
Systematic Review: Urine Biomarker Discovery for Inflammatory Bowel Disease Diagnosis

Abstract
:1. Introduction
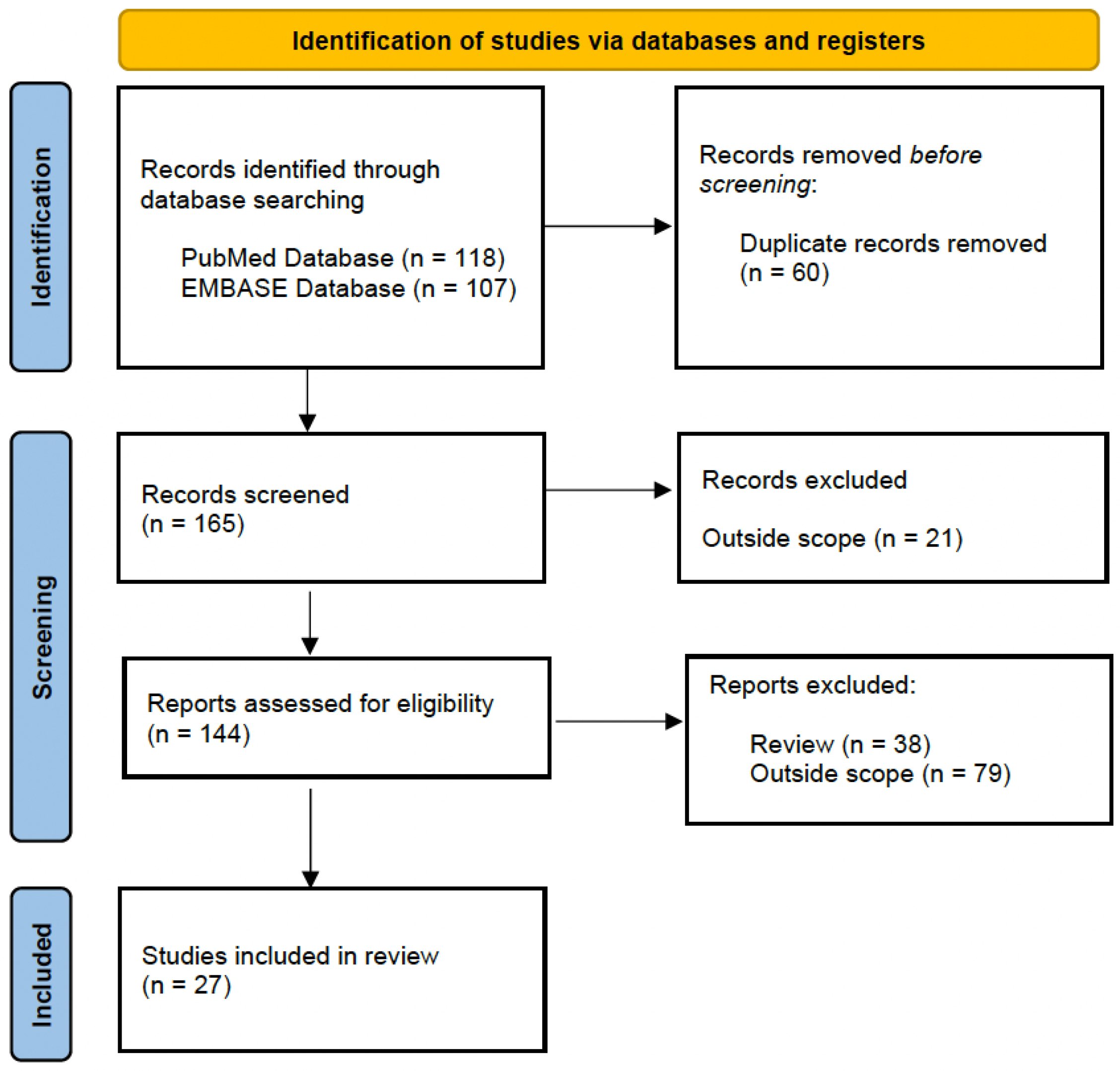
2. Methods
2.1. Literature Search Strategy
2.2. Proteomics and Metabolomics Approaches to Biomarker Discovery
2.2.1. Metabolomics
2.2.2. Proteomics
3. Results and Discussion
3.1. Urinary Biomarkers for IBD Diagnosis
3.2. Animal Models of IBD
3.2.1. Chemically Induced Mouse Models
DSS-Induced Colitis Model
TNBS-Induced Colitis Model
3.2.2. Genetically Engineered Mouse Models of IBD
IL-10-Gene-Deficient Mouse Model

{kind=link}
| Reference | Animal Model | Urinary Biomarker | Detection Methods | Sample Size (Discovery Phase) | Biological Findings |
|---|---|---|---|---|---|
| Dong et al. [47] | DSS-induced acute UC | Metabolites | NMR spectroscopy | 12 DSS-induced acute UC and 12 controls | Higher levels of citric acid cycle intermediates, butyrate, and phenylacetate and lower concentrations of 2-(4-hydroxyphenyl) propanoic acid, methylamine, phenylacetylglycine, indoxyl sulfate, hippurate, 4-cresol glucuronide, 4-cresol sulfate, adipate, azelate, N methylnicotinate, and methyguanidine in urine samples from DSS-treated mice. |
| Schicho et al. [48] | DSS-induced acute UC | Metabolites | NMR spectroscopy | 11 DSS-induced acute UC and 5 controls | Increased levels of ketone bodies, hypoxanthine, and tryptophan and decreased levels of antioxidant metabolites. |
| Qin et al. [51] | TNBS-induced colitis rat model | Proteins | TMT-label quantitative LC-MS/MS | 18 TNBS-induced colitis and 10 controls | A total of 77 proteins were significantly expressed in colitis rats. Nine proteins were validated via PRM. Of these, carbonic anhydrase 1, neutrophil collagenase, and neutrophil gelatinase-associated lipocalin were previously reported as IBD-associated proteins, whereas collectrin, beta-mannosidase, sodium-dependent neutral amino acid transporter B(0)AT3, glyceraldehyde-3-phosphate dehydrogenase and ribonuclease pancreatic gamma-type were newly discovered. |
| Zhang et al. [52] | TNBS-induced acute colitis | Metabolites | UPLC-ESI-QTOF-MS | 7 TNBS-induced acute colitis and 10 control rats | Two tryptophan metabolites (4-(2-aminophenyl)-2,4-dioxobutanoic acid and 4,6-cihydroxyquinoline), two gut microbial metabolites (phenyl-acetylglycine and p-cresol glucuronide), and the bile acid 12a-hydroxy-3-oxocholadienic acid were identified in urine from the TNBS-induced acute colitis rats. |
| Tso et al. [54] | IL-10-gene-deficient mice | Metabolites | NMR spectroscopy | 10 IL-10-gene-deficient mice and 10 wild-type mice | Metabolomics profile from IL-10-gene-deficient mice is gender-, age-, and disease-specific. Changes in profile patterns that appear to be imperative for the development of intestinal inflammation were observed during the development of IBD. |
| Lin et al. [55] | IL-10-gene-deficient mice | Metabolites | GC-MS | 15 IL-10-gene-deficient mice and 10 wild-type mice | Tryptophan metabolism, fucosylation and fatty acid metabolism were perturbed in IL10−/−mice. |
| Lin et al. [56] | IL-10-gene-deficient mice | Metabolites | GC-MS | 20 IL-10-gene-deficient mice and 20 wild-type mice | Identification of fifteen metabolite differences associated with intestinal inflammation and metabolite differences unrelated to inflammation that may indicate novel functions of IL10. |
| Otter et al. [57] | IL-10-gene-deficient mice | Metabolites | Short-column LC-MS | IL-10-gene-deficient mice and wild-type mice | Three differential metabolites were associated with colon inflammation. |
| Murdoch et al. [58] | IL-10-gene-deficient mice | Metabolites | NMR spectroscopy | 4 IL-10-gene-deficient mice and 4 wild-type mice | The metabolic profiles of control and IL-10-gene-deficient mice diverged substantially with the onset of IBD. |
3.3. Urine Samples in IBD
| Reference | Urinary Biomarker | Detection Methods | Sample Size (Discovery Phase) | Treatments | Biological Findings |
|---|---|---|---|---|---|
| Maráková et al. [60] | Proteins | CE-MS/MS | 13 IBD and 6 HC | Azathioprine | Downregulation of serotonin and norepinephrine, and overexpression of histamine and spermidine. |
| Stephens et al. [61] | Metabolites | NMR spectroscopy | 30 CD, 30 UC, and 60 HC | Mesalamine, corticoesteroid, thiopurine, antimetabolite, and anti-TNF | Major differences between IBD and HC including TCA cycle intermediates, amino acids, and gut microflora metabolites. |
| Schicho et al. [62] | Metabolites | NMR spectroscopy | 20 UC, 20 CD, and 40 HC | 5-ASA, steroids, 6-MP, azathioprinea, and immunosupresives | Higher levels of mannitol, allantoin, xylose, and carnitine and decreased levels of betaine and hippurate. |
| Cracowski et al. [63] | Metabolites | GC/electronic impact MS | 23 CD and 23 HC | 5-ASA, corticoesteroid, azathioprine, methotrexate, and cyclosporine | Increased iPF2alpha-III concentrations in patients with CD. A correlation was found between urinary iPF2alpha-III and plasma C-reactive protein concentrations. |
| Williams et al. [64] | Metabolites | NMR spectroscopy | 86 CD, 60 UC, and 60 HC | 5-ASA, azathioprine, and prednisolone | Three specific urinary metabolites related to gut microbial metabolism (hippurate, formate, and 4-cresol sulfate differ between CD, UC, and HC. |
| Martin et al. [65] | Metabolites | NMR spectroscopy | 21 pediatric IBD and 27 HC | No data | The identification of two readouts of nitrogen metabolic (urea and phenylacetylglutamine) may be relevant to monitoring metabolic status in relation to disease state. |
| Martin et al. [66] | Metabolites | LC-MS/MS and GC-MS | 21 pediatric IBD and 27 HC | No data | Metabolic differences encompass central energy metabolism, amino acids, bile acids, and gut microbial metabolites. Levels of pyroglutamic acid, glutamic acid, glycine, and cysteine were significantly higher in children with IBD. |
| El Hassani et al. [67] | Metabolites | GC-MS | 10 pediatric IBD and 10 HC | None | Significant difference in VOC profiles between IBD and HC. |
| Yamamoto et al. [68] | Metabolites | MSI-CE-MS | 26 pediatric IBD | Anti-TNF, immunomodulator, and 5-ASA | Increased excretions of indoxyl sulfate, hydroxyindoxyl sulfate, phenylacetylglutamine, and sialic acid and lower threonine, serine, kynurenine, and hypoxanthine levels in CD compared to UC patients. Excellent discrimination between groups was achieved based on the urinary serine:indoxylsulfate ratio (AUC = 0.972). |
| Siebert et al. [69] | Proteins | CE-MS | 50 IBD (42 CD and 8 CU), 50 OA, 50 RA, 50 PsA and 50 HC | No data | The classifiers for the five groups demonstrated excellent performances, with AUC values between 0.90 and 0.97 per group. |
| Alonso A et al. [70] | Metabolites | NMR spectroscopy | 1210 IMIDs and 100 HC | Anti-TNF, methotrexate, and 5-ASA | The use of the combination of all diagnostic biomarkers as multivariate classifiers demonstrated a good disease prediction accuracy in all IMIDs and particularly in IBD. |
| Keshteli et al. [71] | Metabolites | LC-MS/MS and GC-MS | 53 qUC, 39 IBS and 21 HC | 5-ASA, immunosuppresants, and biologics. | A unique urinary metabolome in IBS patients that could differentiate them from UC patients with an AUC = 0.99. |
| Dawiskiba et al. [72] | Metabolites | NMR spectroscopy | 24 UC, 19 CD and 17 HC | 5-ASA, azathioprine, and acetaminophen | Differences in citrate, hippurate, trigonelline, taurine, succinate, and 2-hydroxybutyrate levels were found in patients with active IBD. Patients with IBD in remission showed up-regulated levels of acetoacetate and decreased levels of citrate, hippurate, taurine, succinate, glycine, alanine, and formate. |
| Markó et al. [73] | Proteins | SE HPLC and MALDI-TOF/MS | 1 CD | Mesalamine and azathioprine | The concentration of t-uAlb was found to be 15 times higher than that of their ir-uAlb during the active state. |
| Bjerrum et al. [74] | Metabolites | NMR spectroscopy | 41 aUC, 33 qUC, and- 25 HC | Mesalazine, glucocorticoids, azathioprine, and anti-TNF | The metabolic profiles of the urine did not allow for differentiation between active UC, inactive UC, and HC. |
| Stanke-Labesque et al. [75] | Metabolites | LC-MS/MS | 32 CD, 28 UC, and 30 HC | Azathioprine, 5-ASA, and glucocorticoids | LTE4 urinary excretion was significantly higher in CD and UC patients compared to HC. LTE4 levels were higher in patients with active disease than in patients in remission. |
| Keshteli et al. [76] | Metabolites | NMR spectroscopy and LC-MS/MS | 38 CD | 5-ASA, azathioprine/6-MP, methotrexate, corticosteroids, and anti-TNF | Endoscopic recurrence was associated with an increased concentration of urinary levoglucosan. Rutgeerts score was positively correlated with levoglucosan and propylene glycol levels. |
| Keshteli et al. [77] | Metabolites | NMR spectroscopy and LC-MS/MS | 20 UC | 5-ASA and immunosuppresants | Three metabolites in urine (trans-aconitate, cysteine and acetamide) and three metabolites in serum (3-hydroxybutyrate, acetoacetate and acetone) were responsible for the discrimination of UC patients with clinical relapse. |
3.3.1. Urinary Markers in the Differentiation of IBD Patients from Healthy Controls (HCs)
3.3.2. Comparison of Proteomics and Metabolomics Profiles between Patients with IBD and Patients with Other Inflammatory Diseases
3.3.3. Urinary Markers in Disease Activity Evaluation in IBD Patients
3.3.4. Urinary Metabolites as Biomarkers for Clinical Relapse in Patients with CD and UC
3.4. Correlation between Animal Models of IBD and Human IBD Diagnostic Markers
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2017, 390, 2769–2778. [Google Scholar] [CrossRef]
- Chaparro, M.; Garre, A.; Núñez Ortiz, A.; Diz-Lois Palomares, M.T.; Rodríguez, C.; Riestra, S.; Vela, M.; Benítez, J.M.; Fernández Salgado, E.; Sánchez Rodríguez, E.; et al. Incidence, clinical characteristics and management of inflammatory bowel disease in spain: Large-scale epidemiological study. J. Clin. Med. 2021, 10, 2885. [Google Scholar] [CrossRef]
- Schaub, S.; Wilkins, J.; Weiler, T.; Sangster, K.; Rush, D.; Nickerson, P. Urine protein profiling with surface-enhanced laser-desorption/ionization time-of-flight mass spectrometry. Kidney Int. 2004, 65, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Theodorescu, D.; Schiffer, E.; Bauer, H.W.; Douwes, F.; Eichhorn, F.; Polley, R.; Schmidt, T.; Schöfer, W.; Zürbig, P.; Good, D.M.; et al. Discovery and validation of urinary biomarkers for prostate cancer. Proteom. Clin. Appl. 2008, 2, 556. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y. Urine-an untapped goldmine for biomarker discovery? Sci. China Life Sci. 2013, 56, 1145–1146. [Google Scholar] [CrossRef] [Green Version]
- Lalmahomed, Z.S.; Bröker, M.E.E.; Van Huizen, N.A.; Coebergh van den Braak, R.R.; Dekker, L.J.; Rizopoulos, D.; Verhoef, C.; Steyerberg, E.W.; Luider, T.M.; Ijzermans, J.N. Hydroxylated collagen peptide in urine as biomarker for detecting colorectal liver metastases. Am. J. Cancer Res. 2016, 6, 321. [Google Scholar]
- Mallafré-muro, C.; Llambrich, M.; Cumeras, R.; Pardo, A.; Brezmes, J.; Marco, S.; Gumà, J. Comprehensive Volatilome and Metabolome Signatures of Colorectal Cancer in Urine: A Systematic Review and Meta-Analysis. Cancers 2021, 13, 2534. [Google Scholar] [CrossRef]
- Ohta, R.; Yamada, T.; Sonoda, H.; Matsuda, A.; Shinji, S.; Takahashi, G.; Iwai, T.; Takeda, K.; Ueda, K.; Kuriyama, S.; et al. Detection of KRAS mutations in circulating tumour DNA from plasma and urine of patients with colorectal cancer. Eur. J. Surg. Oncol. 2021, 47, 3151–3156. [Google Scholar] [CrossRef]
- Nam, D.; Lee, J.Y.; Lee, M.; Kim, J.; Seol, W.; Son, I.; Ho, D.H. Detection and Assessment of α-Synuclein Oligomers in the Urine of Parkinson’s Disease Patients. J. Parkinsons Dis. 2020, 10, 981–991. [Google Scholar] [CrossRef] [PubMed]
- Yao, F.; Hong, X.; Li, S.; Zhang, Y.; Zhao, Q.; Du, W.; Wang, Y.; Ni, J. Urine-Based Biomarkers for Alzheimer’s Disease Identified Through Coupling Computational and Experimental Methods. J. Alzheimers Dis. 2018, 65, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Nolen, B.M.; Lokshin, A.E. The advancement of biomarker-based diagnostic tools for ovarian, breast, and pancreatic cancer through the use of urine as an analytical biofluid. Int. J. Biol. Markers 2011, 26, 141–152. [Google Scholar] [CrossRef]
- Henrotin, Y.; Gharbi, M.; Mazzucchelli, G.; Dubuc, J.E.; De Pauw, E.; Deberg, M. Fibulin 3 peptides Fib3-1 and Fib3-2 are potential biomarkers of osteoarthritis. Arthritis Rheum. 2012, 64, 2260–2267. [Google Scholar] [CrossRef]
- Maahs, D.M.; Siwy, J.; Argilés, À.; Cerna, M.; Delles, C.; Dominiczak, A.F.; Gayrard, N.; Iphöfer, A.; Jänsch, L.; Jerums, G.; et al. Urinary collagen fragments are significantly altered in diabetes: A link to pathophysiology. PLoS ONE 2010, 5, e13051. [Google Scholar] [CrossRef] [Green Version]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Gisbert, J.P.; Chaparro, M. Clinical Usefulness of Proteomics in Inflammatory Bowel Disease: A Comprehensive Review. J Crohns Colitis 2019, 13, 374–384. [Google Scholar] [CrossRef]
- Baldan-Martin, M.; Chaparro, M.; Gisbert, J.P. Tissue Proteomic Approaches to Understand the Pathogenesis of Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2021, 27, 1184–1200. [Google Scholar] [CrossRef]
- Aldars-García, L.; Chaparro, M.; Gisbert, J.P. Systematic Review: The Gut Microbiome and Its Potential Clinical Application in Inflammatory Bowel Disease. Microorganisms 2021, 9, 977. [Google Scholar] [CrossRef] [PubMed]
- Aldars-garcía, L.; Marin, A.C.; Chaparro, M.; Gisbert, J.P. The Interplay between Immune System and Microbiota in Inflammatory Bowel Disease: A Narrative Review. Int. J. Mol. Sci. 2021, 22, 3076. [Google Scholar] [CrossRef] [PubMed]
- Ortega Moreno, L.; Sanz-Garcia, A.; Fernández de la Fuente, M.J.; Arroyo Solera, R.; Fernández-Tomé, S.; Marin, A.C.; Mora-Gutierrez, I.; Fernández, P.; Baldan-Martin, M.; Chaparro, M.; et al. Serum adipokines as non-invasive biomarkers in Crohn’s disease. Sci. Rep. 2020, 10, 18027. [Google Scholar] [CrossRef] [PubMed]
- Pinu, F.R.; Beale, D.J.; Paten, A.M.; Kouremenos, K.; Swarup, S.; Schirra, H.J.; Wishart, D. Systems Biology and Multi-Omics Integration: Viewpoints from the Metabolomics Research Community. Metabolites 2019, 9, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karczewski, K.J.; Snyder, M.P. Integrative omics for health and disease. Nat. Rev. Genet. 2018, 19, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Hasin, Y.; Seldin, M.; Lusis, A. Multi-omics approaches to disease. Genome Biol. 2017, 18, 83. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, I.; Verma, S.; Kumar, S.; Jere, A.; Anamika, K. Multi-omics Data Integration, Interpretation, and Its Application. Bioinform. Biol. Insights 2020, 14, 1177932219899051. [Google Scholar] [CrossRef] [Green Version]
- Mirza, A.H.; Berthelsen, C.H.B.; Seemann, S.E.; Pan, X.; Frederiksen, K.S.; Vilien, M.; Gorodkin, J.; Pociot, F. Transcriptomic landscape of lncRNAs in inflammatory bowel disease. Genome Med. 2015, 7, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schüssler-Fiorenza Rose, S.M.; Contrepois, K.; Moneghetti, K.J.; Zhou, W.; Mishra, T.; Mataraso, S.; Dagan-Rosenfeld, O.; Ganz, A.B.; Dunn, J.; Hornburg, D.; et al. A longitudinal big data approach for precision health. Nat. Med. 2019, 25, 792–804. [Google Scholar] [CrossRef]
- Schulte, C.; Barwari, T.; Joshi, A.; Theofilatos, K.; Zampetaki, A.; Barallobre-Barreiro, J.; Singh, B.; Sörensen, N.A.; Neumann, J.T.; Zeller, T.; et al. Comparative Analysis of Circulating Noncoding RNAs Versus Protein Biomarkers in the Detection of Myocardial Injury. Circ. Res. 2019, 125, 328–340. [Google Scholar] [CrossRef]
- Langley, S.R.; Willeit, K.; Didangelos, A.; Matic, L.P.; Skroblin, P.; Barallobre-Barreiro, J.; Lengquist, M.; Rungger, G.; Kapustin, A.; Kedenko, L.; et al. Extracellular matrix proteomics identifies molecular signature of symptomatic carotid plaques. J. Clin. Investig. 2017, 127, 1546–1560. [Google Scholar] [CrossRef] [Green Version]
- Griss, J.; Viteri, G.; Sidiropoulos, K.; Nguyen, V.; Fabregat, A.; Hermjakob, H. ReactomeGSA—Efficient Multi-Omics Comparative Pathway Analysis. Mol. Cell Proteom. 2020, 19, 2115–2124. [Google Scholar] [CrossRef]
- Nagana Gowda, G.; Raftery, D. Biomarker Discovery and Translation in Metabolomics. Curr. Metab. 2013, 1, 227. [Google Scholar] [CrossRef]
- Weljie, A.M.; Newton, J.; Mercier, P.; Carlson, E.; Slupsky, C.M. Targeted profiling: Quantitative analysis of 1H NMR metabolomics data. Anal. Chem. 2006, 78, 4430–4442. [Google Scholar] [CrossRef]
- Balluff, B.; Frese, C.K.; Maier, S.K.; Schöne, C.; Kuster, B.; Schmitt, M.; Aubele, M.; Höfler, H.; Deelder, A.M.; Heck, A., Jr.; et al. De novo discovery of phenotypic intratumour heterogeneity using imaging mass spectrometry. J. Pathol. 2015, 235, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Roy, R.; Jedinak, A.; Moses, M.A. Mining the Human Proteome: Biomarker Discovery for Human Cancer and Metastases. Cancer J. 2015, 21, 327–336. [Google Scholar] [CrossRef]
- Ferreri, A.J.M.; Illerhaus, G.; Zucca, E.; Cavalli, F. Flows and flaws in primary central nervous system lymphoma. Nat. Rev. Clin. Oncol. 2010, 7, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Gregorich, Z.R.; Ge, Y. Top-down proteomics in health and disease: Challenges and opportunities. Proteomics 2014, 14, 1195–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altelaar, A.F.M.; Munoz, J.; Heck, A.J.R. Next-generation proteomics: Towards an integrative view of proteome dynamics. Nat. Rev. Genet. 2013, 14, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Rifai, N.; Gillette, M.A.; Carr, S.A. Protein biomarker discovery and validation: The long and uncertain path to clinical utility. Nat. Biotechnol. 2006, 24, 971–983. [Google Scholar] [CrossRef]
- Mischak, H.; Kolch, W.; Aivaliotis, M.; Bouyssié, D.; Court, M.; Dihazi, H.; Dihazi, G.H.; Franke, J.; Garin, J.; Gonzalez de Peredo, A.; et al. Comprehensive human urine standards for comparability and standardization in clinical proteome analysis. Proteom. Clin. Appl. 2010, 4, 464–478. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.Q.; Dekker, L.J.M.; Stingl, C.; Güzel, C.; De Marchi, T.; Martens, J.W.; Foekens, J.A.; Luider, T.M.; Umar, A. Quantitative proteomic analysis of microdissected breast cancer tissues: Comparison of label-free and SILAC-based quantification with shotgun, directed, and targeted MS approaches. J. Proteome Res. 2013, 12, 4627–4641. [Google Scholar] [CrossRef]
- Van Der Greef, J.; Stroobant, P.; Van Der Heijden, R. The role of analytical sciences in medical systems biology. Curr. Opin. Chem. Biol. 2004, 8, 559–565. [Google Scholar] [CrossRef]
- Gao, Y. Roadmap to the urine biomarker era. MOJ Proteom. Bioinform. 2014, 1, 00005. [Google Scholar] [CrossRef]
- Günther, C.; Martini, E.; Wittkopf, N.; Amann, K.; Weigmann, B.; Neumann, H.; Waldner, M.J.; Hedrick, S.M.; Tenzer, S.; Neurath, M.F.; et al. Caspase-8 regulates TNF-α-induced epithelial necroptosis and terminal ileitis. Nature 2011, 477, 335–339. [Google Scholar] [CrossRef] [Green Version]
- Powrie, F. Gut reactions: Immune pathways in the intestine in health and disease. EMBO Mol. Med. 2012, 4, 71. [Google Scholar] [CrossRef]
- Wirtz, S.; Neufert, C.; Weigmann, B.; Neurath, M.F. Chemically induced mouse models of intestinal inflammation. Nat. Protoc. 2007, 2, 541–546. [Google Scholar] [CrossRef]
- Wirtz, S.; Neurath, M.F. Mouse models of inflammatory bowel disease. Adv. Drug. Deliv. Rev. 2007, 59, 1073–1083. [Google Scholar] [CrossRef]
- Mizoguchi, A.; Mizoguchi, E. Animal models of IBD: Linkage to human disease. Curr. Opin. Pharmacol. 2010, 10, 578–587. [Google Scholar] [CrossRef] [Green Version]
- Anka Idrissi, D.; Senhaji, N.; Aouiss, A.; Khalki, L.; Tijani, Y.; Zaid, N.; Marhoume, F.Z.; Naya, A.; Oudghiri, M.; Kabine, M.; et al. IL-1 and CD40/CD40L platelet complex: Elements of induction of Crohn’s disease and new therapeutic targets. Arch. Pharm. Res. 2021, 44, 117–132. [Google Scholar] [CrossRef]
- Dong, F.; Zhang, L.; Hao, F.; Tang, H.; Wang, Y. Systemic responses of mice to dextran sulfate sodium-induced acute ulcerative colitis using 1H NMR spectroscopy. J. Proteome Res. 2013, 12, 2958–2966. [Google Scholar] [CrossRef]
- Schicho, R.; Nazyrova, A.; Shaykhutdinov, R.; Duggan, G.; Vogel, H.J.; Storr, M. Quantitative metabolomic profiling of serum and urine in DSS-induced ulcerative colitis of mice by 1H NMR spectroscopy. J. Proteome Res. 2010, 9, 6265–6273. [Google Scholar] [CrossRef]
- Neurath, M.F.; Fuss, I.; Kelsall, B.L.; Stüber, E.; Strober, W. Antibodies to interleukin 12 abrogate established experimental colitis in mice. J. Exp. Med. 1995, 182, 1281–1290. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, M.; Takeshima, F.; Isomoto, H.; Shikuwa, S.; Mizuta, Y.; Ozono, Y.; Kohno, S. Simvastatin attenuates trinitrobenzene sulfonic acid-induced colitis, but not oxazalone-induced colitis. Dig. Dis. Sci. 2008, 53, 1869–1875. [Google Scholar] [CrossRef] [Green Version]
- Qin, W.; Li, L.; Wang, T.; Huang, H.; Gao, Y. Urine Proteome Changes in a TNBS-Induced Colitis Rat Model. Proteom. Clin. Appl. 2019, 13, e1800100. [Google Scholar] [CrossRef]
- Zhang, X.; Choi, F.F.K.; Zhou, Y.; Leung, F.P.; Tan, S.; Lin, S.; Xu, H.; Jia, W.; Sung, J.J.; Cai, Z.; et al. Metabolite profiling of plasma and urine from rats with TNBS-induced acute colitis using UPLC-ESI-QTOF-MS-based metabonomics—A pilot study. FEBS J. 2012, 279, 2322–2338. [Google Scholar] [CrossRef]
- Khor, B.; Gardet, A.; Xavier, R.J. Genetics and pathogenesis of inflammatory bowel disease. Nature 2011, 474, 307–317. [Google Scholar] [CrossRef] [Green Version]
- Tso, V.K.; Sydora, B.C.; Foshaug, R.R.; Churchill, T.A.; Doyle, J.; Slupsky, C.M.; Fedorak, R.N. Metabolomic Profiles Are Gender, Disease and Time Specific i the Interleukin-10 Gene-Deficient Mouse Model of Inflammatory Bowel Disease. PLoS ONE 2013, 8, e67654. [Google Scholar] [CrossRef]
- Lin, H.M.; Edmunds, S.J.; Helsby, N.A.; Ferguson, L.R.; Rowan, D.D. Nontargeted Urinary Metabolite Profiling of a Mouse Model of Crohn ’ s Disease research articles. J. Proteome Res. 2009, 8, 2045–2057. [Google Scholar] [CrossRef]
- Lin, H.M.; Barnett, M.P.G.; Roy, N.C.; Joyce, N.I.; Zhu, S.; Armstrong, K.; Helsby, N.A.; Ferguson, L.R.; Rowan, D.D. Metabolomic analysis identifies inflammatory and noninflammatory metabolic effects of genetic modification in a mouse model of Crohn´s disease. J. Proteome Res. 2010, 9, 1965–1975. [Google Scholar] [CrossRef]
- Otter, D.; Cao, M.; Lin, H.M.; Fraser, K.; Edmunds, S.; Lane, G.; Rowan, D. Identification of urinary biomarkers of colon inflammation in IL10−/− mice using short-column LCMS metabolomics. J. Biomed. Biotechnol. 2011, 2011, 974701. [Google Scholar] [CrossRef] [Green Version]
- Murdoch, T.B.; Fu, H.; MacFarlane, S.; Sydora, B.C.; Fedorak, R.N.; Slupsky, C.M. Urinary metabolic profiles of inflammatory bowel disease in interleukin-10 gene-deficient mice. Anal. Chem. 2008, 8, 5524–5531. [Google Scholar] [CrossRef]
- Ryan, D.; Robards, K.; Prenzler, P.D.; Kendall, M. Recent and potential developments in the analysis of urine: A review. Anal. Chim. Acta. 2011, 684, 17–29. [Google Scholar] [CrossRef]
- Maráková, K.; Piešťanský, J.; Zelinková, Z.; Mikuš, P. Simultaneous determination of twelve biogenic amines in human urine as potential biomarkers of inflammatory bowel diseases by capillary electrophoresis—Tandem mass spectrometry. J. Pharm. Biomed. Anal. 2020, 186, 113294. [Google Scholar] [CrossRef]
- Stephens, N.S.; Siffledeen, J.; Su, X.; Murdoch, T.B.; Fedorak, R.N.; Slupsky, C.M. Urinary NMR metabolomic profiles discriminate inflammatory bowel disease from healthy. J. Crohns Colitis 2013, 7, e42–e48. [Google Scholar] [CrossRef] [Green Version]
- Schicho, R.; Shaykhutdinov, R.; Ngo, J.; Nazyrova, A.; Schneider, C.; Panaccione, R.; Kaplan, G.G.; Vogel, H.J.; Storr, M. Quantitative metabolomic profiling of serum, plasma, and urine by 1H NMR spectroscopy discriminates between patients with inflammatory bowel disease and healthy individuals. J. Proteome Res. 2012, 11, 3344–3357. [Google Scholar] [CrossRef]
- Cracowski, J.L.; Bonaz, B.; Bessard, G.; Bessard, J.; Anglade, C.; Fournet, J. Increased urinary F2-isoprostanes in patients with Crohn’s disease. Am. J. Gastroenterol. 2002, 97, 99–103. [Google Scholar] [CrossRef]
- Williams, H.R.T.; Cox, I.J.; Walker, D.G.; North, B.V.; Patel, V.M.; Marshall, S.E.; Jewell, D.P.; Ghosh, S.; Thomas, H.J.; Teare, J.P.; et al. Characterization of inflammatory bowel disease with urinary metabolic profiling. Am. J. Gastroenterol. 2009, 104, 1435–1444. [Google Scholar] [CrossRef]
- Martin, F.P.; Ezri, J.; Cominetti, O.; Da Silva, L.; Kussmann, M.; Godin, J.P.; Nydegger, A. Urinary Metabolic Phenotyping Reveals Differences in the Metabolic Status of Healthy and Inflammatory Bowel Disease (IBD) Children in Relation to Growth and Disease Activity. Int. J. Mol. Sci. 2016, 17, 1310. [Google Scholar] [CrossRef] [Green Version]
- Martin, F.P.; Su, M.M.; Xie, G.X.; Guiraud, S.P.; Kussmann, M.; Godin, J.P.; Jia, W.; Nydegger, A. Urinary metabolic insights into host-gut microbial interactions in healthy and IBD children. World J. Gastroenterol. 2017, 23, 3643–3654. [Google Scholar] [CrossRef]
- El Hassani, S.E.M.; Bosch, S.; Lemmen, J.P.M.; Brizzio Brentar, M.; Ayada, I.; Wicaksono, A.N.; Covington, J.A.; Benninga, M.A.; de Boer, N.K.H.; de Meij, T.G.J. Simultaneous assessment of urinary and fecal volatile organic compound analysis in de novo pediatric IBD. Sensors 2019, 19, 4496. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Shanmuganathan, M.; Hart, L.; Pai, N.; Britz-Mckibbin, P. Urinary Metabolites Enable Differential Diagnosis and Therapeutic Monitoring of Pediatric Inflammatory Bowel Disease. Metabolites 2021, 11, 245. [Google Scholar] [CrossRef]
- Siebert, S.; Porter, D.; Paterson, C.; Hampson, R.; Gaya, D.; Latosinska, A.; Mischak, H.; Schanstra, J.; Mullen, W.; McInnes, I. Urinary proteomics can define distinct diagnostic inflammatory arthritis subgroups. Sci. Rep. 2017, 7, 40473. [Google Scholar] [CrossRef] [Green Version]
- Alonso, A.; Julià, A.; Vinaixa, M.; Domènech, E.; Fernández-Nebro, A.; Cañete, J.D.; Ferrándiz, C.; Tornero, J.; Gisbert, J.P.; Nos, P.; et al. Urine metabolome profiling of immune-mediated inflammatory diseases. BMC Med. 2016, 14, 133. [Google Scholar] [CrossRef] [Green Version]
- Keshteli, A.H.; Madsen, K.L.; Mandal, R.; Boeckxstaens, G.E.; Bercik, P.; De Palma, G.; Reed, D.E.; Wishart, D.; Vanner, S.; Dieleman, L.A. Comparison of the metabolomic profiles of irritable bowel syndrome patients with ulcerative colitis patients and healthy controls: New insights into pathophysiology and potential biomarkers. Aliment. Pharmacol. Ther. 2019, 49, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Dawiskiba, T.; Deja, S.; Mulak, A.; Ząbek, A.; Jawień, E.; Pawełka, D.; Banasik, M.; Mastalerz-Migas, A.; Balcerzak, W.; Kaliszewski, K.; et al. Serum and urine metabolomic fngerprinting in diagnostics of inflammatory bowel diseases. World J. Gastroenterol. 2014, 20, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Markó, L.; Szigeti, N.; Szabó, Z.; Takátsy, A.; Ludány, A.; Koszegi, T.; Molnár, G.A.; Wittmann, I. Potential urinary biomarkers of disease activity in Crohn’s disease. Scand. J. Gastroenterol. 2010, 45, 1440–1448. [Google Scholar] [CrossRef]
- Bjerrum, J.T.; Nielsen, O.H.; Hao, F.; Tang, H.; Nicholson, J.K.; Wang, Y.; Olsen, J. Metabonomics in ulcerative colitis: Diagnostics, biomarker identification, and insight into the pathophysiology. J. Proteome Res. 2010, 9, 954–962. [Google Scholar] [CrossRef]
- Stanke-Labesque, F.; Pofelski, J.; Moreau-Gaudry, A.; Bessard, G.; Bonaz, B. Urinary leukotriene E4 excretion: A biomarker of inflammatory bowel disease activity. Inflamm. Bowel Dis. 2008, 14, 769–774. [Google Scholar] [CrossRef]
- Keshteli, A.H.; Tso, R.; Dieleman, L.A.; Park, H.; Kroeker, K.I.; Jovel, J.; Gillevet, P.M.; Sikarood, M.; Mandal, R.; Fedorak, R.N.; et al. A Distinctive Urinary Metabolomic Fingerprint Is Linked With Endoscopic Postoperative Disease Recurrence in Crohn’s Disease Patients. Inflamm. Bowel Dis. 2018, 24, 861–870. [Google Scholar] [CrossRef]
- Keshteli, A.H.; Den Brand, F.F.V.; Madsen, K.L.; Park, H.; Kroeker, K.I.; Jovel, J.; Gillevet, P.M.; Sikaroodi, M.; Mandal, R.; Fedorak, R.N.; et al. Dietary and metabolomic determinants of relapse in ulcerative colitis patients: A pilot prospective cohort study. World J. Gastroenterol. 2017, 23, 3890–3899. [Google Scholar] [CrossRef]
- Julian, B.A.; Suzuki, H.; Suzuki, Y.; Tomino, Y.; Spasovski, G.; Novak, J. Sources of Urinary Proteins and their Analysis by Urinary Proteomics for the Detection of Biomarkers of Disease. Proteom. Clin. Appl. 2009, 3, 1029. [Google Scholar] [CrossRef] [Green Version]
- Rotter, M.; Brandmaier, S.; Prehn, C.; Adam, J.; Rabstein, S.; Gawrych, K.; Brüning, T.; Illig, T.; Lickert, H.; Adamski, J.; et al. Stability of targeted metabolite profiles of urine samples under different storage conditions. Metabolomics 2017, 13, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolscheid-Pommerich, R.C.; Kiarmann-Schuiz, U.; Conrad, R.; Stoffel-Wagner, B.; Zur, B. Evaluation of the appropriate time period between sampling and analyzing for automated urinalysis. Biochem. Med. 2016, 26, 82–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldan-Martin, M.; Chaparro, M.; Gisbert, J.P. Systematic Review: Urine Biomarker Discovery for Inflammatory Bowel Disease Diagnosis. Int. J. Mol. Sci. 2023, 24, 10159. https://doi.org/10.3390/ijms241210159
Baldan-Martin M, Chaparro M, Gisbert JP. Systematic Review: Urine Biomarker Discovery for Inflammatory Bowel Disease Diagnosis. International Journal of Molecular Sciences. 2023; 24(12):10159. https://doi.org/10.3390/ijms241210159
Chicago/Turabian StyleBaldan-Martin, Montse, María Chaparro, and Javier P. Gisbert. 2023. "Systematic Review: Urine Biomarker Discovery for Inflammatory Bowel Disease Diagnosis" International Journal of Molecular Sciences 24, no. 12: 10159. https://doi.org/10.3390/ijms241210159