
Comprehensive Analysis of YTH Domain-Containing Genes, Encoding m6A Reader and Their Response to Temperature Stresses and Yersinia ruckeri Infection in Rainbow Trout (Oncorhynchus mykiss)

Abstract
:1. Introduction
2. Results
2.1. Identification of YTH Domain-Containing Genes in Rainbow Trout
2.2. Phylogenetic Analysis of YTH Domain-Containing Genes
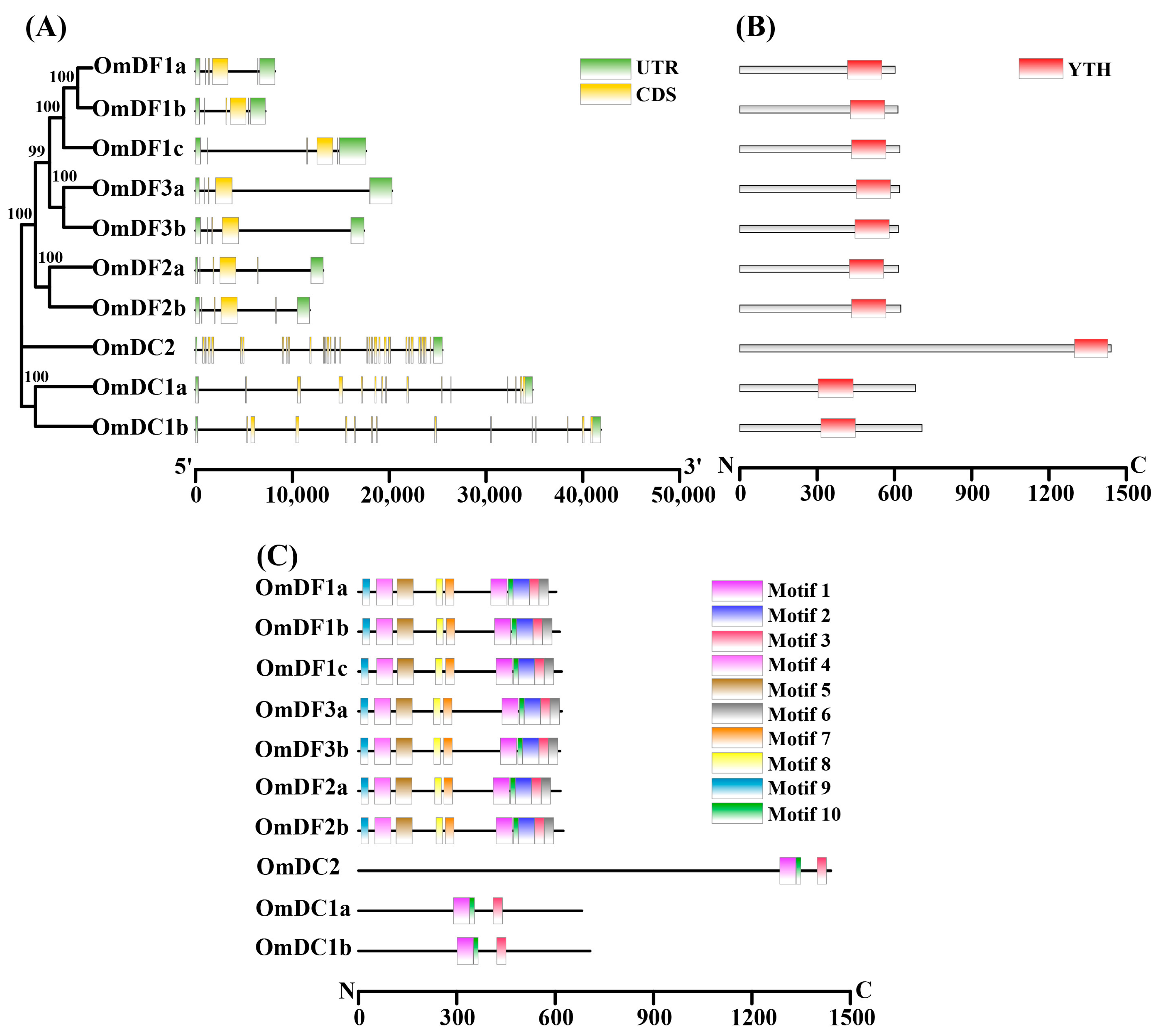
2.3. Gene Structure and Conserved Motif Analysis
2.4. Chromosome Locations and Synteny Relationships of YTH Domain-Containing Genes
2.5. Multiple Sequence Alignments and 3D Protein Structure Analysis
2.6. Expression Patterns of YTH Domain-Containing Genes under Different Temperatures
2.7. Expression Patterns of YTH Domain-Containing Genes after Y. ruckeri Infection
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Identification of YTH Domain-Containing Genes in Rrainbow Trout
4.3. Phylogenetic Analysis
4.4. Gene Structure, Chromosome Location and Syntenic Analysis
4.5. Three-Dimensional (3D) Protein Structure Analysis
4.6. Temperature Experiment
4.7. RNA Extraction and Quantitative Real-Time PCR (qPCR) Experiment
4.8. Bacterial Challenge and Bioinformatic Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Desrosiers, R.; Friderici, K.; Rottman, F. Identification of methylated nucleosides in messenger RNA from Novikoff hepatoma cells. Proc. Natl. Acad. Sci. USA 1974, 71, 3971–3975. [Google Scholar] [CrossRef] [PubMed]
- Roundtree, I.A.; Evans, M.E.; Pan, T.; He, C. Dynamic RNA modifications in gene expression regulation. Cell 2017, 169, 1187–1200. [Google Scholar] [CrossRef] [PubMed]
- Patil, D.P.; Pickering, B.F.; Jaffrey, S.R. Reading m6A in the Transcriptome: m6A-Binding Proteins. Trends Cell Biol. 2017, 28, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, S.; Xiao, W.; Zhao, Y.-L.; Yang, Y.-G. m6A: Signaling for mRNA splicing. RNA Biol. 2016, 13, 756–759. [Google Scholar] [CrossRef]
- Wei, L.H.; Song, P.; Wang, Y.; Lu, Z.; Tang, Q.; Yu, Q.; Xiao, Y.; Zhang, X.; Duan, H.C.; Jia, G. The m6A Reader ECT2 controls trichome morphology by affecting mRNA stability in Arabidopsis. Plant Cell 2018, 30, 968–985. [Google Scholar] [CrossRef]
- Meyer, K.D. m6A-mediated translation regulation. Biochim. Biophys. Acta-Gene Regul. Mech. 2018, 1862, 301–309. [Google Scholar] [CrossRef]
- Chen, L.; Fu, Y.; Hu, Z.; Deng, K.; Song, Z.; Liu, S.; Li, M.; Ou, X.; Wu, R.; Liu, M.; et al. Nuclear m6A reader YTHDC1 suppresses proximal alternative polyadenylation sites by interfering with the 3′ processing machinery. EMBO Rep. 2022, 23, e54686. [Google Scholar] [CrossRef]
- Zhao, B.S.; Wang, X.; Beadell, A.V.; Lu, Z.; Shi, H.; Kuuspalu, A.; Ho, R.K.; He, C. m6A-dependent maternal mRNA clearance facilitates zebrafish maternal-to-zygotic transition. Nature 2017, 542, 475–478. [Google Scholar] [CrossRef]
- Lence, T.; Akhtar, J.; Bayer, M.; Schmid, K.; Spindler, L.; Ho, C.H.; Kreim, N.; Andrade-Navarro, M.A.; Poeck, B.; Helm, M.; et al. m6A modulates neuronal functions and sex determination in Drosophila. Nature 2016, 540, 242–247. [Google Scholar] [CrossRef]
- Liu, S.; Li, Q.; Chen, K.; Zhang, Q.; Li, G.; Zhuo, L.; Zhai, B.; Sui, X.; Hu, X.; Xie, T. The emerging molecular mechanism of m6A modulators in tumorigenesis and cancer progression. Biomed. Pharmacother. 2020, 127, 110098. [Google Scholar] [CrossRef]
- Fry, N.J.; Law, B.A.; Ilkayeva, O.R.; Holley, C.L.; Mansfield, K.D. N6-methyladenosine is required for the hypoxic stabilization of specific mRNAs. RNA 2017, 23, 1444–1455. [Google Scholar] [CrossRef]
- Engel, M.; Eggert, C.; Kaplick, P.M.; Eder, M.; Röh, S.; Tietze, L.; Namendorf, C.; Arloth, J.; Weber, P.; Rex-Haffner, M.; et al. The Role of m6A/m-RNA Methylation in Stress Response Regulation. Neuron 2018, 99, 389–403.e9. [Google Scholar] [CrossRef]
- Meyer, K.D.; Jaffrey, S.R. Rethinking m6A Readers, Writers, and Erasers. Annu. Rev. Cell Dev. Biol. 2017, 33, 319–342. [Google Scholar] [CrossRef] [PubMed]
- Heck, A.M.; Wilusz, C.J. Small changes, big implications: The impact of m6A RNA methylation on gene expression in pluripotency and development. Biochim. Et Biophys. Acta BBA-Gene Regul. Mech. 2019, 1862, 194402. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhao, D.; Wu, J.; Shi, Y. Structure of the YTH domain of human YTHDF2 in complex with an m6A mononucleotide reveals an aromatic cage for m6A recognition. Cell Res. 2014, 24, 1490–1492. [Google Scholar] [CrossRef]
- Xu, C.; Wang, X.; Liu, K.; Roundtree, I.A.; Tempel, W.; Li, Y.; Lu, Z.; He, C.; Min, J. Structural basis for selective binding of m6A RNA by the YTHDC1 YTH domain. Nat. Chem. Biol. 2014, 10, 927–929. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, L. Deep analysis of RNA N6-adenosine methylation (m6A) patterns in human cells. NAR Genom. Bioinform. 2020, 2, lqaa007. [Google Scholar] [CrossRef]
- Kisan, A.; Chhabra, R. Modulation of gene expression by YTH domain family (YTHDF) proteins in human physiology and pathology. J. Cell. Physiol. 2023, 238, 5–31. [Google Scholar] [CrossRef]
- Li, L.; Krasnykov, K.; Homolka, D.; Gos, P.; Mendel, M.; Fish, R.J.; Pandey, R.R.; Pillai, R.S. The XRN1-regulated RNA helicase activity of YTHDC2 ensures mouse fertility independently of m6A recognition. Mol. Cell 2022, 82, 1678–1690.e12. [Google Scholar] [CrossRef]
- Larivera, S.; Meister, G. Domain confusion 2: m6A-independent role of YTHDC2. Mol. Cell 2022, 82, 1608–1609. [Google Scholar] [CrossRef]
- Berlivet, S.; Scutenaire, J.; Deragon, J.-M.; Bousquet-Antonelli, C. Readers of the m6A epitranscriptomic code. Biochim. et Biophys. Acta BBA-Gene Regul. Mech. 2019, 1862, 329–342. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, W.; Shen, F.; Yang, X.; Liu, H.; Dai, S.; Sun, X.; Huang, J.; Guo, Q. YTH domain proteins: A Family of m6A readers in cancer progression. Front. Oncol. 2021, 11, 629560. [Google Scholar] [CrossRef]
- Du, H.; Zhao, Y.; He, J.; Zhang, Y.; Xi, H.; Liu, M.; Ma, J.; Wu, L. YTHDF2 destabilizes m6A-containing RNA through direct recruitment of the CCR4-NOT deadenylase complex. Nat. Commun. 2016, 5, 12626. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Wang, X.; Lu, Z.; Zhao, B.S.; Ma, H.; Hsu, P.J.; Liu, C.; He, C. YTHDF3 facilitates translation and decay of N6-methyladenosine-modified RNA. Cell Res. 2017, 27, 315–328. [Google Scholar] [CrossRef]
- Kasowitz, S.; Ma, J.; Anderson, S.J.; Leu, N.A.; Xu, Y.; Gregory, B.D.; Schultz, R.M.; Wang, P.J. Nuclear m6A reader YTHDC1 regulates alternative polyadenylation and splicing during mouse oocyte development. PLOS Genet. 2018, 14, e1007412. [Google Scholar] [CrossRef]
- Hsu, P.J.; Zhu, Y.; Ma, H.; Guo, Y.; Shi, X.; Liu, Y.; Qi, M.; Lu, Z.; Shi, H.; Wang, J.; et al. Ythdc2 is an N6-methyladenosine binding protein that regulates mammalian spermatogenesis. Cell Res. 2017, 27, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, J.; Rao, H.; Hackert, P.; Sloan, K.E.; Höbartner, C.; Bohnsack, M.T. The m6A reader protein YTHDC2 interacts with the small ribosomal subunit and the 5′–3′ exoribonuclease XRN1. RNA 2018, 24, 1339–1350. [Google Scholar] [CrossRef] [PubMed]
- Xiao, K.; Liu, P.; Yan, P.; Liu, Y.; Song, L.; Liu, Y.; Xie, L. N6-methyladenosine reader YTH N6-methyladenosine RNA binding protein 3 or insulin like growth factor 2 mRNA binding protein 2 knockdown protects human bronchial epithelial cells from hypoxia/reoxygenation injury by inactivating p38 MAPK, AKT, ERK1/2, and NF-κB pathways. Bioengineered 2022, 13, 11973–11986. [Google Scholar] [CrossRef]
- Weng, Y.-L.; Wang, X.; An, R.; Cassin, J.; Vissers, C.; Liu, Y.; Liu, Y.; Xu, T.; Wang, X.; Wong, S.Z.H.; et al. Epitranscriptomic m6A Regulation of Axon Regeneration in the Adult Mammalian Nervous System. Neuron 2018, 97, 313–325.e6. [Google Scholar] [CrossRef]
- Millay, D.P.; O’rourke, J.R.; Sutherland, L.B.; Bezprozvannaya, S.; Shelton, J.M.; Bassel-Duby, R.; Olson, E.N. Myomaker is a membrane activator of myoblast fusion and muscle formation. Nature 2013, 499, 301–305. [Google Scholar] [CrossRef]
- Yang, X.; Wang, J.; Ma, X.; Du, J.; Mei, C.; Zan, L. Transcriptome-wide N6-Methyladenosine Methylome Profiling Reveals m6A Regulation of Skeletal Myoblast Differentiation in Cattle (Bos taurus). Front. Cell Dev. Biol. 2021, 9, 785380. [Google Scholar] [CrossRef]
- Yu, J.; Li, Y.; Wang, T.; Zhong, X. Modification of N6-methyladenosine RNA methylation on heat shock protein expression. PLoS ONE 2018, 13, e0198604. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Zong, X.; Mao, Y.; Qian, S.B. A heat shock-responsive lncRNA Heat acts as a HSF1-directed transcriptional brake via m6A modification. Proc. Natl. Acad. Sci. USA 2021, 118, e2102175118. [Google Scholar] [CrossRef]
- Lu, Z.; Ma, Y.; Li, Q.; Liu, E.; Jin, M.; Zhang, L.; Wei, C. The role of N6-methyladenosine RNA methylation in the heat stress response of sheep (Ovis aries). Cell Stress Chaperones 2019, 24, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Zhang, Y.; Zhang, J.; Wang, J.; Li, Q. The alteration of N6-methyladenosine (m6A) modification at the transcriptome-wide level in response of heat stress in bovine mammary epithelial cells. BMC Genom. 2022, 23, 829. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, Z.; Zou, C.; Liang, S.; Zou, Y.; Liu, Y.; You, F. Sex-dependent RNA editing and N6-adenosine RNA methylation profiling in the gonads of a fish, the olive flounder (Paralichthys olivaceus). Front. Cell Dev. Biol. 2020, 8, 751. [Google Scholar] [CrossRef]
- Thorgaard, G.H.; Bailey, G.S.; Williams, D.; Buhler, D.R.; Kaattari, S.L.; Ristow, S.S.; Hansen, J.D.; Winton, J.R.; Bartholomew, J.L.; Nagler, J.J.; et al. Status and opportunities for genomics research with rainbow trout. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2002, 133, 609–646. [Google Scholar] [CrossRef]
- Bailey, G.S.; Williams, D.E.; Hendricks, J.D. Fish models for environmental carcinogenesis: The rainbow trout. Environ. Health Perspect. 1996, 104, 5–21. [Google Scholar] [CrossRef]
- Berthelot, C.; Brunet, F.; Chalopin, D.; Juanchich, A.; Bernard, M.; Noël, B.; Bento, P.; Da Silva, C.; Labadie, K.; Alberti, A.; et al. The rainbow trout genome provides novel insights into evolution after whole-genome duplication in vertebrates. Nat. Commun. 2014, 5, 3657. [Google Scholar] [CrossRef]
- Aas, T.S.; Åsgård, T.; Ytrestøyl, T. Chemical composition of whole body and fillet of slaughter sized Atlantic salmon (Salmo salar) and rainbow trout (Oncorhynchus mykiss) farmed in Norway in 2020. Aquac. Rep. 2022, 25, 101252. [Google Scholar] [CrossRef]
- Tian, Y.; Gao, Q.; Dong, S.; Zhou, Y.; Yu, H.; Liu, D.; Yang, W. Genome-wide analysis of alternative splicing (AS) mechanism provides insights into salinity adaptation in the livers of three euryhaline teleosts, including Scophthalmus maximus, Cynoglossus semilaevis and Oncorhynchus mykiss. Biology 2022, 11, 222. [Google Scholar] [CrossRef]
- Tian, Y.; Gao, Q.; Yu, H.; Liu, D.; Dong, S.; Zhou, Y.; Yang, W.; Xue, N.; Bao, H.; Yu, Y. Dynamic transcriptome and LC-MS/MS analysis revealed the important roles of taurine and glutamine metabolism in response to environmental salinity changes in gills of rainbow trout (Oncorhynchus mykiss). Int. J. Biol. Macromol. 2022, 221, 1545–1557. [Google Scholar] [CrossRef] [PubMed]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Ao, Q.; Tan, C.; Yang, Y. Genome-wide identification and characterization of YTH domain-containing genes, encoding the m6A readers, and their expression in tomato. Plant Cell Rep. 2021, 40, 1229–1245. [Google Scholar] [CrossRef] [PubMed]
- Zaccara, S.; Ries, R.J.; Jaffrey, S.R. Reading, writing and erasing mRNA methylation. Nat. Rev. Mol. Cell Biol. 2019, 20, 608–624. [Google Scholar] [CrossRef] [PubMed]
- Ehsan, P.A.; Morgane, F.; Mikaela, H.; Craig, R.P. Expression of m6A RNA methylation markers in the hypothalamus of Atlantic salmon. bioRxiv 2022, Preprint, 1–26. [Google Scholar] [CrossRef]
- Kontur, C.; Jeong, M.; Cifuentes, D.; Giraldez, A.J. Ythdf m6A Readers Function Redundantly during Zebrafish Development. Cell Rep. 2020, 33, 108598. [Google Scholar] [CrossRef]
- Li, Y.; Bedi, R.K.; Moroz-Omori, E.V.; Caflisch, A. Structural and dynamic insights into redundant function of YTHDF proteins. J. Chem. Inf. Model. 2020, 60, 5932–5935. [Google Scholar] [CrossRef]
- Xu, C.; Liu, K.; Ahmed, H.; Loppnau, P.; Schapira, M.; Min, J. Structural basis for the discriminative recognition of N6-methyladenosine RNA by the human YT521-B homology domain family of proteins. J. Biol. Chem. 2015, 290, 24902–24913. [Google Scholar] [CrossRef]
- Lasman, L.; Krupalnik, V.; Viukov, S.; Mor, N.; Aguilera-Castrejon, A.; Schneir, D.; Bayerl, J.; Mizrahi, O.; Peles, S.; Tawil, S.; et al. Context-dependent functional compensation between Ythdf m6A reader proteins. Genes Dev. 2020, 34, 1373–1391. [Google Scholar] [CrossRef]
- Beitinger, T.L.; Bennett, W.A.; McCauley, R.W. Temperature Tolerances of North American Freshwater Fishes Exposed to Dynamic Changes in Temperature. Environ. Biol. Fishes 2000, 58, 237–275. [Google Scholar] [CrossRef]
- Alfonso, S.; Gesto, M.; Sadoul, B. Temperature increase and its effects on fish stress physiology in the context of global warming. J. Fish Biol. 2021, 98, 1496–1508. [Google Scholar] [CrossRef] [PubMed]
- Theler, D.; Dominguez, C.; Blatter, M.; Boudet, J.; Allain, F.H. Solution structure of the YTH domain in complex with N6-methyladenosine RNA: A reader of methylated RNA. Nucleic Acids Res. 2014, 42, 13911–13919. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wan, J.; Gao, X.; Zhang, X.; Jaffrey, S.R.; Qian, S.B. Dynamic m6A mRNA methylation directs translational control of heat shock response. Nature 2015, 26, 591–594. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.D.; Patil, D.P.; Zhou, J.; Zinoviev, A.; Skabkin, M.A.; Elemento, O.; Pestova, T.V.; Qian, S.-B.; Jaffrey, S.R. 5’ UTR m6A Promotes Cap-Independent Translation. Cell 2015, 163, 999–1010. [Google Scholar] [CrossRef]
- Patil, D.P.; Chen, C.-K.; Pickering, B.F.; Chow, A.; Jackson, C.; Guttman, M.; Jaffrey, S.R. m6A RNA methylation promotes XIST-mediated transcriptional repression. Nature 2016, 537, 369–373. [Google Scholar] [CrossRef]
- Timcheva, K. Role of YTHDC1, Nuclear Reader of N6-Methyladenosine (m6A) RNA Modification, in the Regulation of the Cellular Heat Stress Response. Ph.D. Thesis, Université Grenoble Alpes, Grenoble, France, 2020. Available online: https://theses.hal.science/tel-03793808 (accessed on 25 May 2016).
- Zong, X.; Xiao, X.; Shen, B.; Jiang, Q.; Wang, H.; Lu, Z.; Wang, F.; Jin, M.; Min, J.; Wang, F.; et al. The N 6-methyladenosine RNA-binding protein YTHDF1 modulates the translation of TRAF6 to mediate the intestinal immune response. Nucleic Acids Res. 2021, 49, 5537–5552. [Google Scholar] [CrossRef]
- Hao, W.-Y.; Lou, Y.; Hu, G.-Y.; Qian, C.-Y.; Liang, W.-R.; Zhao, J.; Wang, X.-H. RNA m6A reader YTHDF1 facilitates inflammation via enhancing NLRP3 translation. Biochem. Biophys. Res. Commun. 2022, 616, 76–81. [Google Scholar] [CrossRef]
- Xing, Y.; Cheng, D.; Shi, C.; Shen, Z. The protective role of YTHDF1-knock down macrophages on the immune paralysis of severe sepsis rats with ECMO. Microvasc. Res. 2021, 137, 104178. [Google Scholar] [CrossRef]
- Chou, K.-C.; Shen, H.-B. Cell-PLoc: A package of Web servers for predicting subcellular localization of proteins in various organisms. Nat. Protoc. 2008, 3, 153–162. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.; Sun, H.; Xu, C. YTH domain: A family of N6-methyladenosine (m6A) readers. Genom. Proteom. Bioinform. 2018, 16, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Sun, S.; Li, S.; Lu, T.; Shi, D. Transcriptome profiling of immune response to Yersinia ruckeri in spleen of rainbow trout (Oncorhynchus mykiss). BMC Genom. 2021, 22, 292. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Chromosome Position | Gene Length (bp) | ORF (bp) | Exon Number | CDS (aa) | Molecular Weight (kD) | Theoretical pI | Putative Localization | GenBank Accession Number | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Nucleus | Cytoplasm | ||||||||||
| OmDF1a | LOC110492754 | Chr16(−): 46478316–46486506 | 8191 | 1812 | 6 | 603 | 65,089.20 | 7.82 | − | + | OQ801191 |
| OmDF1b | LOC110532506 | Chr09(+): 58975469–58982674 | 7206 | 1845 | 6 | 614 | 66,138.59 | 8.24 | − | + | OQ801192 |
| OmDF1c | LOC110492496 | Chr16(−): 61779118–61796715 | 17,598 | 1863 | 6 | 620 | 67,739.79 | 8.92 | − | + | OQ801193 |
| OmDF2a | LOC110505278 | Chr25(−): 6680033–6693201 | 13,169 | 1851 | 6 | 616 | 66,434.05 | 8.92 | − | + | OQ801194 |
| OmDF2b | LOC110498162 | Chr19(+): 56850905–56862683 | 11,779 | 1878 | 6 | 625 | 67,365.05 | 8.84 | − | + | OQ801195 |
| OmDF3a | LOC110489655 | Chr15(+): 15345480–15365760 | 20,281 | 1863 | 5 | 620 | 68,462.39 | 8.89 | − | + | OQ801196 |
| OmDF3b | LOC110535333 | Chr11(−): 15251271–15268657 | 17,387 | 1848 | 5 | 615 | 67,827.79 | 8.97 | − | + | OQ801197 |
| OmDC1a | LOC110523074 | Chr05(−): 724584–759382 | 34,799 | 2049 | 15 | 682 | 78,915.25 | 7.84 | + | − | OQ801198 |
| OmDC1b | LOC110536768 | Chr12(−): 5222326–5264164 | 41,839 | 2124 | 15 | 707 | 81,222.87 | 9.45 | + | − | OQ801199 |
| OmDC2 | LOC110509841 | ChrY(−): 23853359–23878838 | 25,480 | 4326 | 32 | 1441 | 160,900.86 | 8.65 | + | − | OQ801200 |
| Motif | Length (aa) | Sequence |
|---|---|---|
| 1 | 50 | YNPKDFDWNLKNGRVFIIKSYSEDDIHRSIKYSIWCSTEHGNKRLD-GAYR |
| 2 | 50 | HFCGVAEMRSPVDYNTSAGVWSQDKWKGKFDVDWLFVKDVPNSQLRHIRL |
| 3 | 29 | ENNDNKPVTNSRDTQEVPLEKAKQVLKII |
| 4 | 50 | MSDPYLPSYYAPSIGFPYSLSEAPWSTGGDPPMPYLPYGQLSNGEH-HFM |
| 5 | 50 | FJYQHGFNFFPENPDFSAWGTSGSQGQSTQSSAYSGSYSYPPSSLG-GAJV |
| 6 | 29 | AGFKHTTSIFDDFSHYEKRQEEEEAVRKT |
| 7 | 27 | GGALPPPPIKHNMDIGTWDNKGSMNKV |
| 8 | 21 | PPKPTSWAAIASKPAKPQPKK |
| 9 | 23 | QRPKGQGNKVQNGSLHQKETVND |
| 10 | 15 | AKGPVYLLFSVNGSG |
| Gene | Primer Sequence (5′ to 3′) |
|---|---|
| OmDF1a | Forward (F): CCAACTCTAGGGACACTCAA Reverse (R): GCTGGCTCATAGGTCTTTCT |
| OmDF1b | Forward (F): TGGACTGGCTGTTTGTGAAAG Reverse (R): TCGGGACCCTCGCTGTATT |
| OmDF1c | Forward (F): ATCCGCCTTGAGAACAACGAC Reverse (R): CCTCCTCTTCCTCCTGCCTCT |
| OmDF2a | Forward (F): CCCCTTTCACCCAGAACGAGC Reverse (R): TGTCCCAAGTGCCGATGTCC |
| OmDF2b | Forward (F): CACCGCTCCATCAAGTACAACATC Reverse (R): TCTCCGCCACGCCACAAA |
| OmDF3a | Forward (F): GTCAACGGCAGTGGTCATTTCT Reverse (R): TTGGGCACGTCCTTCACAAA |
| OmDF3b | Forward (F): GGTGAAGTGGGCGTTTGTGA Reverse (R): AGGTGGTATGCTTGAAAGTGGC |
| OmDC1a | Forward (F): GGCTGGCGGGAAGAAACT Reverse (R): CAAGGTGGGCTGCTTTGG |
| OmDC1b | Forward (F): GGGATGAACGCCAAGATGC Reverse (R): GCGAGGGTCCAGTGTAAAGC |
| OmDC2 | Forward (F): CCAAAGCACCCGAACCAC Reverse (R): GAGCACCACGGCACATAACA |
| 18s rRNA | Forward (F): ATGGCCGTTCTTAGTTGGTG Reverse (R): TCAGTCTCGTGTGGCTGAAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, H.; Gao, Q.; Wang, W.; Liu, D.; He, J.; Tian, Y. Comprehensive Analysis of YTH Domain-Containing Genes, Encoding m6A Reader and Their Response to Temperature Stresses and Yersinia ruckeri Infection in Rainbow Trout (Oncorhynchus mykiss). Int. J. Mol. Sci. 2023, 24, 9348. https://doi.org/10.3390/ijms24119348
Yu H, Gao Q, Wang W, Liu D, He J, Tian Y. Comprehensive Analysis of YTH Domain-Containing Genes, Encoding m6A Reader and Their Response to Temperature Stresses and Yersinia ruckeri Infection in Rainbow Trout (Oncorhynchus mykiss). International Journal of Molecular Sciences. 2023; 24(11):9348. https://doi.org/10.3390/ijms24119348
Chicago/Turabian StyleYu, Han, Qinfeng Gao, Wen Wang, Dazhi Liu, Jinghong He, and Yuan Tian. 2023. "Comprehensive Analysis of YTH Domain-Containing Genes, Encoding m6A Reader and Their Response to Temperature Stresses and Yersinia ruckeri Infection in Rainbow Trout (Oncorhynchus mykiss)" International Journal of Molecular Sciences 24, no. 11: 9348. https://doi.org/10.3390/ijms24119348