar] [CrossRef] [PubMed]
- Mackinney, G. Absorption of Light by Chlorophyll Solutions. J. Biol. Chem. 1941, 140, 315–322. [Google Scholar] [CrossRef]
- Wellburn, A.R.; Lichtenthaler, H. Formulae and Program to Determine Total Carotenoids and Chlorophylls A and B of Leaf Extracts in Different Solvents. In Advances in Photosynthesis Research: Proceedings of the VIth International Congress on Photosynthesis, Brussels, Belgium, 1–6 August 1983; Springer: Berlin/Heidelberg, Germany, 1984; Volume 2, pp. 9–12. [Google Scholar]
- Carillo, P.; Gibon, Y. Protocol: Extraction and determination of proline. Prometh. Wiki 2011, 2011, 1–5. [Google Scholar]
- Watanabe, S.; Kojima, K.; Ide, Y.; Sasaki, S. Effects of saline and osmotic stress on proline and sugar accumulation in Populus euphratica in vitro. Plant Cell Tissue Organ Cult. (PCTOC) 2000, 63, 199–206. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NaCl (mM) | MeJa (µM) | Fresh Weight (g) | Dry Weight (g) |
|---|---|---|---|
| 0 | 0 | 0.800 ± 0.189 a | 0.091 ± 0.013 a |
| 200 | 0 | 0.397 ± 0.027 b | 0.053 ± 0.005 b |
| 10 | 0.730 ± 0.031 a | 0.110 ± 0.005 a | |
| 15 | 0.665 ± 0.039 ab | 0.102 ± 0.00 ab |
| NaCl (mM) | MeJa (µM) | Chlorophyll a (mg/gFW) | Chlorophyll b (mg/gFW) | Total Chlorophyll (mg/gFW) |
|---|---|---|---|---|
| 0 | 0 | 8.17 ± 1.890 ab | 4.392 ± 1.075 a | 11.96 ± 2.98 ab |
| 200 | 0 | 4.308 ± 0.949 b | 3.172 ± 0.820 b | 6.667 ± 2.078 b |
| 10 | 11.992 ± 3.440 a | 5.011 ± 0.516 a | 16.710 ± 3.900 a | |
| 15 | 9.104 ± 0.656 ab | 4.585 ± 0.513 ab | 13.438 ± 0.151 a |
| Elements | 0 mM NaCl (wt%) | 200 mM NaCl (wt%) | 200 mM + 10 µM MeJa (wt%) | 200 mM + 15 µM MeJa (wt%) |
|---|---|---|---|---|
| Na+ | 0.00 ± 0.00 c | 2.16 ± 0.52 a | 0.97 ± 0.24 b | 0.635 ± 0.155 bc |
| Si+ | 0.295 ± 0.095 a | 0.00 ± 0.00 b | 0.72 0.24 ab | 0.675 ± 0.225 ab |
| K+ | 7.46 ± 3.38 a | 1.49 ± 0.17 b | 2.53 ± 1.2 ab | 4.17 ± 1.97 ab |
| Element Ratio | ||||
| Na+/K+ | 0.00 ± 0.00 c | 1.436 ± 0.186 a | 0.419 ± 0.117 b | 0.166 ± 0.047 bc |
| Gene Name | Accession Numbers | Amplicon Size | Primer Sequence | ||
|---|---|---|---|---|---|
| LOC ID | mRNA Sequence ID | Forward (5′-3′) | Reverse (5′-3′) | ||
| SbLOX | LOC8064218 | GQ369443.1 | 204 bp | GTACCGCTACGACGTCTACA | GTCAACTCTCGTGCAGCAAA |
| SbAOS | LOC8086184 | XM_002463784.2 | 191 bp | ACCATCACCTCGCTCAAGAA | TCACACAGTATCACGGCACT |
| SbAOC | LOC8063218 | XM_002465042.2 | 161 bp | GTACGAGGCCATCTACAGCT | AGGGGAAGACGATCTGGTTG |
| SbOPR | LOC8070775 | XM_002438007.2 | 166 bp | GGGTATGATCGGGAGGAAGG | CAACGGGATCTTGCGTGTAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mulaudzi, T.; Sias, G.; Nkuna, M.; Ndou, N.; Hendricks, K.; Ikebudu, V.; Koo, A.J.; Ajayi, R.F.; Iwuoha, E. Seed Priming with MeJa Prevents Salt-Induced Growth Inhibition and Oxidative Damage in Sorghum bicolor by Inducing the Expression of Jasmonic Acid Biosynthesis Genes. Int. J. Mol. Sci. 2023, 24, 10368. https://doi.org/10.3390/ijms241210368
Mulaudzi T, Sias G, Nkuna M, Ndou N, Hendricks K, Ikebudu V, Koo AJ, Ajayi RF, Iwuoha E. Seed Priming with MeJa Prevents Salt-Induced Growth Inhibition and Oxidative Damage in Sorghum bicolor by Inducing the Expression of Jasmonic Acid Biosynthesis Genes. International Journal of Molecular Sciences. 2023; 24(12):10368. https://doi.org/10.3390/ijms241210368
Chicago/Turabian StyleMulaudzi, Takalani, Gershwin Sias, Mulisa Nkuna, Nzumbululo Ndou, Kaylin Hendricks, Vivian Ikebudu, Abraham J. Koo, Rachel F. Ajayi, and Emmanuel Iwuoha. 2023. "Seed Priming with MeJa Prevents Salt-Induced Growth Inhibition and Oxidative Damage in Sorghum bicolor by Inducing the Expression of Jasmonic Acid Biosynthesis Genes" International Journal of Molecular Sciences 24, no. 12: 10368. https://doi.org/10.3390/ijms241210368