

Comparative Analysis of Fcγ and Complement Receptors Presence on Monocytes in Pulmonary Sarcoidosis and Tuberculosis

Abstract
:1. Introduction
2. Results
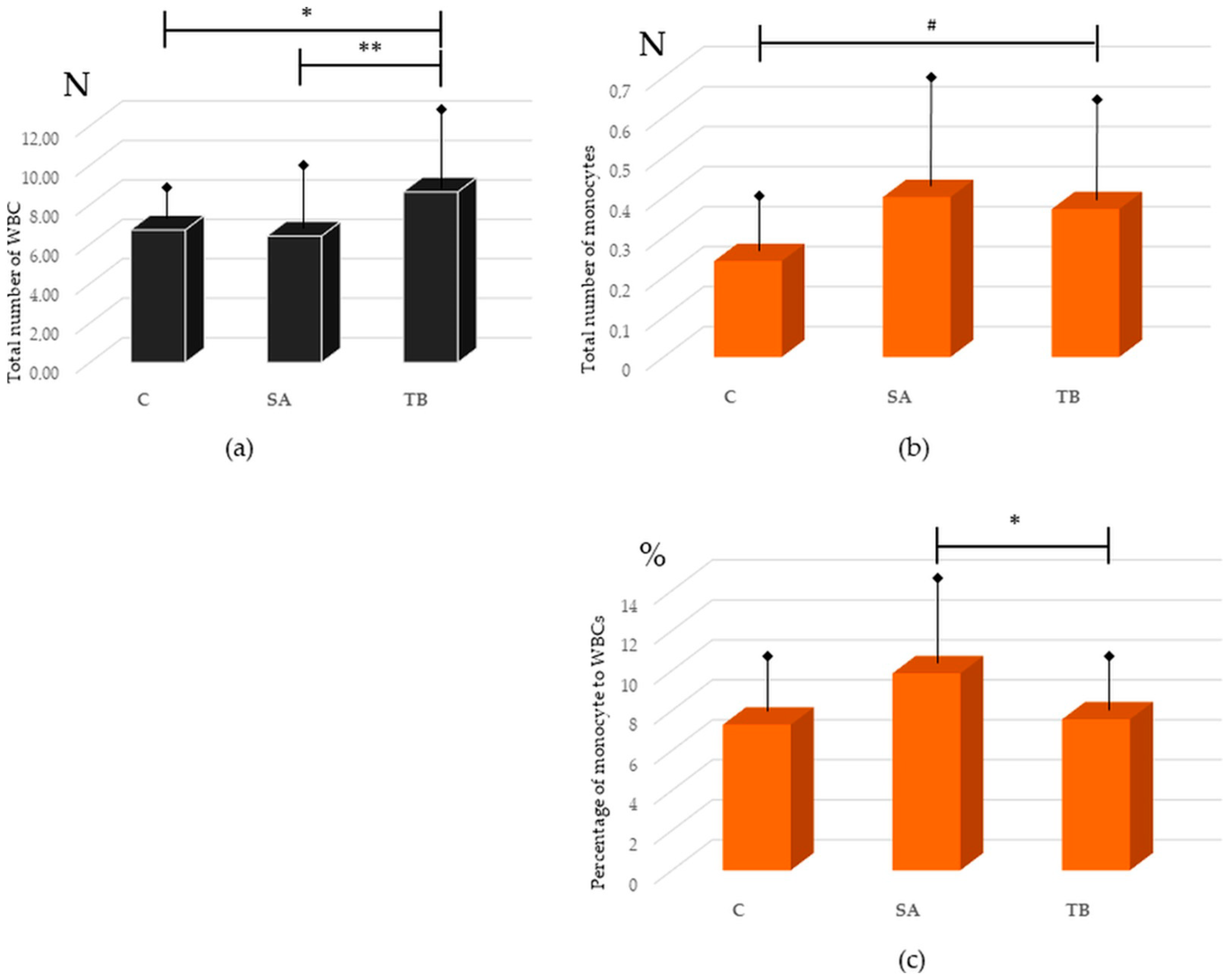
2.1. Presence of Monocytes in Peripheral Blood from SA Patients, TB Patients, and Healthy Control
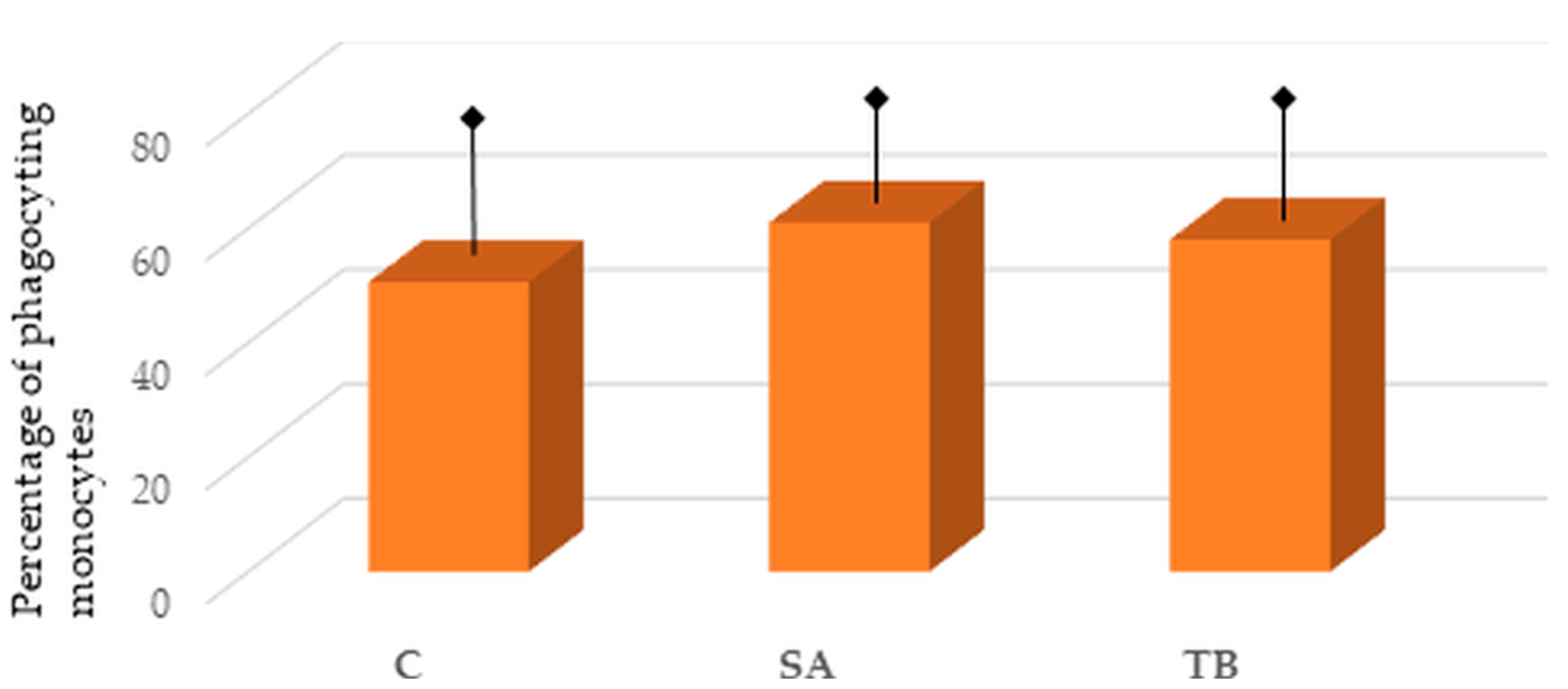
2.2. Percentage of Phagocyting Blood Monocytes in the Tested Groups
2.3. Comparative Analysis of the Occurrence of FcγRI-III and CR1, 3, and 4 on Peripheral Blood Monocytes in All Tested Groups
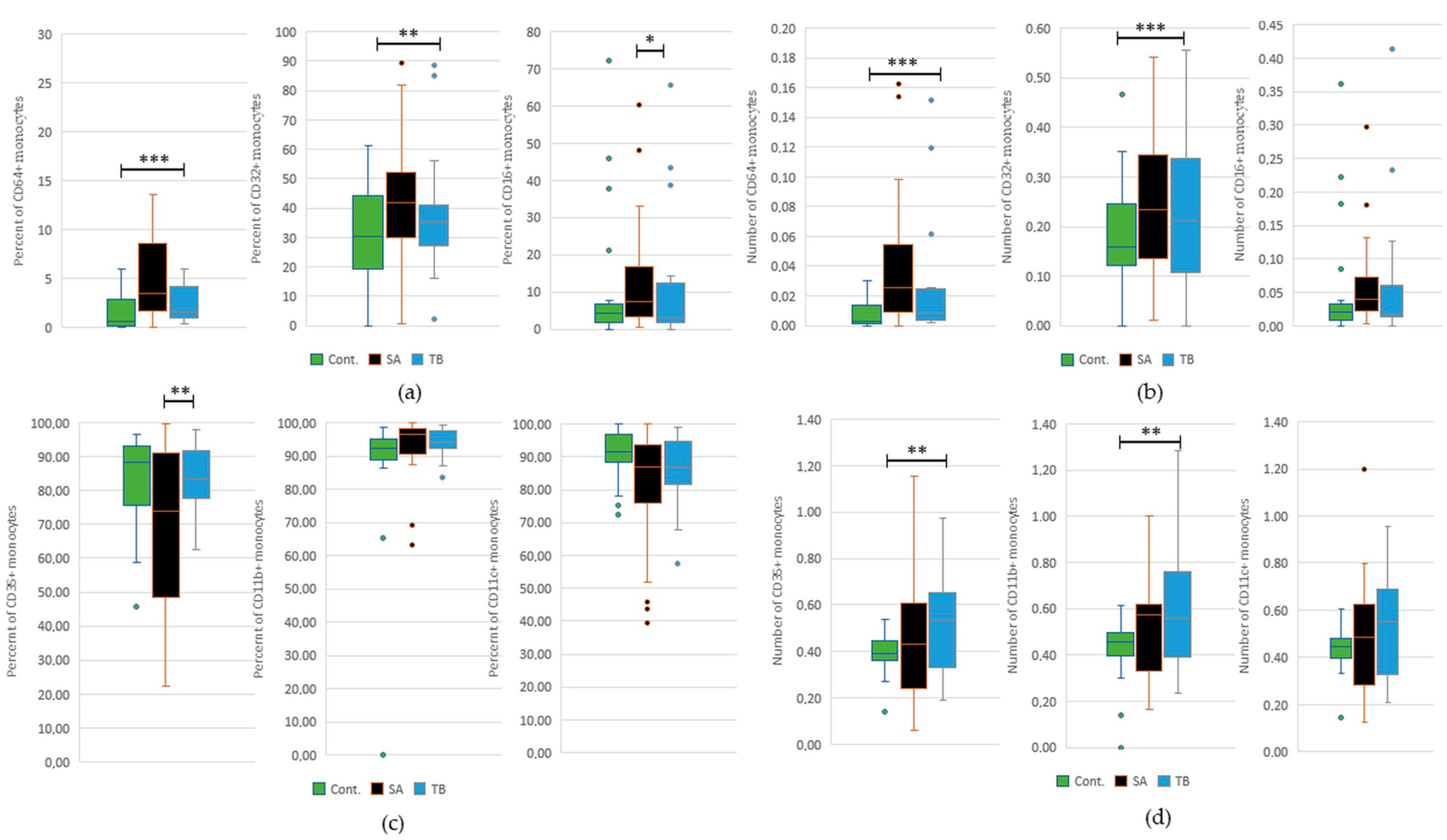
2.4. Comparative Analysis of the Percentage and Total Number of Monocytes with FcγRI-III and/or CR1, 3, and 4 in Peripheral Blood of SA and TB Patients
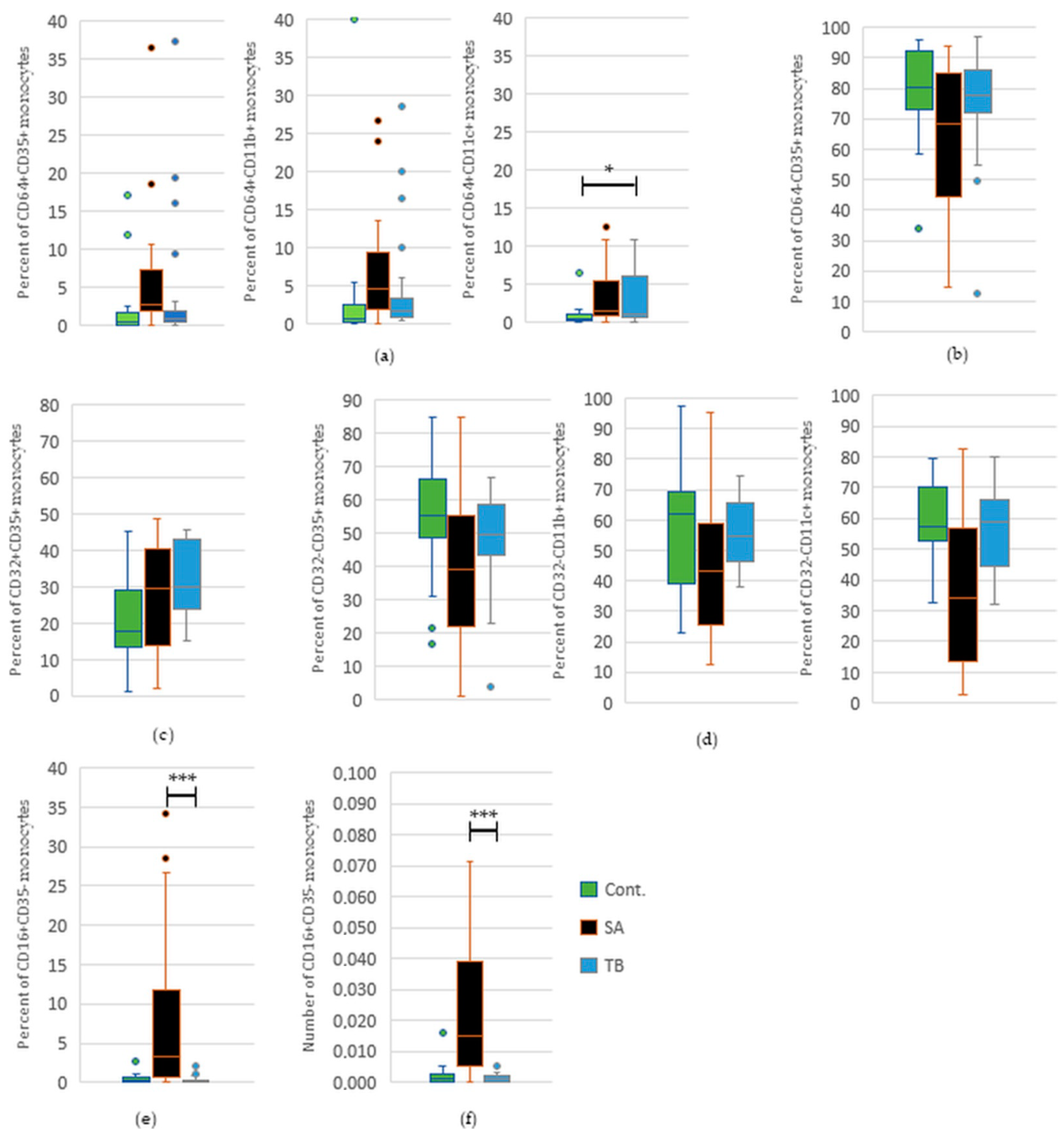
2.5. Comparative Analysis of the Percentage and Total Number of Monocytes with FcγRI-III and/or CR1, 3, and 4 in Peripheral Blood of TB Patients and Healthy Controls
3. Discussion
4. Materials and Methods
4.1. Study Participants
4.1.1. Patients with Sarcoidosis
4.1.2. Patients with Tuberculosis
4.1.3. Healthy Controls
4.2. Methods
4.2.1. Sample Collection and Preparation
4.2.2. Used Reagents and Antibodies
4.2.3. PBMC Isolation
4.2.4. Flow Cytometry
Analysis of Fcγ and Complement Receptor Presence at the Surface of CD14+ Monocytes
Analysis of the Phagocytic Activity of Monocytes
4.2.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mortaz, E.; Masjedi, M.R.; Abedini, A.; Matroodi, S.; Kiani, A.; Soroush, D.; Adcock, I.M. Common features of tuberculosis and sarcoidosis. Int. J. Mycobacteriol. 2016, 5 (Suppl. 1), S240–S241. [Google Scholar] [CrossRef]
- Dubaniewicz, A.; Zimmermann, A.; Dudziak, M.; Typiak, M.; Skotarczak, M. Tuberculosis in the course of sarcoidosis treatment: Is genotyping necessary for personalized therapy? Expert Rev. Clin. Immunol. 2013, 9, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Statement on sarcoidosis. Joint statement of the american thoracic society (ats), the european respiratory society (ers) and the world association of sarcoidosis and other granulomatous disorders (wasog) adopted by the ats board of directors and by the ers executive committee, February. Am. J. Respir. Crit. Care Med. 1999, 160, 736–755. [Google Scholar] [CrossRef]
- Chen, E.S.; Moller, D.R. Etiology of sarcoidosis. Clin. Chest. Med. 2008, 29, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Dubaniewicz, A. Mycobacterial heat shock proteins in sarcoidosis and tuberculosis. Int. J. Mol. Sci. 2023, 24, 5084. [Google Scholar] [CrossRef] [PubMed]
- Dubaniewicz, A. Mycobacterium tuberculosis heat shock proteins and autoimmunity in sarcoidosis. Autoimmun. Rev. 2010, 9, 419–424. [Google Scholar] [CrossRef]
- Dubaniewicz, A.; Kämpfer, S.; Singh, M. Serum anti-mycobacterial heat shock proteins antibodies in sarcoidosis and tuberculosis. Tuberculosis 2006, 86, 60–67. [Google Scholar] [CrossRef]
- Dubaniewicz, A.; Holownia, A.; Kalinowski, L.; Wybieralska, M.; Dobrucki, I.T.; Singh, M. Is mycobacterial heat shock protein 16 kda, a marker of the dormant stage of mycobacterium tuberculosis, a sarcoid antigen? Hum. Immunol. 2013, 74, 45–51. [Google Scholar] [CrossRef]
- Dubaniewicz, A.; Dubaniewicz-Wybieralska, M.; Sternau, A.; Zwolska, Z.; Izycka-Swieszewska, E.; Augustynowicz-Kopec, E.; Skokowski, J.; Singh, M.; Zimnoch, L. Mycobacterium tuberculosis complex and mycobacterial heat shock proteins in lymph node tissue from patients with pulmonary sarcoidosis. J. Clin. Microbiol. 2006, 44, 3448–3451. [Google Scholar] [CrossRef] [Green Version]
- Dubaniewicz, A.; Trzonkowski, P.; Dubaniewicz-Wybieralska, M.; Dubaniewicz, A.; Singh, M.; Myśliwski, A. Comparative analysis of mycobacterial heat shock proteins-induced apoptosis of peripheral blood mononuclear cells in sarcoidosis and tuberculosis. J. Clin. Immunol. 2006, 26, 243–250. [Google Scholar] [CrossRef]
- Dubaniewicz, A.; Trzonkowski, P.; Dubaniewicz-Wybieralska, M.; Dubaniewicz, A.; Singh, M.; Myśliwski, A. Mycobacterial heat shock protein-induced blood t lymphocytes subsets and cytokine pattern: Comparison of sarcoidosis with tuberculosis and healthy controls. Respirology 2007, 12, 346–354. [Google Scholar] [CrossRef]
- Dubaniewicz, A.; Rękawiecki, B.; Piprek, M.; Skotarczak, M.; Dubaniewicz-Wybieralska, M. Monocyte/neutrophil phagocytic activity ratio in differentiating tuberculosis from sarcoidosis. Eur. Respir. J. 2019, 54, PA1378. Available online: http://erj.ersjournals.com/content/54/suppl_63/PA1378.abstract (accessed on 14 April 2023). [CrossRef]
- Dubaniewicz, A.; Typiak, M.; Wybieralska, M.; Szadurska, M.; Nowakowski, S.; Staniewicz-Panasik, A.; Rogoza, K.; Sternau, A.; Deeg, P.; Trzonkowski, P. Changed phagocytic activity and pattern of fcγ and complement receptors on blood monocytes in sarcoidosis. Hum. Immunol. 2012, 73, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Lepzien, R.; Liu, S.; Czarnewski, P.; Nie, M.; Österberg, B.; Baharom, F.; Pourazar, J.; Rankin, G.; Eklund, A.; Bottai, M.; et al. Monocytes in sarcoidosis are potent tumour necrosis factor producers and predict disease outcome. Eur. Respir. J. 2021, 58, 2003468. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Daha, M.R.; Kallenberg, C.G. The complement system in systemic autoimmune disease. J. Autoimmun. 2010, 34, J276–J286. [Google Scholar] [CrossRef]
- Nimmerjahn, F.; Ravetch, J.V. Fcgamma receptors as regulators of immune responses. Nat. Rev. Immunol. 2008, 8, 34–47. [Google Scholar] [CrossRef]
- Vandendriessche, S.; Cambier, S.; Proost, P.; Marques, P.E. Complement receptors and their role in leukocyte recruitment and phagocytosis. Front. Cell Dev. Biol. 2021, 9, 624025. [Google Scholar] [CrossRef]
- Typiak, M.J.; Rębała, K.; Dudziak, M.; Dubaniewicz, A. Polymorphism of fcgr3a gene in sarcoidosis. Hum. Immunol. 2014, 75, 283–288. [Google Scholar] [CrossRef]
- Typiak, M.; Rębała, K.; Dudziak, M.; Słomiński, J.M.; Dubaniewicz, A. Polymorphism of fcgr2a, fcgr2c, and fcgr3b genes in the pathogenesis of sarcoidosis. Adv. Exp. Med. Biol. 2016, 905, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Typiak, M.; Rękawiecki, B.; Rębała, K.; Dubaniewicz, A. Comparative analysis of fcgr gene polymorphism in pulmonary sarcoidosis and tuberculosis. Cells 2023, 12, 1221. [Google Scholar] [CrossRef]
- Lambert, C.; Preijers, F.W.; Yanikkaya Demirel, G.; Sack, U. Monocytes and macrophages in flow: An escca initiative on advanced analyses of monocyte lineage using flow cytometry. Cytom. B Clin. Cytom 2017, 92, 180–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Lookeren Campagne, M.; Wiesmann, C.; Brown, E.J. Macrophage complement receptors and pathogen clearance. Cell Microbiol. 2007, 9, 2095–2102. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, F.; Gordan, S.; Lux, A. Fcγr dependent mechanisms of cytotoxic, agonistic, and neutralizing antibody activities. Trends Immunol. 2015, 36, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Pishesha, N.; Harmand, T.J.; Ploegh, H.L. A guide to antigen processing and presentation. Nat. Rev. Immunol. 2022, 22, 751–764. [Google Scholar] [CrossRef]
- Perez, R.L.; Rivera-Marrero, C.A.; Roman, J. Pulmonary granulomatous inflammation: From sarcoidosis to tuberculosis. Semin. Respir. Infect. 2003, 18, 23–32. [Google Scholar] [CrossRef]
- Baughman, R.P.; Culver, D.A.; Judson, M.A. A concise review of pulmonary sarcoidosis. Am. J. Respir. Crit. Care Med. 2011, 183, 573–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hijdra, D.; Vorselaars, A.D.; Crommelin, H.A.; van Moorsel, C.H.; Meek, B.; Claessen, A.M.; Rijkers, G.T.; Grutters, J.C. Can intermediate monocytes predict response to infliximab therapy in sarcoidosis? Eur. Respir. J. 2016, 48, 1242–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fingerle-Rowson, G.; Angstwurm, M.; Andreesen, R.; Ziegler-Heitbrock, H.W.L. Selective depletion of cd14+ cd16+ monocytes by glucocorticoid therapy. Clin. Exp. Immunol. 1998, 112, 501–506. [Google Scholar] [CrossRef]
- Vanham, G.; Edmonds, K.; Qing, L.; Hom, D.; Toossi, Z.; Jones, B.; Daley, C.L.; Huebner, R.; Kestens, L.; Gigase, P.; et al. Generalized immune activation in pulmonary tuberculosis: Co-activation with hiv infection. Clin. Exp. Immunol. 1996, 103, 30–34. [Google Scholar] [CrossRef]
- Yassin, R.J.; Hamblin, A.S. Altered expression of cd11/cd18 on the peripheral blood phagocytes of patients with tuberculosis. Clin. Exp. Immunol. 1994, 97, 120–125. [Google Scholar] [CrossRef]
- Gonzalez-Juarrero, M.; Shim, T.S.; Kipnis, A.; Junqueira-Kipnis, A.P.; Orme, I.M. Dynamics of macrophage cell populations during murine pulmonary tuberculosis. J. Immunol. 2003, 171, 3128–3135. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SA n = 24 (%) | TB n = 20 (%) | C n = 20 (%) | ||
|---|---|---|---|---|
| Age (years) | Mean ± SD | 36.6 ± 8.8 | 45.9 ± 16.7 | 33.9 ± 11 |
| Range | 23–56 | 30–60 | 20–54 | |
| Sex | Female | 10 (42) | 5 (25) | 10 (50) |
| Male | 14 (58) | 15 (75) | 10 (50) | |
| Smokers | 20 (83) | 16 (81) | 15 (75) | |
| BCG vaccinated | 24 (100) | 20 (100) | 20 (100) | |
| Positive PPD skin test | 0 | 20 (100) | 0 | |
| Löfgren’s syndrome | 0 | 0 | 0 | |
| Relapse | 0 | 0 | 0 | |
| Symptoms | Cough | 11 (46) | 18 (90) | 0 |
| Dyspnea | 0 | 5 (25) | 0 | |
| Increased body temperature | 4 (17) | 13 (65) | 0 | |
| Night sweats | 0 | 10 (50) | 0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Typiak, M.; Trzonkowski, P.; Skotarczak, M.; Dubaniewicz, A. Comparative Analysis of Fcγ and Complement Receptors Presence on Monocytes in Pulmonary Sarcoidosis and Tuberculosis. Int. J. Mol. Sci. 2023, 24, 9713. https://doi.org/10.3390/ijms24119713
Typiak M, Trzonkowski P, Skotarczak M, Dubaniewicz A. Comparative Analysis of Fcγ and Complement Receptors Presence on Monocytes in Pulmonary Sarcoidosis and Tuberculosis. International Journal of Molecular Sciences. 2023; 24(11):9713. https://doi.org/10.3390/ijms24119713
Chicago/Turabian StyleTypiak, Marlena, Piotr Trzonkowski, Monika Skotarczak, and Anna Dubaniewicz. 2023. "Comparative Analysis of Fcγ and Complement Receptors Presence on Monocytes in Pulmonary Sarcoidosis and Tuberculosis" International Journal of Molecular Sciences 24, no. 11: 9713. https://doi.org/10.3390/ijms24119713