
Mast Cells in Upper and Lower Airway Diseases: Sentinels in the Front Line
 ,
,  , , and
, , and
Abstract
:1. Introduction
1.1. Origin and Development
1.2. Subclasses
1.3. Receptors
1.4. Signal Transduction
1.5. Granules
1.6. Mediators
1.7. Interactions with Other Cells
1.8. Host Defense
1.9. Immunometabolism in Mast Cells
2. Mast Cells in the Airway
2.1. Upper Airways: Allergic Rhinitis
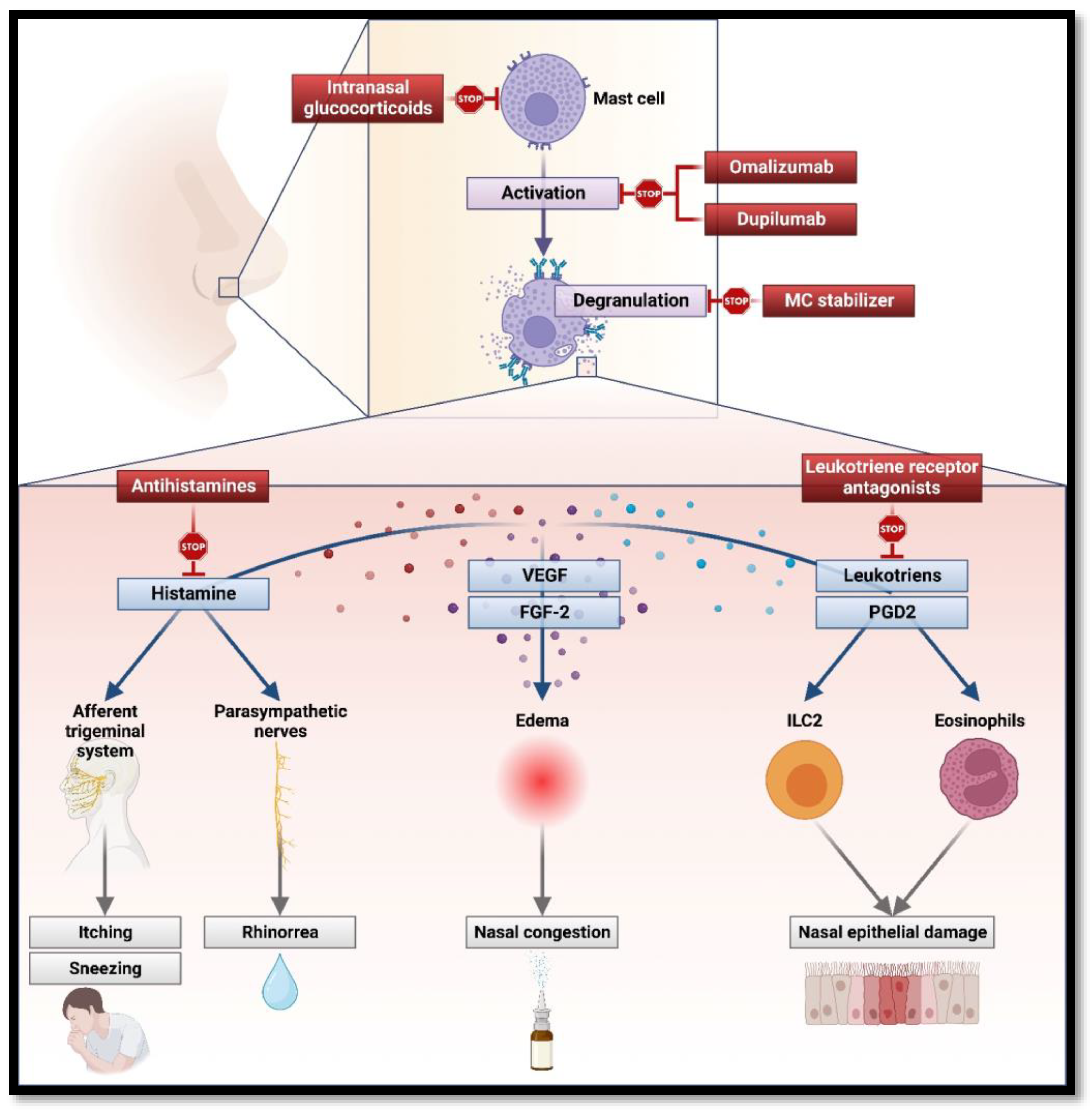
2.1.1. MCs in the Pathogenesis of AR
2.1.2. Therapeutical Perspective on MCs’ Role in AR
2.2. Other Rhinopathies: The Role of Nasal Cytology
2.3. Lower Airways: Asthma
- Variable expiratory airflow limitation, which puts asthma in the group of obstructive respiratory diseases. The major processes that determine this trait are airway inflammation, contraction of airway smooth muscle (ASM), excessive mucus secretion, and mucosal edema due to increased vascular permeability [80].
- Airway/bronchial hyperresponsiveness, maybe the most distinctive trait [81].
- In patients with long-standing asthma, persistent or incompletely reversible airflow limitation due to reduced respiratory muscle function, loss of elastic recoil, and airway wall remodeling. The key elements of this remodeling are denudation of the epithelial layer, goblet cell and mucous gland hyperplasia and hypertrophy, subepithelial fibrosis, abnormal extracellular matrix deposition, vascular proliferation, and increased ASM mass [82].
2.3.1. Expiratory Airflow Limitation
2.3.2. Bronchial Hyperresponsiveness
2.3.3. Airway Remodeling
2.3.4. Other Features
2.3.5. Chronic Activation
2.3.6. Protective Factors
2.3.7. Non-Atopic Asthma

{kind=link}
{kind=link}
| Mediator | Function | Clinical Effect |
|---|---|---|
| Histamine | Bronchoconstriction Mucosal edema Airway remodeling | Bronchospasm Airflow limitation Irreversible obstruction |
| Heparin | Reduce mast cells’ production of pro-inflammatory cytokines | Immunomodulant |
| Chymase | Airway remodeling | Irreversible obstruction |
| Tryptase | Bronchoconstriction | Airflow limitation |
| Serotonin | ASM proliferation | Airflow limitation |
| Il-4 | Immune cell recruitment Airway remodeling | Prolong inflammation and its effects |
| IL-6 | MCs’ proliferation and activation | Prolong inflammation and its effects |
| IL-13 | Mucus production tissue repair Increasing histamine signaling | Airflow limitation Irreversible obstruction |
| TNF-α | Production of pro-inflammatory cytokines Proliferation of airway epithelium | Prolong inflammation and its effects Airflow limitation |
| Chemokines (e.g., CCL1, CCL2, CCL3, CCL4, CCL5, CCL7) | Immune cell recruitment | Prolong inflammation and its effects |
| Growth factors (e.g., VEGF-A, VEGF-C) | Lympho-angiogenesis Airway remodeling | Irreversible obstruction |
2.3.8. Mast-Cell Centered Therapies in Asthma
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Levi-Schaffer, F.; Gibbs, B.F.; Hallgren, J.; Pucillo, C.; Redegeld, F.; Siebenhaar, F.; Vitte, J.; Mezouar, S.; Michel, M.; Puzzovio, P.G.; et al. Selected recent advances in understanding the role of human mast cells in health and disease. J. Allergy Clin. Immunol. 2022, 149, 1833–1844. [Google Scholar] [CrossRef] [PubMed]
- Dahlin, J.S.; Maurer, M.; Metcalfe, D.D.; Pejler, G.; Sagi-Eisenberg, R.; Nilsson, G. The ingenious mast cell: Contemporary insights into mast cell behavior and function. Allergy Eur. J. Allergy Clin. Immunol. 2022, 77, 83–99. [Google Scholar] [CrossRef] [PubMed]
- Aponte-López, A.; Muñoz-Cruz, S. Mast Cells in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1273, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Komi, D.E.A.; Redegeld, F.A. Role of Mast Cells in Shaping the Tumor Microenvironment. Clin. Rev. Allergy Immunol. 2020, 58, 313–325. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, P. Beiträge zur Theorie und Praxis der histologischen Färbung. Collect. Pap. Paul Ehrlich 2013, 29–64. [Google Scholar] [CrossRef]
- Varricchi, G.; Raap, U.; Rivellese, F.; Marone, G.; Gibbs, B.F. Human mast cells and basophils—How are they similar how are they different? Immunol. Rev. 2018, 282, 8–34. [Google Scholar] [CrossRef]
- Franco, C.B.; Chen, C.C.; Drukker, M.; Weissman, I.L.; Galli, S.J. Distinguishing Mast Cell and Granulocyte Differentiation at the Single-Cell Level. Cell Stem Cell 2010, 6, 361–368. [Google Scholar] [CrossRef] [Green Version]
- Kirshenbaum, A. Demostration that human mast cells arise from progenitor cell population that is CD44. J. Immunol. 1991, 94, 2333–2342. [Google Scholar]
- Dahlin, J.S.; Hallgren, J. Mast cell progenitors: Origin, development and migration to tissues. Mol. Immunol. 2015, 63, 9–17. [Google Scholar] [CrossRef]
- Holt, P.G.; Jones, C.A. The development of the immune system during pregnancy and early life. Allergy Eur. J. Allergy Clin. Immunol. 2000, 55, 688–697. [Google Scholar] [CrossRef]
- Bradding, P.; Arthur, G. Mast cells in asthma-state of the art. Clin. Exp. Allergy 2016, 46, 194–263. [Google Scholar] [CrossRef]
- Schuster, C.; Vaculik, C.; Prior, M.; Fiala, C.; Mildner, M.; Eppel, W.; Stingl, G.; Elbe-Bürger, A. Phenotypic characterization of leukocytes in prenatal human dermis. J. Investig. Dermatol. 2012, 132, 2581–2592. [Google Scholar] [CrossRef] [Green Version]
- Msallam, R. Fetal mast cells mediate postnatal allergic responses dependent on maternal IgE. Science 2020, 370, 941–950. [Google Scholar] [CrossRef]
- Murphy, R.C.; Hallstrand, T.S. Exploring the origin and regulatory role of mast cells in asthma. Curr. Opin. Allergy Clin. Immunol. 2021, 21, 71–78. [Google Scholar] [CrossRef]
- Dahlin, J.S.; Ekoff, M.; Grootens, J.; Löf, L.; Amini, R.M.; Hagberg, H.; Ungerstedt, J.S.; Olsson-Strömberg, U.; Nilsson, G. KIT signaling is dispensable for human mast cell progenitor development. Blood 2017, 130, 1785–1794. [Google Scholar] [CrossRef] [Green Version]
- Derakhshan, T.; Boyce, J.A.; Dwyer, D.F. Defining mast cell differentiation and heterogeneity through single-cell transcriptomics analysis. J. Allergy Clin. Immunol. 2022, 150, 739–747. [Google Scholar] [CrossRef]
- Bradding, P. Human lung mast cell heterogeneity. Thorax 2009, 64, 278–280. [Google Scholar] [CrossRef] [Green Version]
- St John, A.L.; Rathore, A.P.S.; Ginhoux, F. New perspectives on the origins and heterogeneity of mast cells. Nat. Rev. Immunol. 2023, 23, 55–68. [Google Scholar] [CrossRef]
- Irani, A.M.; Craig, S.S.; DeBlois, G.; Elson, C.O.; Schechter, N.M.; Schwartz, L.B. Deficiency of the Tryptase-Positive, Chymase-Negative Mast Cell Type in Gastrointestinal Mucosa of Patients with Defective T Lymphocyte Function. J. Immunol. 1987, 138, 4381–4386. [Google Scholar] [CrossRef]
- Trautmann, A.; Toksoy, A.; Engelhardt, E.; Bröcker, E.B.; Gillitzer, R. Mast cell involvement in normal human skin wound healing: Expression of monocyte chemoattractant protein-I is correlated with recruitment of mast cells which synthesize interleukin-4 in vivo. J. Pathol. 2000, 190, 100–106. [Google Scholar] [CrossRef]
- Lyons, D.O.; Pullen, N.A. Beyond IgE: Alternative mast cell activation across different disease states. Int. J. Mol. Sci. 2020, 21, 1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vadas, P.; Gold, M.; Perelman, B.; Liss, G.M.; Lack, G.; Blyth, T.; Simons, F.E.R.; Simons, K.J.; Cass, D.; Yeung, J. Platelet-Activating Factor, PAF Acetylhydrolase, and Severe Anaphylaxis. N. Engl. J. Med. 2008, 358, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Agier, J.; Pastwińska, J.; Brzezińska-Błaszczyk, E. An overview of mast cell pattern recognition receptors. Inflamm. Res. 2018, 67, 737–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theoharides, T.C.; Enakuaa, S.; Sismanopoulos, N.; Asadi, S.; Papadimas, E.C.; Angelidou, A.; Alysandratos, K.D. Contribution of stress to asthma worsening through mast cell activation. Ann. Allergy Asthma Immunol. 2012, 109, 14–19. [Google Scholar] [CrossRef]
- Redegeld, F.A.; Yu, Y.; Kumari, S.; Charles, N.; Blank, U. Non-IgE mediated mast cell activation. Immunol. Rev. 2018, 282, 87–113. [Google Scholar] [CrossRef]
- Boyden, S.E.; Desai, A.; Cruse, G.; Young, M.L.; Bolan, H.C.; Scott, L.M.; Eisch, A.R.; Long, R.D.; Lee, C.-C.R.; Satorius, C.L.; et al. Vibratory Urticaria Associated with a Missense Variant in ADGRE2. N. Engl. J. Med. 2016, 374, 656–663. [Google Scholar] [CrossRef]
- Kolkhir, P.; Altrichter, S.; Munoz, M.; Hawro, T.; Maurer, M. New treatments for chronic urticaria. Ann. Allergy Asthma Immunol. 2020, 124, 2–12. [Google Scholar] [CrossRef]
- Dispenza, M.C.; Krier-Burris, R.A.; Chhiba, K.D.; Undem, B.J.; Robida, P.A.; Bochner, B.S. Bruton’s tyrosine kinase inhibition effectively protects against human IgE-mediated anaphylaxis. J. Clin. Investig. 2020, 130, 4759–4770. [Google Scholar] [CrossRef]
- Harrison, C.A.; Bastan, R.; Peirce, M.J.; Munday, M.R.; Peachell, P.T. Role of calcineurin in the regulation of human lung mast cell and basophil function by cyclosporine and FK506. Br. J. Pharmacol. 2007, 150, 509–518. [Google Scholar] [CrossRef] [Green Version]
- West, P.W.; Bahri, R.; Garcia-Rodriguez, K.M.; Sweetland, G.; Wileman, G.; Shah, R.; Montero, A.; Rapley, L.; Bulfone-Paus, S. Interleukin-33 Amplifies Human Mast Cell Activities Induced by Complement Anaphylatoxins. Front. Immunol. 2021, 11, 615236. [Google Scholar] [CrossRef]
- Sumbayev, V.V.; Yasinska, I.; Oniku, A.E.; Streatfield, C.L.; Gibbs, B.F. Involvement of hypoxia-inducible factor-1 in the inflammatory responses of human LAD2 mast cells and basophils. PLoS ONE 2012, 7, e34259. [Google Scholar] [CrossRef] [Green Version]
- Fettrelet, T.; Gigon, L.; Karaulov, A.; Yousefi, S.; Simon, H.U. The enigma of eosinophil degranulation. Int. J. Mol. Sci. 2021, 22, 7091. [Google Scholar] [CrossRef]
- Begueret, H.; Berger, P.; Vernejoux, J.M.; Dubuisson, L.; Marthan, R.; Tunon-De-Lara, J.M. Inflammation of bronchial smooth muscle in allergic asthma. Thorax 2007, 62, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Sahid, M.N.A.; Kiyoi, T. Mast cell activation markers for in vitro study. J. Immunoass. Immunochem. 2020, 41, 778–816. [Google Scholar] [CrossRef]
- Ramu, S.; Akbarshahi, H.; Mogren, S.; Berlin, F.; Cerps, S.; Menzel, M.; Hvidtfeldt, M.; Porsbjerg, C.; Uller, L.; Andersson, C.K. Direct effects of mast cell proteases, tryptase and chymase, on bronchial epithelial integrity proteins and anti-viral responses. BMC Immunol. 2021, 22, 35. [Google Scholar] [CrossRef]
- Varricchi, G.; Rossi, F.W.; Galdiero, M.R.; Granata, F.; Criscuolo, G.; Spadaro, G.; De Paulis, A.; Marone, G. Physiological Roles of Mast Cells: Collegium Internationale Allergologicum Update 2019. Int. Arch. Allergy Immunol. 2019, 179, 247–261. [Google Scholar] [CrossRef]
- Watts, A.M.; Cripps, A.W.; West, N.P.; Cox, A.J. Modulation of allergic inflammation in the nasal mucosa of allergic rhinitis sufferers with topical pharmaceutical agents. Front. Pharmacol. 2019, 10, 294. [Google Scholar] [CrossRef] [Green Version]
- Bradding, P.; Walls, A.F.; Holgate, S.T. The role of the mast cell in the pathophysiology of asthma. J. Allergy Clin. Immunol. 2006, 117, 1277–1284. [Google Scholar] [CrossRef]
- Bradding, P.; Pejler, G. The controversial role of mast cells in fibrosis. Immunol. Rev. 2018, 282, 198–231. [Google Scholar] [CrossRef] [Green Version]
- Gangwar, R.S.; Pahima, H.; Puzzovio, P.G.; Levi-Schaffer, F. Update on Eosinophil Interaction with Mast Cells: The Allergic Effector Unit. Methods Mol. Biol. 2021, 2241, 221–242. [Google Scholar] [CrossRef]
- Lotfi-Emran, S.; Ward, B.R.; Le, Q.T.; Pozez, A.L.; Manjili, M.H.; Woodfolk, J.A.; Schwartz, L.B. Human mast cells present antigen to autologous CD4+ T cells. J. Allergy Clin. Immunol. 2018, 141, 311–321.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kambayashi, T.; Laufer, T.M. Atypical MHC class II-expressing antigen-presenting cells: Can anything replace a dendritic cell? Nat. Rev. Immunol. 2014, 14, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Mion, F.; D’Incà, F.; Danelli, L.; Toffoletto, B.; Guarnotta, C.; Frossi, B.; Burocchi, A.; Rigoni, A.; Gerdes, N.; Lutgens, E.; et al. Mast Cells Control the Expansion and Differentiation of IL-10–Competent B Cells. J. Immunol. 2014, 193, 4568–4579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Incà, F.; Pucillo, C.E. Exosomes: Tiny clues for mast cell communication. Front. Immunol. 2015, 6, 73. [Google Scholar] [CrossRef] [Green Version]
- Dudeck, A.; Suender, C.A.; Kostka, S.L.; von Stebut, E.; Maurer, M. Mast cells promote Th1 and Th17 responses by modulating dendritic cell maturation and function. Eur. J. Immunol. 2011, 41, 1883–1893. [Google Scholar] [CrossRef]
- Palm, A.K.E.; Garcia-Faroldi, G.; Lundberg, M.; Pejler, G.; Kleinau, S. Activated mast cells promote differentiation of B cells into effector cells. Sci. Rep. 2016, 6, 20531. [Google Scholar] [CrossRef] [Green Version]
- Piliponsky, A.M.; Romani, L. The contribution of mast cells to bacterial and fungal infection immunity. Immunol. Rev. 2018, 282, 188–197. [Google Scholar] [CrossRef]
- Meiners, J.; Reitz, M.; Rüdiger, N.; Turner, J.E.; Heepmann, L.; Rudolf, L.; Hartmann, W.; McSorley, H.J.; Breloer, M. IL-33 facilitates rapid expulsion of the parasitic nematode Strongyloides ratti from the intestine via ILC2- and IL-9-driven mast cell activation. PLoS Pathog. 2020, 16, e1009121. [Google Scholar] [CrossRef]
- Rathore, A.P.; St John, A.L. Protective and pathogenic roles for mast cells during viral infections. Curr. Opin. Immunol. 2020, 66, 74–81. [Google Scholar] [CrossRef]
- Warburg, O. Origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef]
- Hedeskov, C.J. Early effects of phytohaemagglutinin on glucose metabolism of normal human lymphocytes. Biochem. J. 1968, 110, 373–380. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Marquardt, C.; Foker, J. Aerobic glycolysis during lymphocyte proliferation. Nature 1976, 261, 702–705. [Google Scholar] [CrossRef]
- Chakravarty, N. Glycolysis in rat peritoneal mast cells. J. Cell Biol. 1965, 25, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Goretzki, A.; Lin, Y.J.; Schülke, S. Immune metabolism in allergies, does it matter?—A review of immune metabolic basics and adaptations associated with the activation of innate immune cells in allergy. Allergy Eur. J. Allergy Clin. Immunol. 2021, 76, 3314–3331. [Google Scholar] [CrossRef]
- Chakravarty, N. Further Observations on the Inhibition of Histamine Release by 2-Deoxyglucose. Acta Physiol. Scand. 1968, 72, 425–432. [Google Scholar] [CrossRef]
- Caslin, H.L.; Taruselli, M.T.; Haque, T.; Pondicherry, N.; Baldwin, E.A.; Barnstein, B.O.; Ryan, J.J. Inhibiting Glycolysis and ATP Production Attenuates IL-33-Mediated Mast Cell Function and Peritonitis. Front. Immunol. 2018, 9, 3026. [Google Scholar] [CrossRef]
- Ryu, H.; Walker, J.K.L.; Kim, S.; Koo, N.; Barak, L.S.; Noguchi, T.; Kang, B.Y.; Kim, K.M. Regulation of M 2-type pyruvate kinase mediated by the high-affinity IgE receptors is required for mast cell degranulation. Br. J. Pharmacol. 2008, 154, 1035–1046. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.J.; Goretzki, A.; Rainer, H.; Zimmermann, J.; Schülke, S. Immune Metabolism in TH2 Responses: New Opportunities to Improve Allergy Treatment—Cell Type-Specific Findings (Part 2). Curr. Allergy Asthma Rep. 2023, 23, 41–52. [Google Scholar] [CrossRef]
- Gelardi, M.; Giancaspro, R.; Cassano, M.; Ribatti, D. The Underestimated Role of Mast Cells in the Pathogenesis of Rhinopathies. Int. Arch. Allergy Immunol. 2022, 183, 153–159. [Google Scholar] [CrossRef]
- Banafea, G.H.; Bakhashab, S.; Alshaibi, H.F.; Natesan Pushparaj, P.; Rasool, M. The role of human mast cells in allergy and asthma. Bioengineered 2022, 13, 7049–7064. [Google Scholar] [CrossRef]
- Schuler IV, C.F.; Montejo, J.M. Allergic Rhinitis in Children and Adolescents. Pediatr. Clin. N. Am. 2019, 66, 981–993. [Google Scholar] [CrossRef] [PubMed]
- Casale, T.B.; Dykewicz, M.S. Clinical Implications of the Allergic Rhinitis-Asthma Link. Am. J. Med. Sci. 2004, 327, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Wise, S.K.; Lin, S.Y.; Toskala, E.; Orlandi, R.R.; Akdis, C.A.; Alt, J.A.; Azar, A.; Baroody, F.M.; Bachert, C.; Canonica, G.W.; et al. International Consensus Statement on Allergy and Rhinology: Allergic Rhinitis. Int. Forum Allergy Rhinol. 2018, 8, 108–352. [Google Scholar] [CrossRef] [PubMed]
- Fokkens, W.J.; Lund, V.J.; Hopkins, C.; Hellings, P.W.; Kern, S.R. International Rhinology Journal European Position Paper on Rhinosinusitis and Nasal Polyps. Epos 2020, 1, 7–8. [Google Scholar]
- Heffler, E.; Brussino, L.; Del Giacco, S.; Paoletti, G.; Minciullo, P.L.; Varricchi, G.; Scadding, G.; Malvezzi, L.; De Virgilio, A.; Spriano, G.; et al. New drugs in early-stage clinical trials for allergic rhinitis. Expert Opin. Investig. Drugs 2019, 28, 267–273. [Google Scholar] [CrossRef]
- Zoabi, Y.; Levi-Schaffer, F.; Eliashar, R. Allergic Rhinitis: Pathophysiology and Treatment Focusing on Mast Cells. Biomedicines 2022, 10, 2486. [Google Scholar] [CrossRef]
- Puzzovio, P.G.; Brüggemann, T.R.; Pahima, H.; Mankuta, D.; Levy, B.D.; Levi-Schaffer, F. Cromolyn Sodium differentially regulates human mast cell and mouse leukocyte responses to control allergic inflammation. Pharmacol. Res. 2022, 178, 106172. [Google Scholar] [CrossRef]
- Jo-Watanabe, A.; Okuno, T.; Yokomizo, T. The role of leukotrienes as potential therapeutic targets in allergic disorders. Int. J. Mol. Sci. 2019, 20, 3580. [Google Scholar] [CrossRef] [Green Version]
- Tsabouri, S. Omalizumab for the treatment of allergic rhinitis: A systematic review and meta-analysis*. Rhinology 2021, 59, 501–510. [Google Scholar] [CrossRef]
- Cavaliere, C.; Begvarfaj, E.; Incorvaia, C.; Sposato, B.; Brunori, M.; Ciofalo, A.; Greco, A.; de Vincentiis, M.; Masieri, S. Long-term omalizumab efficacy in allergic rhinitis. Immunol. Lett. 2020, 227, 81–87. [Google Scholar] [CrossRef]
- Gevaert, P.; Omachi, T.A.; Corren, J.; Mullol, J.; Han, J.; Lee, S.E.; Kaufman, D.; Ligueros-Saylan, M.; Howard, M.; Zhu, R.; et al. Efficacy and safety of omalizumab in nasal polyposis: 2 randomized phase 3 trials. J. Allergy Clin. Immunol. 2020, 146, 595–605. [Google Scholar] [CrossRef]
- Weinstein, S.F.; Katial, R.; Jayawardena, S.; Pirozzi, G.; Staudinger, H.; Eckert, L.; Joish, V.N.; Amin, N.; Maroni, J.; Rowe, P.; et al. Efficacy and safety of dupilumab in perennial allergic rhinitis and comorbid asthma. J. Allergy Clin. Immunol. 2018, 142, 171–177.e1. [Google Scholar] [CrossRef] [Green Version]
- Bachert, C.; Han, J.K.; Desrosiers, M.; Hellings, P.W.; Amin, N.; Lee, S.E.; Mullol, J.; Greos, L.S.; Bosso, J.V.; Laidlaw, T.M.; et al. Efficacy and safety of dupilumab in patients with severe chronic rhinosinusitis with nasal polyps (LIBERTY NP SINUS-24 and LIBERTY NP SINUS-52): Results from two multicentre, randomised, double-blind, placebo-controlled, parallel-group phase 3 trials. Lancet 2019, 394, 1638–1650. [Google Scholar] [CrossRef] [Green Version]
- Peters, A.T.; Han, J.K.; Hellings, P.; Heffler, E.; Gevaert, P.; Bachert, C.; Xu, Y.; Chuang, C.C.; Neupane, B.; Msihid, J.; et al. Indirect Treatment Comparison of Biologics in Chronic Rhinosinusitis with Nasal Polyps. J. Allergy Clin. Immunol. Pract. 2021, 9, 2461–2471.e5. [Google Scholar] [CrossRef]
- Akdis, M.; Akdis, C.A. Mechanisms of allergen-specific immunotherapy: Multiple suppressor factors at work in immune tolerance to allergens. J. Allergy Clin. Immunol. 2014, 133, 621–631. [Google Scholar] [CrossRef]
- Piliponsky, A.M.; Acharya, M.; Shubin, N.J. Mast cells in viral, bacterial, and fungal infection immunity. Int. J. Mol. Sci. 2019, 20, 2851. [Google Scholar] [CrossRef] [Green Version]
- Heffler, E.; Landi, M.; Caruso, C.; Fichera, S.; Gani, F.; Guida, G.; Liuzzo, M.T.; Pistorio, M.P.; Pizzimenti, S.; Riccio, A.M.; et al. Nasal cytology: Methodology with application to clinical practice and research. Clin. Exp. Allergy 2018, 48, 1092–1106. [Google Scholar] [CrossRef]
- Dwyer, D.F.; Ordovas-Montanes, J.; Allon, S.J.; Buchheit, K.M.; Vukovic, M.; Derakhshan, T.; Feng, C.; Lai, J.; Hughes, T.K.; Nyquist, S.K.; et al. Human airway mast cells proliferate and acquire distinct inflammation-driven phenotypes during type 2 inflammation. Sci. Immunol. 2021, 6, eabb7221. [Google Scholar] [CrossRef]
- Takabayashi, T.; Schleimer, R. Formation of nasal polyps: The roles of innate type 2 inflammation and deposition of fibrin. J. Allergy Clin. Immunol. 2020, 145, 751. [Google Scholar] [CrossRef] [Green Version]
- GINA Committee. Global Strategy for Asthma Management and Prevention 2022 Update; Global Initiative for Asthma: Fontana, WI, USA, 2022; p. 225. [Google Scholar]
- Cockcroft, D.W.; Davis, B.E. Mechanisms of airway hyperresponsiveness. J. Allergy Clin. Immunol. 2006, 118, 551–559. [Google Scholar] [CrossRef]
- Al-Muhsen, S.; Johnson, J.; Hamid, Q. Remodeling in asthma. J. Allergy Clin. Immunol. 2011, 128, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Brightling, C.E.; Symon, F.A.; Birring, S.S.; Bradding, P.; Wardlaw, A.J.; Pavord, I.D. Comparison of airway immunopathology of eosinophilic bronchitis and asthma. Thorax 2003, 58, 528–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradding, P.; Brightling, C. Mast cell infiltration of airway smooth muscle in asthma. Respir. Med. 2007, 101, 1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brightling, C.E.; Kaur, D.; Berger, P.; Morgan, A.J.; Wardlaw, A.J.; Bradding, P. Differential expression of CCR3 and CXCR3 by human lung and bone marrow-derived mast cells: Implications for tissue mast cell migration. J. Leukoc. Biol. 2005, 77, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Brightling, C.E.; Ammit, A.J.; Kaur, D.; Black, J.L.; Wardlaw, A.J.; Hughes, J.M.; Bradding, P. The CXCL10/CXCR3 axis mediates human lung mast cell migration to asthmatic airway smooth muscle. Am. J. Respir. Crit. Care Med. 2005, 171, 1103–1108. [Google Scholar] [CrossRef]
- Hamanou, H.; Kawa, S.; Ochi, Y.; Unno, H.; Shiba, N.; Wajiki, M.; Nakazawa, K.; Shimojo, H.; Kiyosawa, K. Hydronephrosis associated with retroperitoneal fibrosis and sclerosing pancreatitis. Lancet 2002, 359, 1403–1404. [Google Scholar] [CrossRef]
- Berlin, F.; Mogren, S.; Tutzauer, J.; Andersson, C.K. Mast cell proteases tryptase and chymase induce migratory and morphological alterations in bronchial epithelial cells. Int. J. Mol. Sci. 2021, 22, 5250. [Google Scholar] [CrossRef]
- Metcalfe, D.D.; Pawankar, R.; Ackerman, S.J.; Akin, C.; Clayton, F.; Falcone, F.H.; Gleich, G.J.; Irani, A.M.; Johansson, M.W.; Klion, A.D.; et al. Biomarkers of the involvement of mast cells, basophils and eosinophils in asthma and allergic diseases. World Allergy Organ. J. 2016, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, S.E.; Schwartz, L.B. Activation of pulmonary mast cells by bronchoalveolar allergen challenge: In vivo release of histamine and tryptase in atopic subjects with and without asthma. Am. Rev. Respir. Dis. 1988, 137, 1002–1008. [Google Scholar] [CrossRef]
- Howarth, P.H.; Durham, S.R.; Lee, T.H.; Kay, A.B.; Church, M.K.; Holgate, S.T. Influence of albuterol, cromolyn sodium and ipratropium bromide on the airway and circulating mediator reponses to allergen bronchial provocation in asthma. Am. Rev. Respir. Dis. 1985, 132, 986–992. [Google Scholar]
- Kapsali, T.; Permutt, S.; Laube, B.; Scichilone, N.; Togias, A. Potent bronchoprotective effect of deep inspiration and its absence in asthma. J. Appl. Physiol. 2000, 89, 711–720. [Google Scholar] [CrossRef] [Green Version]
- Carroll, N.G.; Mutavdzic, S.; James, A.L. Distribution and degranulation of airway mast cells in normal and asthmatic subjects. Eur. Respir. J. 2002, 19, 879–885. [Google Scholar] [CrossRef] [Green Version]
- Carroll, N.G.; Mutavdzic, S.; James, A.L. Increased mast cells and neutrophils in submucosal mucous glands and mucus plugging in patients with asthma. Thorax 2002, 57, 677–682. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.W.; Oh, C.K.; Cho, S.H.; Hu, G.; Martin, R.; Demissie-Sanders, S.; Li, K.; Moyle, M.; Yao, Z. Amphiregulin expression in human mast cells and its effect on the primary human lung fibroblasts. J. Allergy Clin. Immunol. 2005, 115, 287–294. [Google Scholar] [CrossRef]
- Kuyper, L.M.; Paré, P.D.; Hogg, J.C.; Lambert, R.K.; Ionescu, D.; Woods, R.; Bai, T.R. Characterization of airway plugging in fatal asthma. Am. J. Med. 2003, 115, 6–11. [Google Scholar] [CrossRef]
- Al-Muhsen, S.Z.; Shablovsky, G.; Olivenstein, R.; Mazer, B.; Hamid, Q. The expression of stem cell factor and c-kit receptor in human asthmatic airways. Clin. Exp. Allergy 2004, 34, 911–916. [Google Scholar] [CrossRef]
- Saha, S.K.; Berry, M.A.; Parker, D.; Siddiqui, S.; Morgan, A.; May, R.; Monk, P.; Bradding, P.; Wardlaw, A.J.; Pavord, I.D.; et al. Increased sputum and bronchial biopsy IL-13 expression in severe asthma. J. Allergy Clin. Immunol. 2008, 121, 685–691. [Google Scholar] [CrossRef] [Green Version]
- Woodman, L.; Siddiqui, S.; Cruse, G.; Sutcliffe, A.; Saunders, R.; Kaur, D.; Bradding, P.; Brightling, C. Mast Cells Promote Airway Smooth Muscle Cell Differentiation via Autocrine Up-Regulation of TGF-β1. J. Immunol. 2008, 181, 5001–5007. [Google Scholar] [CrossRef] [Green Version]
- Kaur, D.; Saunders, R.; Hollins, F.; Woodman, L.; Doe, C.; Siddiqui, S.; Bradding, P.; Brightling, C. Mast Cell Fibroblastoid Differentiation Mediated by Airway Smooth Muscle in Asthma. J. Immunol. 2010, 185, 6105–6114. [Google Scholar] [CrossRef] [Green Version]
- Bradding, P.; Roberts, J.A.; Britten, K.M.; Montefort, S.; Djukanovic, R.; Mueller, R.; Heusser, C.H.; Howarth, P.H.; Holgate, S.T. Interleukin-4, -5, and -6 and tumor necrosis factor-alpha in normal and asthmatic airways: Evidence for the human mast cell as a source of these cytokines. Am. J. Respir. Cell Mol. Biol. 1994, 10, 471–480. [Google Scholar] [CrossRef]
- Berry, M.A.; Hargadon, B.; Shelley, M.; Parker, D.; Shaw, D.E.; Green, R.H.; Bradding, P.; Brightling, C.E.; Wardlaw, A.J.; Pavord, I.D. Evidence of a Role of Tumor Necrosis Factor α in Refractory Asthma. N. Engl. J. Med. 2006, 354, 697–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roche, W.R.; Williams, J.H.; Beasley, R.; Holgate, S.T. Subepithelial Fibrosis in the Bronchi of Asthmatics. Lancet 1989, 333, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Su, G.; Zhong, Y.; Xiong, Z.; Huang, T.; Quan, J.; Huang, J.; Wen, X.; Luo, C.; Zheng, W.; et al. HB-EGF-induced IL-8 secretion from airway epithelium leads to lung fibroblast proliferation and migration. BMC Pulm. Med. 2021, 21, 347. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Pei, Q.M.; Jiang, P.; Liu, J.; Sun, R.F.; Qian, X.J.; Liu, J.B. Upregulation of MUC5AC by VEGF in human primary bronchial epithelial cells: Implications for asthma. Respir. Res. 2019, 20, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigoraş, A.; Cǎruntu, I.D.; Grigoraş, C.C.; Mihǎescu, T.; Amǎlinei, C. Relationship between immunohistochemical assessment of bronchial mucosa microvascularization and clinical stage in asthma. Rom. J. Morphol. Embryol. 2012, 53, 485–490. [Google Scholar]
- Lazarinis, N.; Bood, J.; Gomez, C.; Kolmert, J.; Lantz, A.-S.; Gyllfors, P.; Davis, A.; Wheelock, C.E.; Dahlén, S.-E.; Dahlén, B. Leukotriene E4 induces airflow obstruction and mast cell activation through the cysteinyl leukotriene type 1 receptor. J. Allergy Clin. Immunol. 2018, 142, 1080–1089. [Google Scholar] [CrossRef] [Green Version]
- Löfdahl, A.; Wenglén, C.; Rydell-Törmänen, K.; Westergren-Thorsson, G.; Larsson-Callerfelt, A.K. Effects of 5-Hydroxytryptamine Class 2 Receptor Antagonists on Bronchoconstriction and Pulmonary Remodeling Processes. Am. J. Pathol. 2018, 188, 1113–1119. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, S.; Sutcliffe, A.; Shikotra, A.; Woodman, L.; Doe, C.; McKenna, S.; Wardlaw, A.; Bradding, P.; Pavord, I.; Brightling, C. Vascular remodeling is a feature of asthma and nonasthmatic eosinophilic bronchitis. J. Allergy Clin. Immunol. 2007, 120, 813–819. [Google Scholar] [CrossRef]
- Birring, S.S.; Parker, D.; Brightling, C.E.; Bradding, P.; Wardlaw, A.J.; Pavord, I.D. Induced Sputum Inflammatory Mediator Concentrations in Chronic Cough. Am. J. Respir. Crit. Care Med. 2004, 169, 15–19. [Google Scholar] [CrossRef]
- Djukanović, R.; Wilson, S.J.; Kraft, M.; Jarjour, N.N.; Steel, M.; Chung, K.F.; Bao, W.; Fowler-Taylor, A.; Matthews, J.; Busse, W.W.; et al. Effects of treatment with anti-immunoglobulin E antibody omalizumab on airway inflammation in allergic asthma. Am. J. Respir. Crit. Care Med. 2004, 170, 583–893. [Google Scholar] [CrossRef]
- Kulka, M.; Alexopoulou, L.; Flavell, R.A.; Metcalfe, D.D. Activation of mast cells by double-stranded RNA: Evidence for activation through Toll-like receptor 3. J. Allergy Clin. Immunol. 2004, 114, 174–182. [Google Scholar] [CrossRef] [Green Version]
- Holgate, S.T. The Quintiles Prize Lecture 2004. The identification of the adenosine A2B receptor as a novel therapeutic target in asthma. Br. J. Pharmacol. 2005, 145, 1009–1015. [Google Scholar] [CrossRef] [Green Version]
- Gross, A.R.; Theoharides, T.C. Chondroitin sulfate inhibits secretion of TNF and CXCL8 from human mast cells stimulated by IL-33. BioFactors 2019, 45, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, S.; Willmes, D.M.; Nassiri, M.; Babina, M.; Worm, M. PGE2 deficiency predisposes to anaphylaxis by causing mast cell hyperresponsiveness. J. Allergy Clin. Immunol. 2020, 146, 1387–1396.e13. [Google Scholar] [CrossRef]
- Maric, J.; Ravindran, A.; Mazzurana, L.; Björklund, Å.K.; Van Acker, A.; Rao, A.; Friberg, D.; Dahlén, S.E.; Heinemann, A.; Konya, V.; et al. Prostaglandin E2 suppresses human group 2 innate lymphoid cell function. J. Allergy Clin. Immunol. 2018, 141, 1761–1773.e6. [Google Scholar] [CrossRef] [Green Version]
- Maun, H.R.; Jackman, J.K.; Choy, D.F.; Loyet, K.M.; Staton, T.L.; Jia, G.; Dressen, A.; Hackney, J.A.; Bremer, M.; Walters, B.T.; et al. An Allosteric Anti-tryptase Antibody for the Treatment of Mast Cell-Mediated Severe Asthma. Cell 2019, 179, 417–431.e19. [Google Scholar] [CrossRef] [Green Version]
- Mogren, S.; Berlin, F.; Ramu, S.; Sverrild, A.; Porsbjerg, C.; Uller, L.; Andersson, C.K. Mast cell tryptase enhances wound healing by promoting migration in human bronchial epithelial cells. Cell Adhes. Migr. 2021, 15, 202–214. [Google Scholar] [CrossRef]
- Miyagawa, Y.; Murakami, A.; Ebihara, N. The proteolytic effect of mast cell tryptase to eotaxin-1/CCL11·eotaxin-2/CCL24 and eotaxin-3/CCL26 produced by conjunctival fibroblasts. Jpn. J. Ophthalmol. 2019, 63, 215–220. [Google Scholar] [CrossRef]
- Cristinziano, L.; Poto, R.; Criscuolo, G.; Ferrara, A.L.; Galdiero, M.R.; Modestino, L.; Loffredo, S.; de Paulis, A.; Marone, G.; Spadaro, G.; et al. Il-33 and superantigenic activation of human lung mast cells induce the release of angiogenic and lymphangiogenic factors. Cells 2021, 10, 145. [Google Scholar] [CrossRef]
- Ebina, M. Remodeling of airway walls in fatal asthmatics decreases lymphatic distribution; beyond thickening of airway smooth muscle layers. Allergol. Int. 2008, 57, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Bentley, A.M.; Menz, G.; Storz, C.; Robinson, D.S.; Bradley, B.; Jeffery, P.K.; Durham, S.R.; Kay, A.B. Identification of T lymphocytes, macrophages, and activated eosinophils in the bronchial mucosa in intrinsic asthma: Relationship to symptoms and bronchial responsiveness. Am. Rev. Respir. Dis. 1992, 146, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Humbert, M.; Grant, J.A.; Taborda-Barata, L.; Durham, S.R.; Pfister, R.; Menz, G.; Barkans, J.; Ying, S.; Kay, A.B. High-affinity IgE receptor (FcεRI)-bearing cells in bronchial biopsies from atopic and nonatopic asthma. Am. J. Respir. Crit. Care Med. 1996, 153, 1931–1937. [Google Scholar] [CrossRef] [PubMed]
- Saetta, M.; Di Stefano, A.; Maestrelli, P.; De Marzo, N.; Milani, G.F.; Pivirotto, F.; Mapp, C.E.; Fabbri, L.M. Airway mucosal inflammation in occupational asthma induced by toluene diisocyanate. Am. Rev. Respir. Dis. 1992, 145, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.D.; Bye, P.T.P.; Schoeffel, R.E.; Seale, J.P.; Taylor, K.M.; Ferris, L. Arterial plasma histamine levels at rest, and during and after exercise in patients with asthma: Effects of terbutaline aerosol. Thorax 1981, 36, 259–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brannan, J.D.; Turton, J.A. The inflammatory basis of exercise-induced bronchoconstriction. Phys. Sportsmed. 2010, 38, 67–73. [Google Scholar] [CrossRef]
- Paivandy, A.; Pejler, G. Novel Strategies to Target Mast Cells in Disease. J. Innate Immun. 2021, 13, 131–147. [Google Scholar] [CrossRef]
- Levy, M.L.; Bacharier, L.B.; Bateman, E.; Boulet, L.P.; Brightling, C.; Buhl, R.; Brusselle, G.; Cruz, A.A.; Drazen, J.M.; Duijts, L.; et al. Key recommendations for primary care from the 2022 Global Initiative for Asthma (GINA) update. npj Prim. Care Respir. Med. 2023, 33, 7. [Google Scholar] [CrossRef]
- Peachell, P. Regulation of mast cells by β-agonists. Clin. Rev. Allergy Immunol. 2006, 31, 131–142. [Google Scholar] [CrossRef]
- Luo, Y.; Jin, M.; Lou, L.; Yang, S.; Li, C.; Li, X.; Zhou, M.; Cai, C. Role of arachidonic acid lipoxygenase pathway in Asthma. Prostaglandins Other Lipid Mediat. 2022, 158, 106609. [Google Scholar] [CrossRef]
- Mellor, E.A.; Frank Austen, K.; Boyce, J.A. Cysteinyl leukotrienes and uridine diphosphate induce cytokine generation by human mast cells through an interleukin 4-regulated pathway that is inhibited by leukotriene receptor antagonists. J. Exp. Med. 2002, 195, 583–592. [Google Scholar] [CrossRef] [Green Version]
- Nappi, E.; Paoletti, G.; Malvezzi, L.; Ferri, S.; Racca, F.; Messina, M.R.; Puggioni, F.; Heffler, E.; Canonica, G.W. Comorbid allergic rhinitis and asthma: Important clinical considerations. Expert Rev. Clin. Immunol. 2022, 18, 747–758. [Google Scholar] [CrossRef]
- Ducharme, F.; di Salvio, F. Anti-leukotriene agents compared to inhaled corticosteroids in the management of recurrent and/or chronic asthma in adults and children. Cochrane Database Syst. Rev. 2004, 1, CD002314. [Google Scholar] [CrossRef] [Green Version]
- Gasser, P.; Tarchevskaya, S.S.; Guntern, P.; Brigger, D.; Ruppli, R.; Zbären, N.; Kleinboelting, S.; Heusser, C.; Jardetzky, T.S.; Eggel, A. The mechanistic and functional profile of the therapeutic anti-IgE antibody ligelizumab differs from omalizumab. Nat. Commun. 2020, 11, 165. [Google Scholar] [CrossRef] [Green Version]
- Maurer, M.; Giménez-Arnau, A.M.; Sussman, G.; Metz, M.; Baker, D.R.; Bauer, A.; Bernstein, J.A.; Brehler, R.; Chu, C.-Y.; Chung, W.-H.; et al. Ligelizumab for Chronic Spontaneous Urticaria. N. Engl. J. Med. 2019, 381, 1321–1332. [Google Scholar] [CrossRef] [Green Version]
- Trischler, J.; Bottoli, I.; Janocha, R.; Heusser, C.; Jaumont, X.; Lowe, P.; Gautier, A.; Pethe, A.; Woessner, R.; Zerwes, H.G.; et al. Ligelizumab treatment for severe asthma: Learnings from the clinical development programme. Clin. Transl. Immunol. 2021, 10, e1255. [Google Scholar] [CrossRef]
- Buchheit, K.M.; Lewis, E.; Gakpo, D.; Hacker, J.; Sohail, A.; Taliaferro, F.; Berreondo Giron, E.; Asare, C.; Vukovic, M.; Bensko, J.C.; et al. Mepolizumab targets multiple immune cells in aspirin-exacerbated respiratory disease. J. Allergy Clin. Immunol. 2021, 148, 574–584. [Google Scholar] [CrossRef]
- Burchett, J.R.; Dailey, J.M.; Kee, S.A.; Pryor, D.T.; Kotha, A.; Kankaria, R.A.; Straus, D.B.; Ryan, J.J. Targeting Mast Cells in Allergic Disease: Current Therapies and Drug Repurposing. Cells 2022, 11, 3031. [Google Scholar] [CrossRef]
- Menzies-Gow, A.; Corren, J.; Bourdin, A.; Chupp, G.; Israel, E.; Wechsler, M.E.; Brightling, C.E.; Griffiths, J.M.; Hellqvist, Å.; Bowen, K.; et al. Tezepelumab in Adults and Adolescents with Severe, Uncontrolled Asthma. N. Engl. J. Med. 2021, 384, 1800–1809. [Google Scholar] [CrossRef]
- Ebina-Shibuya, R.; Leonard, W.J. Role of thymic stromal lymphopoietin in allergy and beyond. Nat. Rev. Immunol. 2023, 23, 24–37. [Google Scholar] [CrossRef]
- Diver, S.; Khalfaoui, L.; Emson, C.; Wenzel, S.E.; Menzies-Gow, A.; Wechsler, M.E.; Johnston, J.; Molfino, N.; Parnes, J.R.; Megally, A.; et al. Effect of tezepelumab on airway inflammatory cells, remodelling, and hyperresponsiveness in patients with moderate-to-severe uncontrolled asthma (CASCADE): A double-blind, randomised, placebo-controlled, phase 2 trial. Lancet Respir. Med. 2021, 9, 1299–1312. [Google Scholar] [CrossRef]
- Wechsler, M.E.; Ruddy, M.K.; Pavord, I.D.; Israel, E.; Rabe, K.F.; Ford, L.B.; Maspero, J.F.; Abdulai, R.M.; Hu, C.-C.; Martincova, R.; et al. Efficacy and Safety of Itepekimab in Patients with Moderate-to-Severe Asthma. N. Engl. J. Med. 2021, 385, 1656–1668. [Google Scholar] [CrossRef] [PubMed]
- Porsbjerg, C.M.; Sverrild, A.; Lloyd, C.M.; Menzies-Gow, A.N.; Bel, E.H. Anti-alarmins in asthma: Targeting the airway epithelium with next-generation biologics. Eur. Respir. J. 2020, 56, 2000260. [Google Scholar] [CrossRef] [PubMed]
- Kerr, S.C.; Gonzalez, J.R.; Schanin, J.; Peters, M.C.; Lambrecht, B.N.; Brock, E.C.; Charbit, A.; Ansel, K.M.; Youngblood, B.A.; Fahy, J.V. An anti-siglec-8 antibody depletes sputum eosinophils from asthmatic subjects and inhibits lung mast cells. Clin. Exp. Allergy 2020, 50, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Poto, R.; Criscuolo, G.; Marone, G.; Brightling, C.E.; Varricchi, G. Human Lung Mast Cells: Therapeutic Implications in Asthma. Int. J. Mol. Sci. 2022, 23, 14466. [Google Scholar] [CrossRef]
- Cahill, K.N.; Katz, H.R.; Cui, J.; Lai, J.; Kazani, S.; Crosby-Thompson, A.; Garofalo, D.; Castro, M.; Jarjour, N.; DiMango, E.; et al. KIT Inhibition by Imatinib in Patients with Severe Refractory Asthma. N. Engl. J. Med. 2017, 376, 1911–1920. [Google Scholar] [CrossRef] [Green Version]
- Georas, S.N.; Donohue, P.; Connolly, M.; Wechsler, M.E. JAK inhibitors for asthma. J. Allergy Clin. Immunol. 2021, 148, 953–963. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costanzo, G.; Costanzo, G.A.M.L.; Del Moro, L.; Nappi, E.; Pelaia, C.; Puggioni, F.; Canonica, G.W.; Heffler, E.; Paoletti, G. Mast Cells in Upper and Lower Airway Diseases: Sentinels in the Front Line. Int. J. Mol. Sci. 2023, 24, 9771. https://doi.org/10.3390/ijms24119771
Costanzo G, Costanzo GAML, Del Moro L, Nappi E, Pelaia C, Puggioni F, Canonica GW, Heffler E, Paoletti G. Mast Cells in Upper and Lower Airway Diseases: Sentinels in the Front Line. International Journal of Molecular Sciences. 2023; 24(11):9771. https://doi.org/10.3390/ijms24119771
Chicago/Turabian StyleCostanzo, Giovanni, Giulia Anna Maria Luigia Costanzo, Lorenzo Del Moro, Emanuele Nappi, Corrado Pelaia, Francesca Puggioni, Giorgio Walter Canonica, Enrico Heffler, and Giovanni Paoletti. 2023. "Mast Cells in Upper and Lower Airway Diseases: Sentinels in the Front Line" International Journal of Molecular Sciences 24, no. 11: 9771. https://doi.org/10.3390/ijms24119771