

miR-3174 Is a New Tumor Suppressor MicroRNA That Inhibits Several Tumor-Promoting Genes in Glioblastoma
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
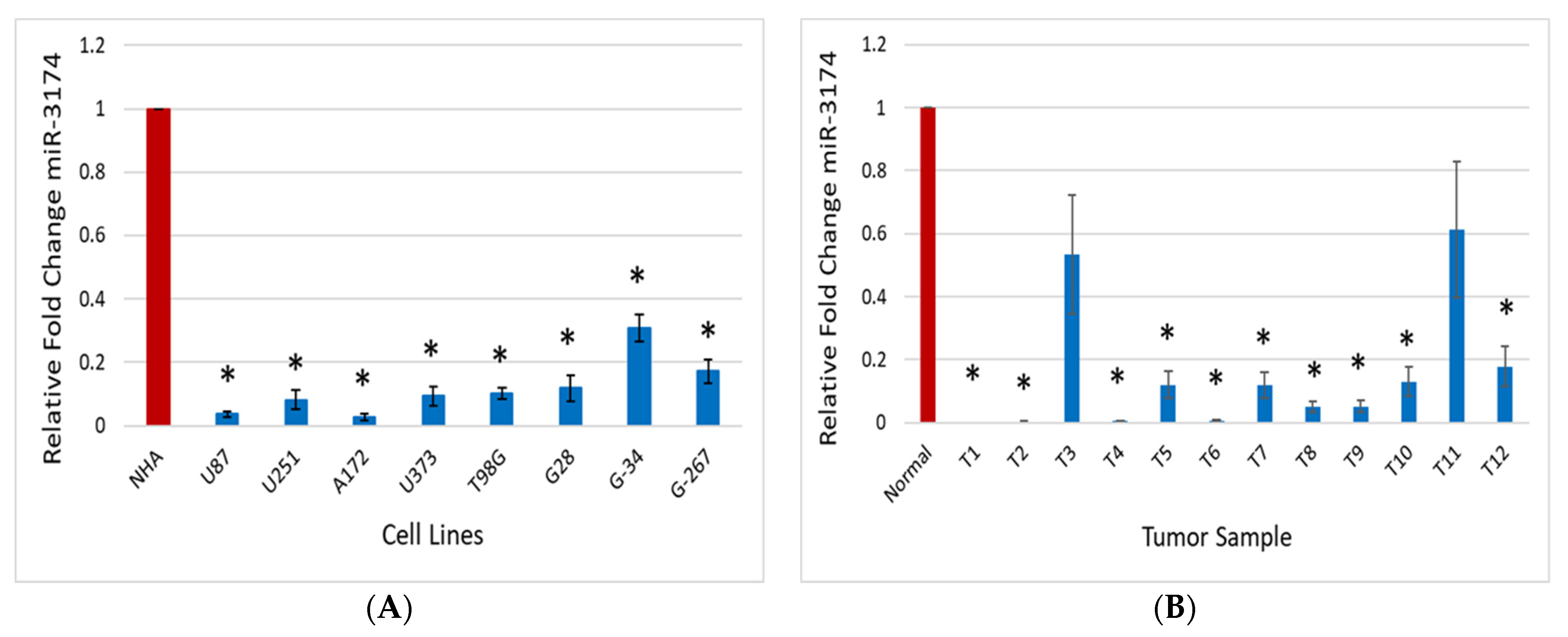
2.1. miR-3174 Expression Is Downregulated in GBM Tissues, Cell Lines and GSCs
2.2. miR-3174 Inhibits Proliferation and Invasion of GBM Cells
2.3. miR-3174 Impedes the Sphere Formation Ability of GSCs
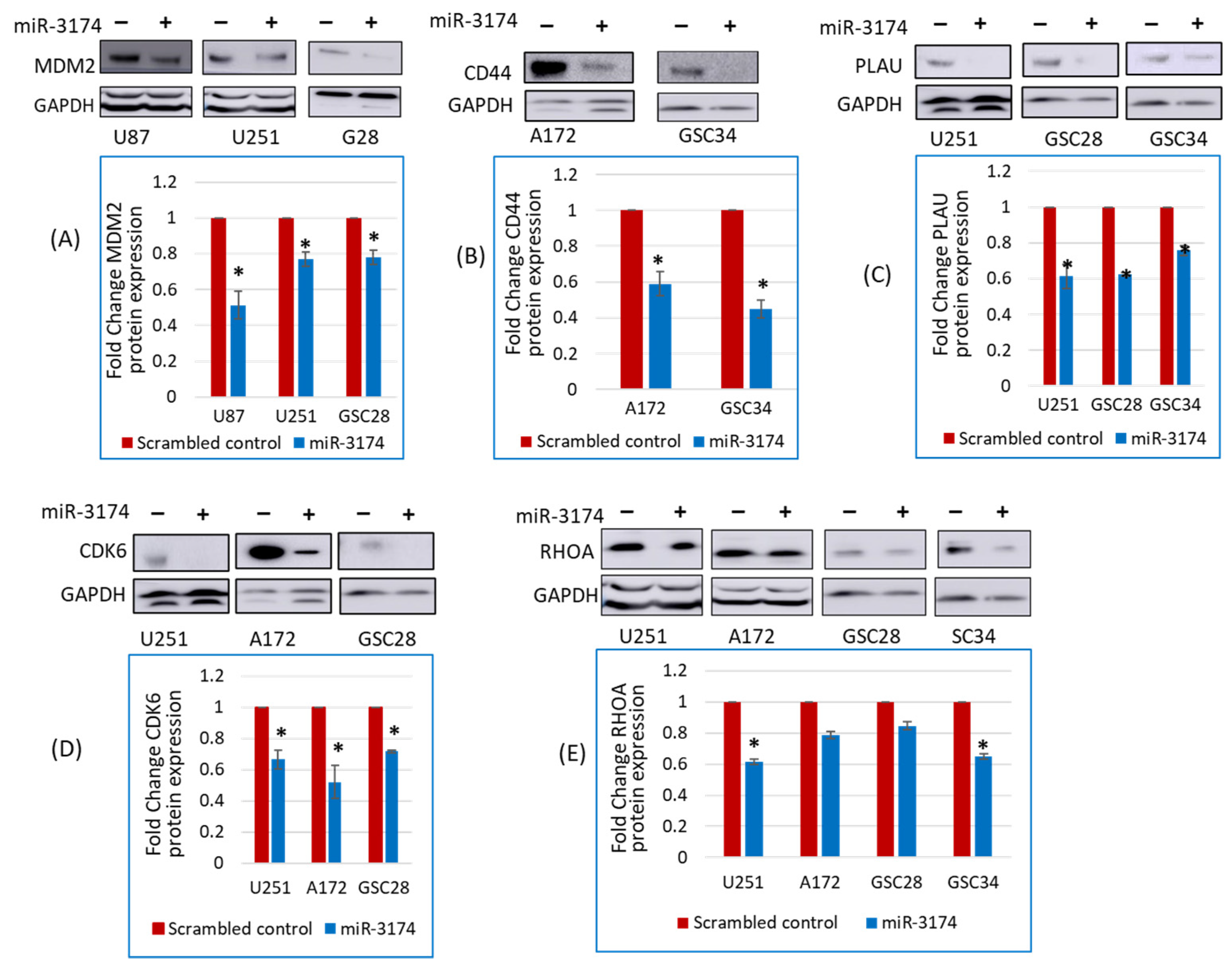
2.4. miR-3174 Negatively Regulates the Expression of Multiple Tumor-Promoting Genes
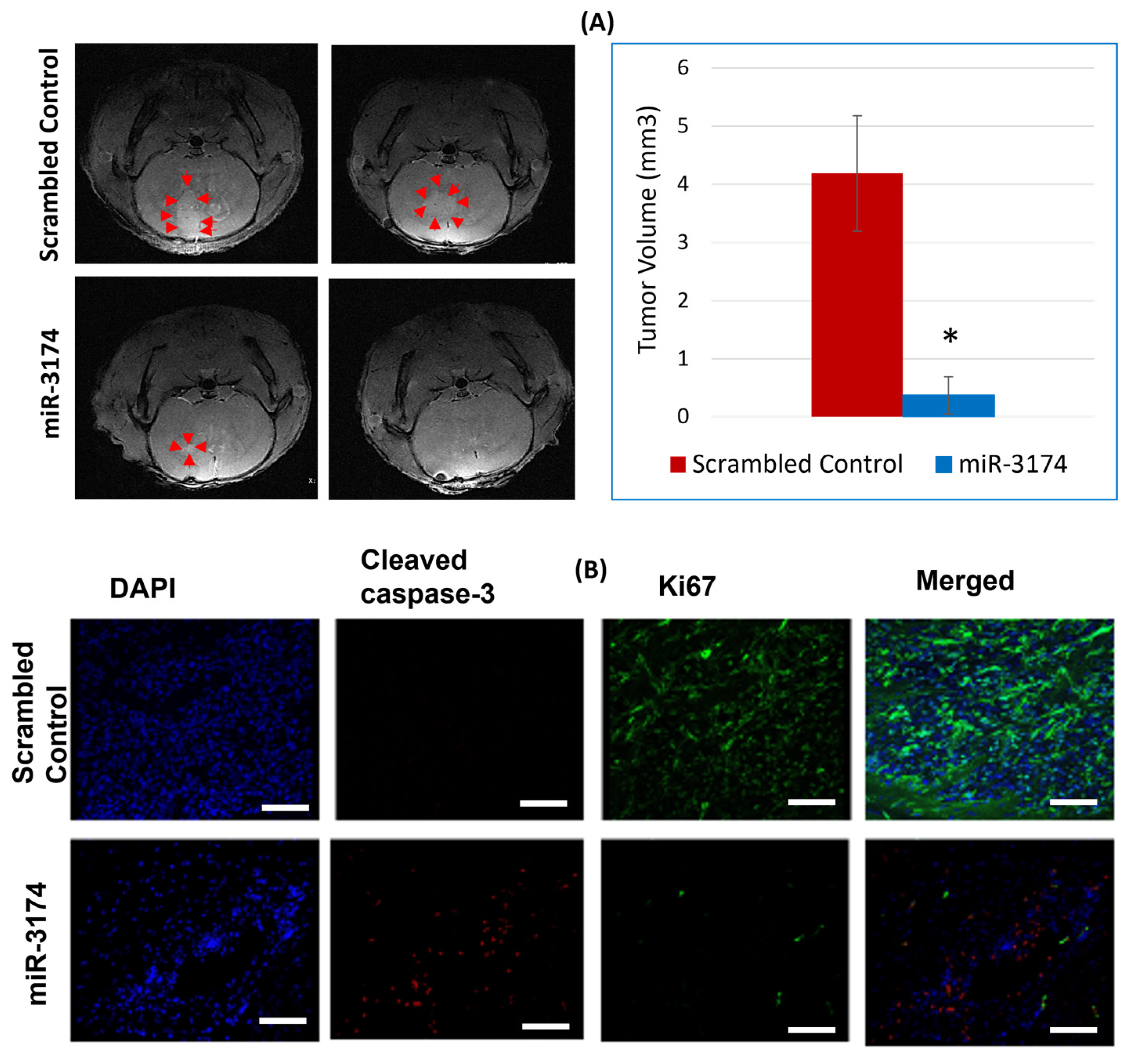
2.5. miR-3174 Suppresses GBM Growth in a Mouse Xenograft Model In Vivo
miR-3174 Promotes Cell Apoptosis and Inhibits Proliferation of Tumors In Vivo
3. Discussion
4. Materials and Methods
4.1. Tumor Specimens and Cell Lines
4.2. Reagents
4.3. Quantitative RT-PCR
4.4. Cell Growth Assay
4.5. Invasion Assay
4.6. Glioma Stem Cell Sphere Formation Assay
4.7. Immunoblot Analysis
4.8. In Vivo Experiments
4.9. Immuno-Histochemical Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-coding RNAs and their integrated networks. J. Integr. Bioinform. 2019, 16, 20190027. [Google Scholar] [CrossRef] [PubMed]
- Slack, F.J.; Chinnaiyan, A.M. The role of non-coding RNAs in oncology. Cell 2019, 179, 1033–1055. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy, E.Q.; Zhang, Y.; Colόn, R.R.; Cain, S.R.; Gibert, M.K., Jr.; Dube, C.J.; Hafner, M.; Abounader, R. MicroRNA 3928 suppresses glioblastoma through downregulation of several oncogenes and upregulation of p53. Int. J. Mol. Sci. 2022, 23, 3930. [Google Scholar] [CrossRef] [PubMed]
- Smolarz, B.; Durczynski, A.; Romanowicz, H.; Szyłło, K.; Hogendorf, P. MiRNAs in Cancer. Int. J. Mol. Sci 2022, 23, 2805. [Google Scholar] [CrossRef] [PubMed]
- Abadi, A.J.; Zarrabi, A.; Gholami, M.H.; Mirzaei, S.; Hashemi, F.; Zabolian, A.; Entezari, M.; Hushmandi, K.; Ashrafizadeh, M.; Khan, H. Small in size, but large in action: microRNAs as potential modulators of PTEN in breast and lung cancers. Biomolecules 2021, 11, 304. [Google Scholar] [CrossRef]
- Yeh, M.; Wang, Y.-Y.; Yoo, J.Y.; Oh, C.; Otani, Y.; Kang, J.M.; Park, E.S.; Kim, E.; Chung, S.; Jeon, Y.-J. MicroRNA-138 suppresses glioblastoma proliferation through downregulation of CD44. Sci. Rep. 2021, 11, 9219. [Google Scholar] [CrossRef]
- Hanif, F.; Muzaffar, K.; Perveen, K.; Malhi, S.M.; Simjee, S.U. Glioblastoma multiforme: A review of its epidemiology and pathogenesis through clinical presentation and treatment. Asian Pac. J. Cancer Prev. APJCP 2017, 18, 3. [Google Scholar]
- van Solinge, T.S.; Nieland, L.; Chiocca, E.A.; Broekman, M.L. Advances in local therapy for glioblastoma—Taking the fight to the tumour. Nat. Rev. Neurol. 2022, 18, 221–236. [Google Scholar] [CrossRef]
- DeCordova, S.; Shastri, A.; Tsolaki, A.G.; Yasmin, H.; Klein, L.; Singh, S.K.; Kishore, U. Molecular heterogeneity and immunosuppressive microenvironment in glioblastoma. Front. Immunol. 2020, 11, 1402. [Google Scholar] [CrossRef]
- Auffinger, B.; Spencer, D.; Pytel, P.; Ahmed, A.U.; Lesniak, M.S. The role of glioma stem cells in chemotherapy resistance and glioblastoma multiforme recurrence. Expert Rev. Neurother. 2015, 15, 741–752. [Google Scholar] [CrossRef]
- Toraih, E.A.; Aly, N.M.; Abdallah, H.Y.; Al-Qahtani, S.A.; Shaalan, A.A.; Hussein, M.H.; Fawzy, M.S. MicroRNA–target cross-talks: Key players in glioblastoma multiforme. Tumor Biol. 2017, 39, 1010428317726842. [Google Scholar] [CrossRef]
- Tang, Y.; Zong, S.; Zeng, H.; Ruan, X.; Yao, L.; Han, S.; Hou, F. MicroRNAs and angiogenesis: A new era for the management of colorectal cancer. Cancer Cell Int. 2021, 21, 221. [Google Scholar] [CrossRef] [PubMed]
- Balandeh, E.; Mohammadshafie, K.; Mahmoudi, Y.; Hossein Pourhanifeh, M.; Rajabi, A.; Bahabadi, Z.R.; Mohammadi, A.H.; Rahimian, N.; Hamblin, M.R.; Mirzaei, H. Roles of non-coding RNAs and angiogenesis in glioblastoma. Front. Cell Dev. Biol. 2021, 2021, 2543. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Guessous, F.; Zhang, Y.; DiPierro, C.; Kefas, B.; Johnson, E.; Marcinkiewicz, L.; Jiang, J.; Yang, Y.; Schmittgen, T.D. MicroRNA-34a inhibits glioblastoma growth by targeting multiple oncogenes. Cancer Res. 2009, 69, 7569–7576. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Dodbele, S.; Park, T.; Glass, R.; Bhat, K.; Sulman, E.P.; Zhang, Y.; Abounader, R. MicroRNA-29a inhibits glioblastoma stem cells and tumor growth by regulating the PDGF pathway. J. Neuro-Oncol. 2019, 145, 23–34. [Google Scholar] [CrossRef]
- Breunig, C.; Pahl, J.; Küblbeck, M.; Miller, M.; Antonelli, D.; Erdem, N.; Wirth, C.; Will, R.; Bott, A.; Cerwenka, A. MicroRNA-519a-3p mediates apoptosis resistance in breast cancer cells and their escape from recognition by natural killer cells. Cell Death Dis. 2017, 8, e2973. [Google Scholar] [CrossRef]
- Liu, J.; M Albrecht, A.; Ni, X.; Yang, J.; Li, M. Glioblastoma tumor initiating cells: Therapeutic strategies targeting apoptosis and microRNA pathways. Curr. Mol. Med. 2013, 13, 352–357. [Google Scholar]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or tumor suppressor? The duplicity of microRNAs in cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef]
- Li, Q.; Shen, J.; Li, J.; Dai, D.; Saeed, M.; Li, C. LINC00958/miR-3174/PHF6 axis is responsible for triggering proliferation, migration and invasion of endometrial cancer. Eur. Rev. Med. Pharm. Sci. 2021, 25, 6853–6861. [Google Scholar]
- Guo, J.; Liu, Y.; Zhao, P. Bioinformatic Analysis Identified Potentially Prognostic Long Noncoding RNAs and MicroRNAs for Gastric Cancer. BioMed Res. Int. 2021, 2021, 6683136. [Google Scholar] [CrossRef]
- Yu, C.; Wang, Y.; Liu, T.; Sha, K.; Song, Z.; Zhao, M.; Wang, X. The microRNA miR-3174 suppresses the expression of ADAM15 and inhibits the proliferation of patient-derived bladder cancer cells. OncoTargets Ther. 2020, 13, 4157. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Ma, W. Cisatracurium inhibits the proliferation, migration and invasion of breast cancer cells by regulating the expression of miR-3174. Cell. Mol. Biol. 2020, 66, 1–7. [Google Scholar] [CrossRef]
- Zhang, D.; Yang, N. MiR-3174 functions as an oncogene in rectal cancer by targeting PCBD2. Eur. Rev. Med. Pharm. Sci. 2019, 23, 2417–2426. [Google Scholar]
- Wang, Q.; Yang, X.; Zhou, X.; Wu, B.; Zhu, D.; Jia, W.; Chu, J.; Wang, J.; Wu, J.; Kong, L. MiR-3174 promotes proliferation and inhibits apoptosis by targeting FOXO1 in hepatocellular carcinoma. Biochem. Biophys. Res. Commun. 2020, 526, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zuo, X.; Wei, D. Concise review: Emerging role of CD44 in cancer stem cells: A promising biomarker and therapeutic target. Stem Cells Transl. Med. 2015, 4, 1033–1043. [Google Scholar] [CrossRef]
- Nebenfuehr, S.; Kollmann, K.; Sexl, V. The role of CDK6 in cancer. Int. J. Cancer 2020, 147, 2988–2995. [Google Scholar] [CrossRef] [PubMed]
- YP Lam, E.; di Tomaso, H.-K.; Ng, J.C.S.; Pang, M.F.; Roussel, N.M.; Hjelm, P. Expression of p19 INK4d, CDK4, CDK6 in glioblastoma multiforme. Br. J. Neurosurg. 2000, 14, 28–32. [Google Scholar]
- Hou, H.; Sun, D.; Zhang, X. The role of MDM2 amplification and overexpression in therapeutic resistance of malignant tumors. Cancer Cell Int. 2019, 19, 216. [Google Scholar] [CrossRef]
- Chen, G.; Sun, J.; Xie, M.; Yu, S.; Tang, Q.; Chen, L. PLAU promotes cell proliferation and epithelial-mesenchymal transition in head and neck squamous cell carcinoma. Front. Genet. 2021, 12, 651882. [Google Scholar] [CrossRef]
- Al-Koussa, H.; Atat, O.E.; Jaafar, L.; Tashjian, H.; El-Sibai, M. The role of Rho GTPases in motility and invasion of glioblastoma cells. Anal. Cell. Pathol. 2020, 2020, 9274016. [Google Scholar] [CrossRef]
- Sinn, H.-P.; Heider, K.-H.; Skroch-Angel, P.; von Minckwitz, G.; Kaufmann, M.; Herrlich, P.; Ponta, H. Human mammary carcinomas express homologues of rat metastasis-associated variants of CD44. Breast Cancer Res. Treat. 1995, 36, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Tölg, C.; Hofmann, M.; Herrlich, P.; Ponta, H. Splicing choice from ten variant exons establishes CD44 variability. Nucleic Acids Res. 1993, 21, 1225–1229. [Google Scholar] [CrossRef] [PubMed]
- Eibl, R.H.; Pietsch, T.; Moll, J.; Skroch-Angel, P.; Heider, K.-H.; von Ammon, K.; Wiestler, O.D.; Ponta, H.; Kleihues, P.; Herrlich, P. Expression of variant CD44 epitopes in human astrocytic brain tumors. J. Neuro-Oncol. 1995, 26, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhao, S.; Karnad, A.; Freeman, J.W. The biology and role of CD44 in cancer progression: Therapeutic implications. J. Hematol. Oncol. 2018, 11, 64. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-Y.; Huang, C.-S.; Yang, Y.-P.; Liu, C.-Y.; Liu, Y.-Y.; Wu, W.-W.; Lu, K.-H.; Chen, K.-H.; Chang, Y.-L.; Lee, S.-D. The subpopulation of CD44-positive cells promoted tumorigenicity and metastatic ability in lung adenocarcinoma. J. Chin. Med. Assoc. 2019, 82, 196–201. [Google Scholar] [CrossRef]
- Xu, H.; Wu, K.; Tian, Y.; Liu, Q.; Han, N.; Yuan, X.; Zhang, L.; Wu, G.S.; Wu, K. CD44 correlates with clinicopathological characteristics and is upregulated by EGFR in breast cancer. Int. J. Oncol. 2016, 49, 1343–1350. [Google Scholar] [CrossRef]
- Liu, T.; Yan, Z.; Liu, Y.; Choy, E.; Hornicek, F.J.; Mankin, H.; Duan, Z. CRISPR-Cas9-mediated silencing of CD44 in human highly metastatic osteosarcoma cells. Cell. Physiol. Biochem. 2018, 46, 1218–1230. [Google Scholar] [CrossRef]
- Sadeghi, A.; Roudi, R.; Mirzaei, A.; Zare Mirzaei, A.; Madjd, Z.; Abolhasani, M. CD44 epithelial isoform inversely associates with invasive characteristics of colorectal cancer. Biomark. Med. 2019, 13, 419–426. [Google Scholar] [CrossRef]
- Hou, C.; Ishi, Y.; Motegi, H.; Okamoto, M.; Ou, Y.; Chen, J.; Yamaguchi, S. Overexpression of CD44 is associated with a poor prognosis in grade II/III gliomas. J. Neuro-Oncol. 2019, 145, 201–210. [Google Scholar] [CrossRef]
- Kolliopoulos, C.; Ali, M.M.; Castillejo-Lopez, C.; Heldin, C.-H.; Heldin, P. CD44 Depletion in Glioblastoma Cells Suppresses Growth and Stemness and Induces Senescence. Cancers 2022, 14, 3747. [Google Scholar] [CrossRef]
- Bond, G.L.; Hu, W.; Bond, E.E.; Robins, H.; Lutzker, S.G.; Arva, N.C.; Bargonetti, J.; Bartel, F.; Taubert, H.; Wuerl, P. A single nucleotide polymorphism in the MDM2 promoter attenuates the p53 tumor suppressor pathway and accelerates tumor formation in humans. Cell 2004, 119, 591–602. [Google Scholar] [CrossRef]
- Iwakuma, T.; Lozano, G. MDM2, an introduction. Mol. Cancer Res. 2003, 1, 993–1000. [Google Scholar] [PubMed]
- Feng, F.Y.; Zhang, Y.; Kothari, V.; Evans, J.R.; Jackson, W.C.; Chen, W.; Johnson, S.B.; Luczak, C.; Wang, S.; Hamstra, D.A. MDM2 inhibition sensitizes prostate cancer cells to androgen ablation and radiotherapy in a p53-dependent manner. Neoplasia 2016, 18, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Malumbres, M.; Barbacid, M. Cell cycle, CDKs and cancer: A changing paradigm. Nat. Rev. Cancer 2009, 9, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; He, H.; Tao, B.-B.; Zhao, Z.-Y.; Hu, G.-H.; Luo, C.; Chen, J.-X.; Ding, X.-H.; Sheng, P.; Dong, Y. Knockdown of CDK6 enhances glioma sensitivity to chemotherapy. Oncol. Rep. 2012, 28, 909–914. [Google Scholar]
- Li, M.; Xiao, A.; Floyd, D.; Olmez, I.; Lee, J.; Godlewski, J.; Bronisz, A.; Bhat, K.P.; Sulman, E.P.; Nakano, I. CDK4/6 inhibition is more active against the glioblastoma proneural subtype. Oncotarget 2017, 8, 55319. [Google Scholar] [CrossRef]
- Chau, Z.; Gulati, N.; Nandu, H.; Nagpal, J.; Zohrabian, V.; Murali, R.; Jhanwar-Uniyal, M. RhoA/Rho kinase influences glioblastoma multiforme cell migration and proliferation via interaction with TGF-β and ERK signaling pathways. Cancer Res. 2007, 67 (Suppl. S9), 5408. [Google Scholar]
- Li, J.; Fan, H.; Zhou, X.; Xiang, Y.; Liu, Y. Prognostic significance and gene co-expression network of PLAU and PLAUR in gliomas. Front. Oncol. 2022, 11, 5689. [Google Scholar] [CrossRef]
- Hsu, D.W.; Efird, J.T.; Hedley-Whyte, E.T. Prognostic role of urokinase-type plasminogen activator in human gliomas. Am. J. Pathol. 1995, 147, 114. [Google Scholar]
- Zhang, X.; Fei, Z.; Bu, X.y.; Zhen, H.n.; Zhang, Z.w.; Gu, J.w.; Chen, Y.j. Expression and significance of urokinase type plasminogen activator gene in human brain gliomas. J. Surg. Oncol. 2000, 74, 90–94. [Google Scholar] [CrossRef]
- Huang, S.-W.; Ali, N.-D.; Zhong, L.; Shi, J. MicroRNAs as biomarkers for human glioblastoma: Progress and potential. Acta Pharmacol. Sin. 2018, 39, 1405–1413. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanif, F.; Zhang, Y.; Dube, C.; Gibert, M.K., Jr.; Saha, S.; Hudson, K.; Marcinkiewicz, P.; Kefas, B.; Guessous, F.; Abounader, R. miR-3174 Is a New Tumor Suppressor MicroRNA That Inhibits Several Tumor-Promoting Genes in Glioblastoma. Int. J. Mol. Sci. 2023, 24, 9326. https://doi.org/10.3390/ijms24119326
Hanif F, Zhang Y, Dube C, Gibert MK Jr., Saha S, Hudson K, Marcinkiewicz P, Kefas B, Guessous F, Abounader R. miR-3174 Is a New Tumor Suppressor MicroRNA That Inhibits Several Tumor-Promoting Genes in Glioblastoma. International Journal of Molecular Sciences. 2023; 24(11):9326. https://doi.org/10.3390/ijms24119326
Chicago/Turabian StyleHanif, Farina, Ying Zhang, Collin Dube, Myron K. Gibert, Jr., Shekhar Saha, Kadie Hudson, Pawel Marcinkiewicz, Benjamin Kefas, Fadila Guessous, and Roger Abounader. 2023. "miR-3174 Is a New Tumor Suppressor MicroRNA That Inhibits Several Tumor-Promoting Genes in Glioblastoma" International Journal of Molecular Sciences 24, no. 11: 9326. https://doi.org/10.3390/ijms24119326