
Optimization of Mass Spectrometry Imaging for Drug Metabolism and Distribution Studies in the Zebrafish Larvae Model: A Case Study with the Opioid Antagonist Naloxone


Abstract
:
1. Introduction
2. Results and Discussion
2.1. In Vivo Absorption, Metabolism, and Excretion Properties of Naloxone in the Zebrafish Larvae Model
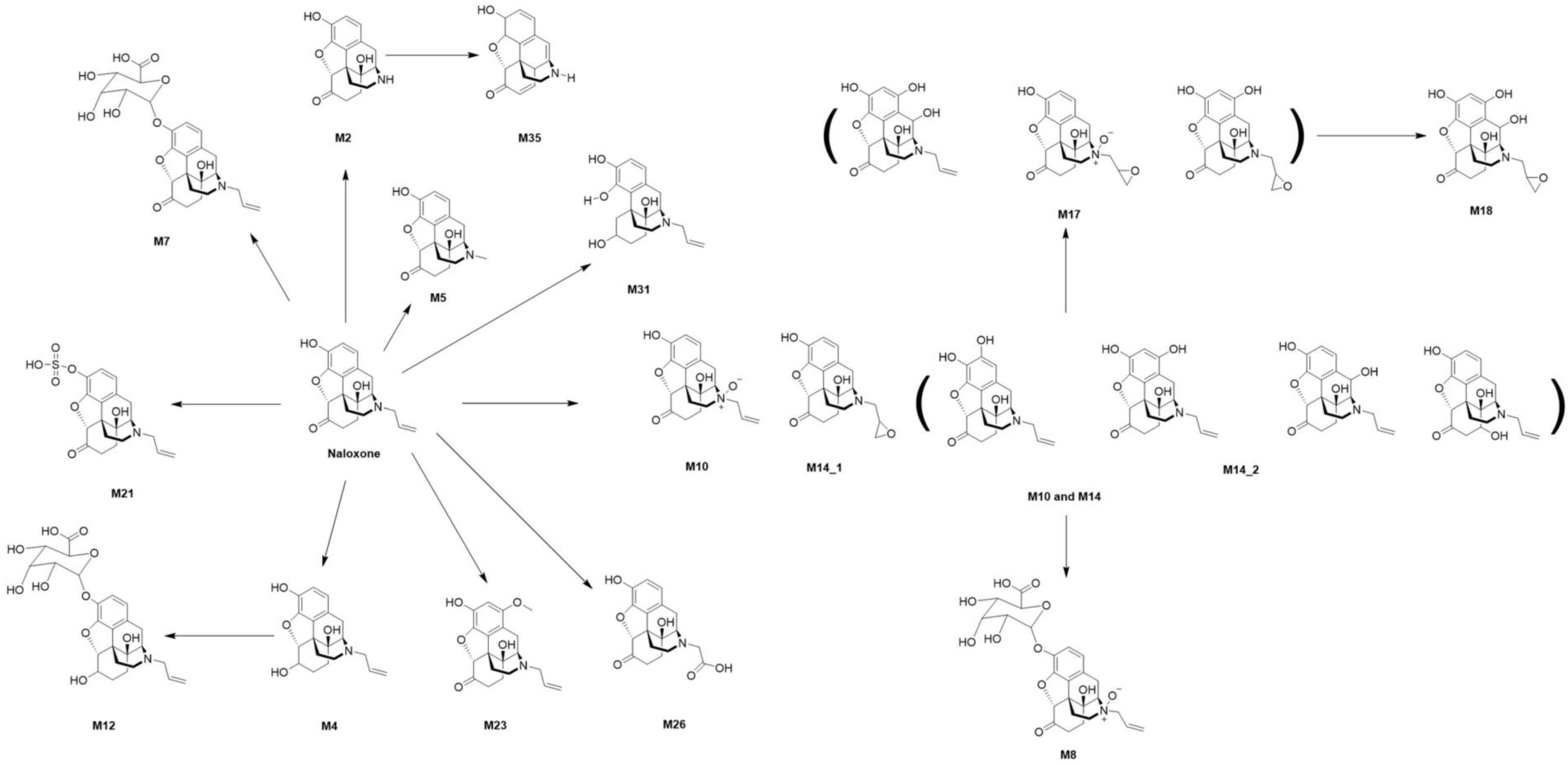
2.2. Localization of Naloxone and Its Metabolites in Whole-Body Sections of Zebrafish Larvae
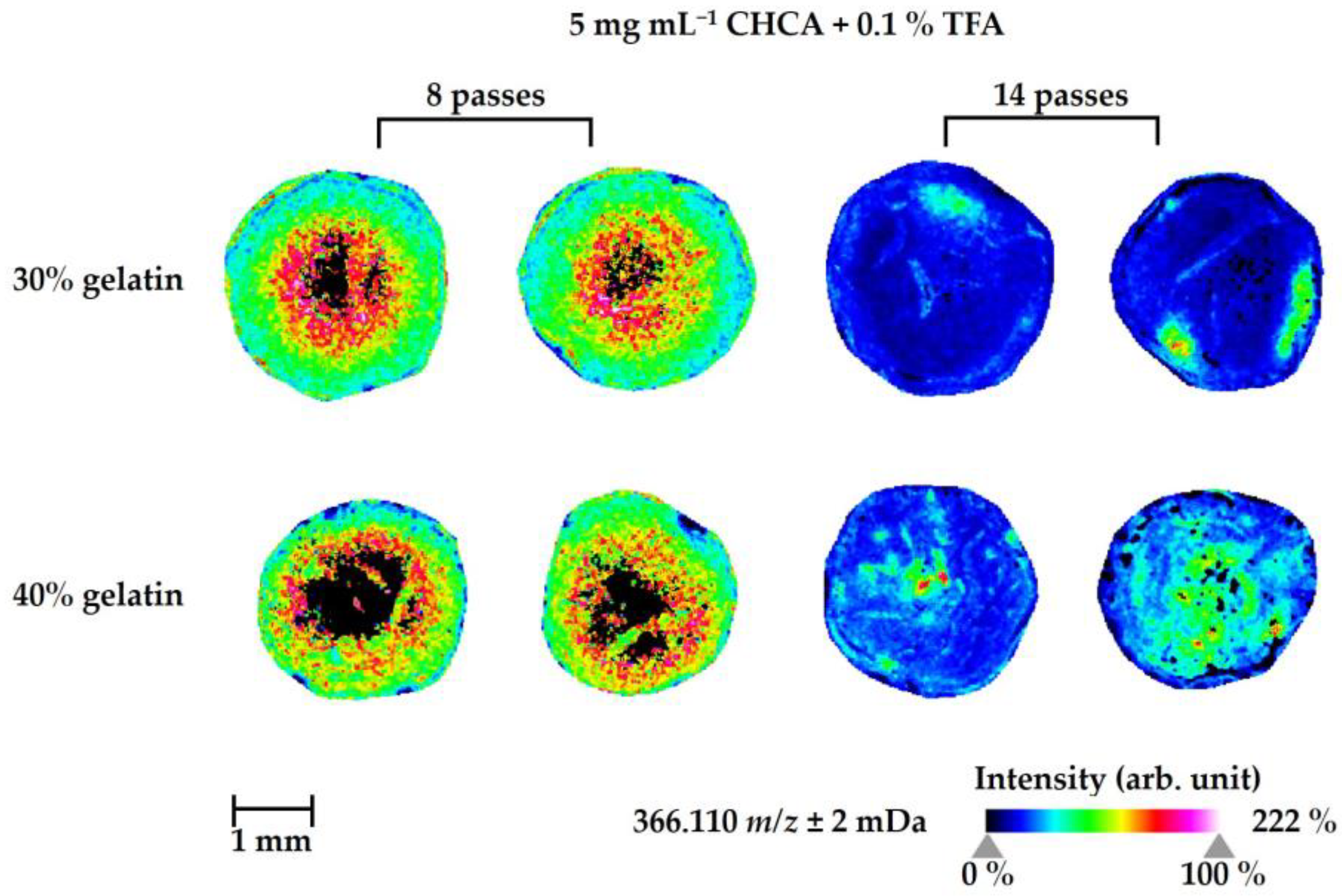
2.3. Optimization of MSI Methods to Study the Drug Distribution in Zebrafish Larvae—A Case Study Using Naloxone
3. Materials and Methods
3.1. Chemicals and Other Materials
3.2. ZF Maintenance and Embryo Collection
3.3. Drug Treatment of ZF Larvae via Medium Exposure
3.4. Manual Sectioning of ZF Larvae
3.5. ZF Sample Preparation for Metabolite Analysis by LC-HRMS/MS
3.6. Mass Spectrometry Imaging Analysis of Naloxone and Its Metabolites by MALDI-FT-ICR
3.7. ZF Larval Homogenate Preparation
3.8. In Vitro Metabolism Analyses Using HepaRG Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davoli, E.; Zucchetti, M.; Matteo, C.; Ubezio, P.; D’Incalci, M.; Morosi, L. The Space Dimension at the Micro Level: Mass Spectrometry Imaging of Drugs in Tissues. Mass Spec. Rev. 2021, 40, 201–214. [Google Scholar]
- Buchberger, A.R.; DeLaney, K.; Johnson, J.; Li, L. Mass Spectrometry Imaging: A Review of Emerging Advancements and Future Insights. Anal. Chem. 2018, 90, 240–265. [Google Scholar] [CrossRef]
- Zemaitis, K.J.; Izydorczak, A.M.; Thompson, A.C.; Wood, T.D. Streamlined Multimodal DESI and MALDI Mass Spectrometry Imaging on a Singular Dual-Source FT-ICR Mass Spectrometer. Metabolites 2021, 11, 253. [Google Scholar] [CrossRef]
- Gorzolka, K.; Kölling, J.; Nattkemper, T.W.; Niehaus, K. Spatio-Temporal Metabolite Profiling of the Barley Germination Process by MALDI MS Imaging. PLoS ONE 2016, 11, e0150208. [Google Scholar] [CrossRef]
- Ntshangase, S.; Mdanda, S.; Naicker, T.; Kruger, H.G.; Baijnath, S.; Govender, T. Spatial distribution of elvitegravir and tenofovir in rat brain tissue: Application of matrix-assisted laser desorption/ionization mass spectrometry imaging and liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2019, 33, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Chen, J.; Zhou, J.; Ge, J.; Zhang, Y.; Li, P.; Li, B. Quantitative MALDI Imaging of Spatial Distributions and Dynamic Changes of Tetrandrine in Multiple Organs of Rats. Theranostics 2019, 9, 932–944. [Google Scholar] [CrossRef]
- Giordano, S.; Pifferi, V.; Morosi, L.; Morelli, M.; Falciola, L.; Cappelletti, G.; Visentin, S.; Licandro, S.A.; Frapolli, R.; Zucchetti, M.; et al. A Nanostructured Matrices Assessment to Study Drug Distribution in Solid Tumor Tissues by Mass Spectrometry Imaging. Nanomaterials 2017, 7, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, C.; Jang, M.; de Jesus, J.; Steven, R.T.; Nikula, C.J.; Elia, E.; Bunch, J.; Bellew, A.T.; Watts, J.F.; Hinder, S.; et al. Imaging mass spectrometry: A new way to distinguish dermal contact from administration of cocaine, using a single fingerprint. Analyst 2021, 146, 4010–4021. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.J.; Bradshaw, R.; Francese, S.; Salter, T.L.; Costa, C.; Ismail, M.; Webb, R.P.; Bosman, I.; Wolff, K.; de Puit, M. Rapid detection of cocaine, benzoylecgonine and methylecgonine in fingerprints using surface mass spectrometry. Analyst 2015, 140, 6254–6259. [Google Scholar] [CrossRef] [Green Version]
- Ifa, D.R.; Manicke, N.E.; Dill, A.L.; Cooks, R.G. Latent fingerprint chemical imaging by mass spectrometry. Science 2008, 321, 805. [Google Scholar] [CrossRef] [Green Version]
- Lawson, N.D.; Weinstein, B.M. In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev. Biol. 2002, 248, 307–318. [Google Scholar] [CrossRef] [Green Version]
- Carten, J.D.; Bradford, M.K.; Farber, S.A. Visualizing digestive organ morphology and function using differential fatty acid metabolism in live zebrafish. Dev. Biol. 2011, 360, 276–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasheed, S.; Fries, F.; Müller, R.; Herrmann, J. Zebrafish: An Attractive Model to Study Staphylococcus aureus Infection and Its Use as a Drug Discovery Tool. Pharmaceuticals 2021, 14, 594. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ji, C.; Yan, L.; Lu, M.; Lu, C.; Zhao, M. The identification of the metabolites of chlorothalonil in zebrafish (Danio rerio) and their embryo toxicity and endocrine effects at environmentally relevant levels. Environ. Pollut. 2016, 218, 8–15. [Google Scholar] [PubMed]
- Wiegand, C.; Krause, E.; Steinberg, C.; Pflugmacher, S. Toxicokinetics of atrazine in embryos of the zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2001, 49, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Barros, T.P.; Alderton, W.K.; Reynolds, H.M.; Roach, A.G.; Berghmans, S. Zebrafish: An emerging technology for in vivo pharmacological assessment to identify potential safety liabilities in early drug discovery. Br. J. Pharmacol. 2008, 154, 1400–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papan, C.; Chen, L. Metabolic fingerprinting reveals developmental regulation of metabolites during early zebrafish embryogenesis. Omics J. Integr. Biol. 2009, 13, 397–405. [Google Scholar] [CrossRef]
- Chng, H.T.; Ho, H.K.; Yap, C.W.; Lam, S.H.; Chan, E.C.Y. An investigation of the bioactivation potential and metabolism profile of Zebrafish versus human. J. Biomol. Screen. 2012, 17, 974–986. [Google Scholar] [CrossRef] [Green Version]
- Jones, H.S.; Trollope, H.T.; Hutchinson, T.H.; Panter, G.H.; Chipman, J.K. Metabolism of ibuprofen in zebrafish larvae. Xenobiotica Fate Foreign Compd. Biol. Syst. 2012, 42, 1069–1075. [Google Scholar] [CrossRef]
- van Wijk, R.C.; Krekels, E.H.J.; Hankemeier, T.; Spaink, H.P.; van der Graaf, P.H. Systems pharmacology of hepatic metabolism in zebrafish larvae. Drug Discov. Today Dis. Model. 2016, 22, 27–34. [Google Scholar] [CrossRef]
- Wu, J.-Q.; Fan, R.-Y.; Zhang, S.-R.; Li, C.-Y.; Shen, L.-Z.; Wei, P.; He, Z.-H.; He, M.-F. A systematical comparison of anti-angiogenesis and anti-cancer efficacy of ramucirumab, apatinib, regorafenib and cabozantinib in zebrafish model. Life Sci. 2020, 247, 117402. [Google Scholar] [PubMed]
- Meijer, A.H. Protection and pathology in TB: Learning from the zebrafish model. Semin. Immunopathol. 2016, 38, 261–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shams, S.; Rihel, J.; Ortiz, J.G.; Gerlai, R. The zebrafish as a promising tool for modeling human brain disorders: A review based upon an IBNS Symposium. Neurosci. Biobehav. Rev. 2018, 85, 176–190. [Google Scholar] [CrossRef] [PubMed]
- Martín-Jiménez, R.; Campanella, M.; Russell, C. New zebrafish models of neurodegeneration. Curr. Neurol. Neurosci. Rep. 2015, 15, 33. [Google Scholar]
- Gonzalez-Nunez, V.; Rodríguez, R.E. The zebrafish: A model to study the endogenous mechanisms of pain. ILAR J. 2009, 50, 373–386. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.M.; Meyer, M.R.; Müller, R.; Herrmann, J. Drug Administration Routes Impact the Metabolism of a Synthetic Cannabinoid in the Zebrafish Larvae Model. Molecules 2020, 25, 4474. [Google Scholar] [CrossRef]
- Park, Y.M.; Dahlem, C.; Meyer, M.R.; Kiemer, A.K.; Müller, R.; Herrmann, J. Induction of Liver Size Reduction in Zebrafish Larvae by the Emerging Synthetic Cannabinoid 4F-MDMB-BINACA and Its Impact on Drug Metabolism. Molecules 2022, 27, 1290. [Google Scholar] [CrossRef]
- Gampfer, T.M.; Wagmann, L.; Park, Y.M.; Cannaert, A.; Herrmann, J.; Fischmann, S.; Westphal, F.; Müller, R.; Stove, C.P.; Meyer, M.R. Toxicokinetics and toxicodynamics of the fentanyl homologs cyclopropanoyl-1-benzyl-4′-fluoro-4-anilinopiperidine and furanoyl-1-benzyl-4-anilinopiperidine. Arch. Toxicol. 2020, 94, 2009–2025. [Google Scholar] [CrossRef] [Green Version]
- Wagmann, L.; Frankenfeld, F.; Park, Y.M.; Herrmann, J.; Fischmann, S.; Westphal, F.; Müller, R.; Flockerzi, V.; Meyer, M.R. How to Study the Metabolism of New Psychoactive Substances for the Purpose of Toxicological Screenings-A Follow-Up Study Comparing Pooled Human Liver S9, HepaRG Cells, and Zebrafish Larvae. Front. Chem. 2020, 8, 539. [Google Scholar] [CrossRef]
- Richter, L.H.J.; Herrmann, J.; Andreas, A.; Park, Y.M.; Wagmann, L.; Flockerzi, V.; Müller, R.; Meyer, M.R. Tools for studying the metabolism of new psychoactive substances for toxicological screening purposes—A comparative study using pooled human liver S9, HepaRG cells, and zebrafish larvae. Toxicol. Lett. 2019, 305, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Kirla, K.T.; Groh, K.J.; Steuer, A.E.; Poetzsch, M.; Banote, R.K.; Stadnicka-Michalak, J.; Eggen, R.I.L.; Schirmer, K.; Kraemer, T. From the Cover: Zebrafish Larvae Are Insensitive to Stimulation by Cocaine: Importance of Exposure Route and Toxicokinetics. Toxicol. Sci. 2016, 154, 183–193. [Google Scholar] [CrossRef]
- Nguyen, D.D.; Saharuka, V.; Kovalev, V.; Stuart, L.; Del Prete, M.; Lubowiecka, K.; de Mot, R.; Venturi, V.; Alexandrov, T. Facilitating Imaging Mass Spectrometry of Microbial Specialized Metabolites with METASPACE. Metabolites 2021, 11, 477. [Google Scholar] [CrossRef] [PubMed]
- Abdelmoula, W.M.; Pezzotti, N.; Hölt, T.; Dijkstra, J.; Vilanova, A.; McDonnell, L.A.; Lelieveldt, B.P.F. Interactive Visual Exploration of 3D Mass Spectrometry Imaging Data Using Hierarchical Stochastic Neighbor Embedding Reveals Spatiomolecular Structures at Full Data Resolution. J. Proteome Res. 2018, 17, 1054–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuso Nerini, I.; Morosi, L.; Zucchetti, M.; Ballerini, A.; Giavazzi, R.; D’Incalci, M. Intratumor heterogeneity and its impact on drug distribution and sensitivity. Clin. Pharmacol. Ther. 2014, 96, 224–238. [Google Scholar] [CrossRef]
- Jove, M.; Spencer, J.; Clench, M.; Loadman, P.M.; Twelves, C. Precision pharmacology: Mass spectrometry imaging and pharmacokinetic drug resistance. Crit. Rev. Oncol. Hematol. 2019, 141, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Huizing, L.R.S.; McDuffie, J.; Cuyckens, F.; van Heerden, M.; Koudriakova, T.; Heeren, R.M.A.; Vreeken, R.J. Quantitative Mass Spectrometry Imaging to Study Drug Distribution in the Intestine Following Oral Dosing. Anal. Chem. 2021, 93, 2144–2151. [Google Scholar] [CrossRef] [PubMed]
- Villacrez, M.; Hellman, K.; Ono, T.; Sugihara, Y.; Rezeli, M.; Ek, F.; Marko-Varga, G.; Olsson, R. Evaluation of Drug Exposure and Metabolism in Locust and Zebrafish Brains Using Mass Spectrometry Imaging. ACS Chem. Neurosci. 2018, 9, 1994–2000. [Google Scholar] [CrossRef]
- Nelson, K.A.; Daniels, G.J.; Fournie, J.W.; Hemmer, M.J. Optimization of whole-body zebrafish sectioning methods for mass spectrometry imaging. J. Biomol. Tech. 2013, 24, 119–127. [Google Scholar] [CrossRef]
- Fujimoto, J.M. Isolation of naloxone-3-glucuronide from human urine. Proceedings of the Society for Experimental Biology and Medicine. Soc. Exp. Biol. Med. 1970, 133, 317–319. [Google Scholar] [CrossRef]
- Weinstein, S.H.; Pfeffer, M.; Schor, J.M.; Indindoli, L.; Mintz, M. Metabolites of naloxone in human urine. J. Pharm. Sci. 1971, 60, 1567–1568. [Google Scholar]
- van Dorp, E.L.A.; Yassen, A.; Dahan, A. Naloxone treatment in opioid addiction: The risks and benefits. Expert Opin. Drug Saf. 2007, 6, 125–132. [Google Scholar] [CrossRef]
- Lowenthal, R.E. FDA Advisory Committee on the Most Appropriate Dose or Doses of Naloxone to Reverse the Effects of Life-Threatening Opioid Overdose in the Community Settings; Joint Meeting of the Anesthetic and Analgesic Drug Products and the Drug Safety and Risk Management Advisory Committees on 5 October, 2016 in Silver Spring, MD, USA. Advisory Committee Briefing Materials, 2 September 2016. Available online: https://fda.report/media/100409/Adapt-Pharma-Briefing-Information-for-the-October-5--2016-Joint-Meeting-of-the-Anesthetic-and-Analgesic-Drug-Products-Advisory-Committee-and-the-Drug-Safety-and-Risk-Management-Advisory-Com.pdf (accessed on 2 July 2021).
- Advisory Committee Briefing Materials. Joint Meeting of the Anesthetic and Analgesic Drug Products and the Drug Safety and Risk Management Advisory Committees on 5 October, 2016 in Silver Spring, MD, USA. Kaleo, Inc. Briefing Dossier, 1 September 2016. Available online: https://www.fda.gov/media/100440/download (accessed on 8 June 2021).
- Rzasa Lynn, R.; Galinkin, J.L. Naloxone dosage for opioid reversal: Current evidence and clinical implications. Ther. Adv. Drug Saf. 2018, 9, 63–88. [Google Scholar] [CrossRef] [Green Version]
- Haidari, M.; Mansani, S.; Ponds, D.; Romero, L.; Alsaab, S. Consumption of Movantik™ (Naloxegol) results in detection of naloxone in the patient’s urine evaluated by confirmatory urine drug testing. Clin. Biochem. 2019, 67, 48–53. [Google Scholar]
- Chiang, C. Pharmacokinetics of the combination tablet of buprenorphine and naloxone. Drug Alcohol Depend. 2003, 70, S39–S47. [Google Scholar] [PubMed]
- Wiegand, S.L.; Swortwood, M.J.; Huestis, M.A.; Thorp, J.; Jones, H.E.; Vora, N.L. Naloxone and Metabolites Quantification in Cord Blood of Prenatally Exposed Newborns and Correlations with Maternal Concentrations. AJP Rep. 2016, 6, e385–e390. [Google Scholar] [CrossRef] [PubMed]
- Swortwood, M.J.; Scheidweiler, K.B.; Barnes, A.J.; Jansson, L.M.; Huestis, M.A. Simultaneous quantification of buprenorphine, naloxone and phase I and II metabolites in plasma and breastmilk by liquid chromatography-tandem mass spectrometry. J. Chromatogr. 2016, 1446, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Misra, A.L.; Pontani, R.B.; Vadlamani, N.L.; Mulé, S.J. Physiological disposition and biotransformation of allyl-1′, 3′—14C naloxone in the rat and some comparative observations on nalorphine. J. Pharmacol. Exp. Ther. 1976, 196, 257–268. [Google Scholar]
- European Medicines Agency. Assessment Report: Nyxoid; EMEA/H/C/004325/0000; European Medicines Agency: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Joshi, A.; Parris, B.; Liu, Y.; Heidbreder, C.; Gerk, P.M.; Halquist, M. Quantitative determination of buprenorphine, naloxone and their metabolites in rat plasma using hydrophilic interaction liquid chromatography coupled with tandem mass spectrometry. Biomed. Chromatogr. BMC 2017, 31, e3785. [Google Scholar]
- Garrett, E.R.; Shyu, W.C.; Ulubelen, A. Pharmacokinetics of morphine and its surrogates. VIII: Naloxone and naloxone conjugate pharmacokinetics in dogs as a function of dose and as affected by simultaneously administered morphine. J. Pharm. Sci. 1986, 75, 1127–1136. [Google Scholar]
- UNODC. World Drug Report 2021: Booklet 5—COVID-19 and Drugs: Impact and Outlook; United Nations Office on Drugs and Crime: Vienna, Austria, 2021. [Google Scholar]
- Teklezgi, B.G.; Pamreddy, A.; Ntshangase, S.; Mdanda, S.; Singh, S.D.; Gopal, N.D.; Naicker, T.; Kruger, H.G.; Govender, T.; Baijnath, S. Mass Spectrometric Imaging of the Brain Demonstrates the Regional Displacement of 6-Monoacetylmorphine by Naloxone. ACS Omega 2020, 5, 12596–12602. [Google Scholar] [CrossRef]
- Asslan, M.; Lauzon, N.; Beus, M.; Maysinger, D.; Rousseau, S. Mass spectrometry imaging in zebrafish larvae for assessing drug safety and metabolism. Anal. Bioanal. Chem. 2021, 413, 5135–5146. [Google Scholar] [CrossRef]
- Hulme, H.; Fridjonsdottir, E.; Gunnarsdottir, H.; Vallianatou, T.; Zhang, X.; Wadensten, H.; Shariatgorji, R.; Nilsson, A.; Bezard, E.; Svenningsson, P.; et al. Simultaneous mass spectrometry imaging of multiple neuropeptides in the brain and alterations induced by experimental parkinsonism and L-DOPA therapy. Neurobiol. Dis. 2020, 137, 104738. [Google Scholar] [CrossRef]
- Balluff, B.; Hopf, C.; Porta Siegel, T.; Grabsch, H.I.; Heeren, R.M.A. Batch Effects in MALDI Mass Spectrometry Imaging. J. Am. Soc. Mass Spectrom. 2021, 32, 628–635. [Google Scholar] [CrossRef]
- Zecchi, R.; Franceschi, P.; Tigli, L.; Amidani, D.; Catozzi, C.; Ricci, F.; Salomone, F.; Pieraccini, G.; Pioselli, B.; Mileo, V. Sample preparation strategy for the detection of steroid-like compounds using MALDI mass spectrometry imaging: Pulmonary distribution of budesonide as a case study. Anal. Bioanal. Chem. 2021, 413, 4363–4371. [Google Scholar] [CrossRef]
- Hansen, H.T.; Janfelt, C. Aspects of Quantitation in Mass Spectrometry Imaging Investigated on Cryo-Sections of Spiked Tissue Homogenates. Anal. Chem. 2016, 88, 11513–11520. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.J.; Dexter, A.; Bunch, J. Exploring Ion Suppression in Mass Spectrometry Imaging of a Heterogeneous Tissue. Anal. Chem. 2018, 90, 5637–5645. [Google Scholar] [CrossRef] [PubMed]
- Lanekoff, I.; Stevens, S.L.; Stenzel-Poore, M.P.; Laskin, J. Matrix effects in biological mass spectrometry imaging: Identification and compensation. Analyst 2014, 139, 3528–3532. [Google Scholar] [CrossRef] [Green Version]
- Barry, J.A.; Groseclose, M.R.; Castellino, S. Quantification and assessment of detection capability in imaging mass spectrometry using a revised mimetic tissue model. Bioanalysis 2019, 11, 1099–1116. [Google Scholar] [CrossRef] [PubMed]
- Angel, P.M.; Caprioli, R.M. Matrix-assisted laser desorption ionization imaging mass spectrometry: In situ molecular mapping. Biochemistry 2013, 52, 3818–3828. [Google Scholar] [CrossRef] [Green Version]
- Zaima, N.; Yoshioka, S.; Sato, Y.; Shinano, S.; Ikeda, Y.; Moriyama, T. Enhanced specificity for phosphatidylcholine analysis by positive ion mode matrix-assisted laser desorption/ionization imaging mass spectrometry. Rapid Commun. Mass Spectrom. RCM 2014, 28, 1453–1458. [Google Scholar] [CrossRef]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio rerio), 4th ed.; University of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Human [39,40,41,42,43,44,45,46,47,48] | Rat [39,43,49,50,51] | Rabbit, Chicken [39] | Dog [43,52] | Data from This Study | |||
|---|---|---|---|---|---|---|---|---|
| HepaRG in Vitro Model † | ZF Larvae ‡ | |||||||
| Plasma | Urine | Plasma, Urine, Feces | Urine | Urine | Larvae | Medium | ||
| Naloxone | + | + | + | + | + | + | + | + |
| M2 | + | + | + | + | + | + | ||
| M4 | + | + | + | + | + | + | + c | |
| M5 | + b | |||||||
| M7 | + | + | + | + | + | + | + | |
| M8 | + c | |||||||
| M9 | + | |||||||
| M10 | + a,b | |||||||
| M12 | + | + d | + c | |||||
| M14 | + a,b | |||||||
| M17 | + e | + b | ||||||
| M18 | + b | |||||||
| M21 | + | + | + | + | ||||
| M23 | + c | |||||||
| M26 | + b | |||||||
| M35 | + | + c | ||||||
| Total number of detected metabolites | 3 | 3 | 7 | 3 | 2 | 4 | 8 | 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, Y.M.; Meyer, M.R.; Müller, R.; Herrmann, J. Optimization of Mass Spectrometry Imaging for Drug Metabolism and Distribution Studies in the Zebrafish Larvae Model: A Case Study with the Opioid Antagonist Naloxone. Int. J. Mol. Sci. 2023, 24, 10076. https://doi.org/10.3390/ijms241210076
Park YM, Meyer MR, Müller R, Herrmann J. Optimization of Mass Spectrometry Imaging for Drug Metabolism and Distribution Studies in the Zebrafish Larvae Model: A Case Study with the Opioid Antagonist Naloxone. International Journal of Molecular Sciences. 2023; 24(12):10076. https://doi.org/10.3390/ijms241210076
Chicago/Turabian StylePark, Yu Mi, Markus R. Meyer, Rolf Müller, and Jennifer Herrmann. 2023. "Optimization of Mass Spectrometry Imaging for Drug Metabolism and Distribution Studies in the Zebrafish Larvae Model: A Case Study with the Opioid Antagonist Naloxone" International Journal of Molecular Sciences 24, no. 12: 10076. https://doi.org/10.3390/ijms241210076