
Immunohistochemistry Reveals TRPC Channels in the Human Hearing Organ—A Novel CT-Guided Approach to the Cochlea
 , , ,
, , ,
Abstract
:1. Introduction


2. Results and Discussion
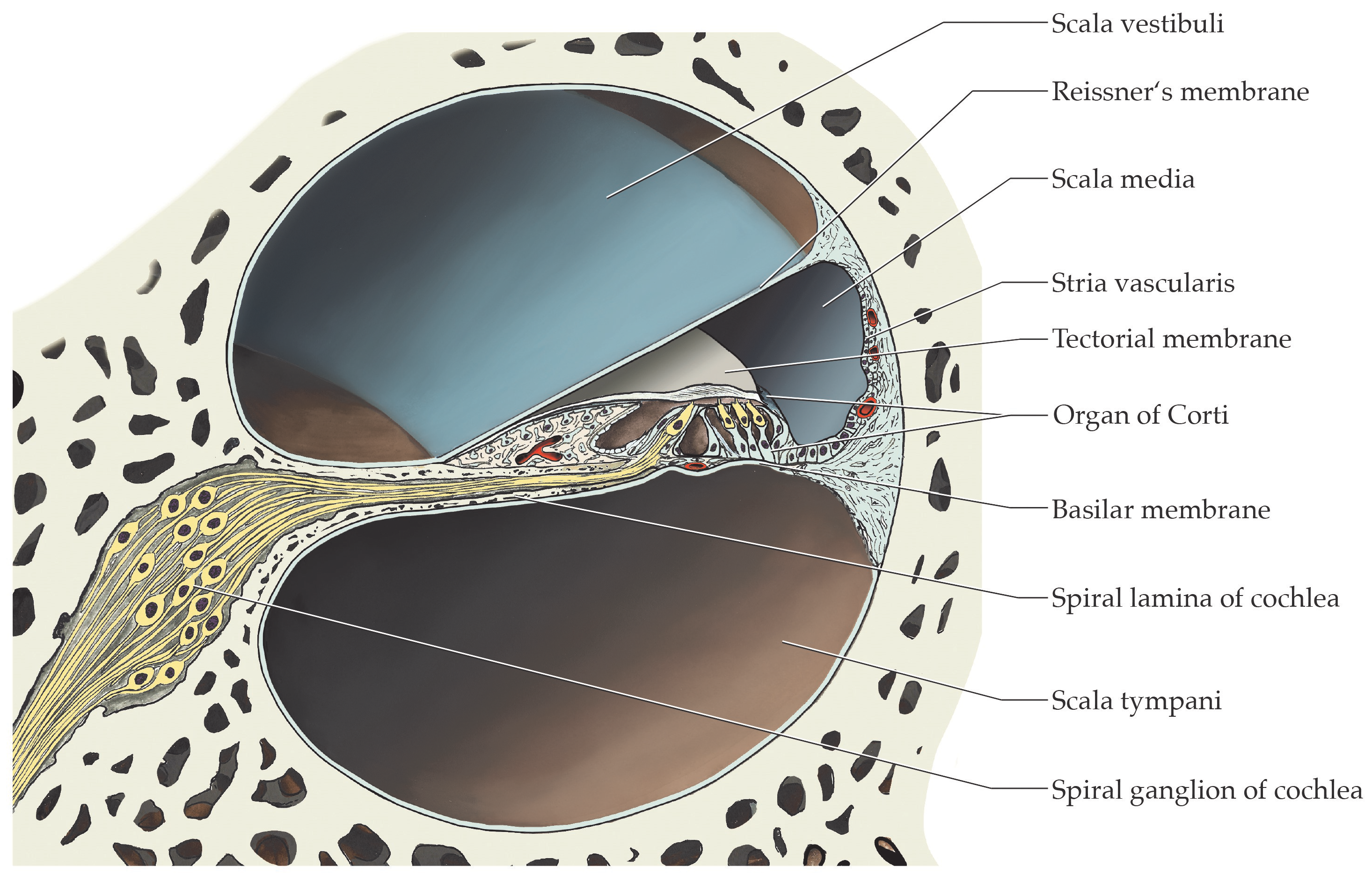
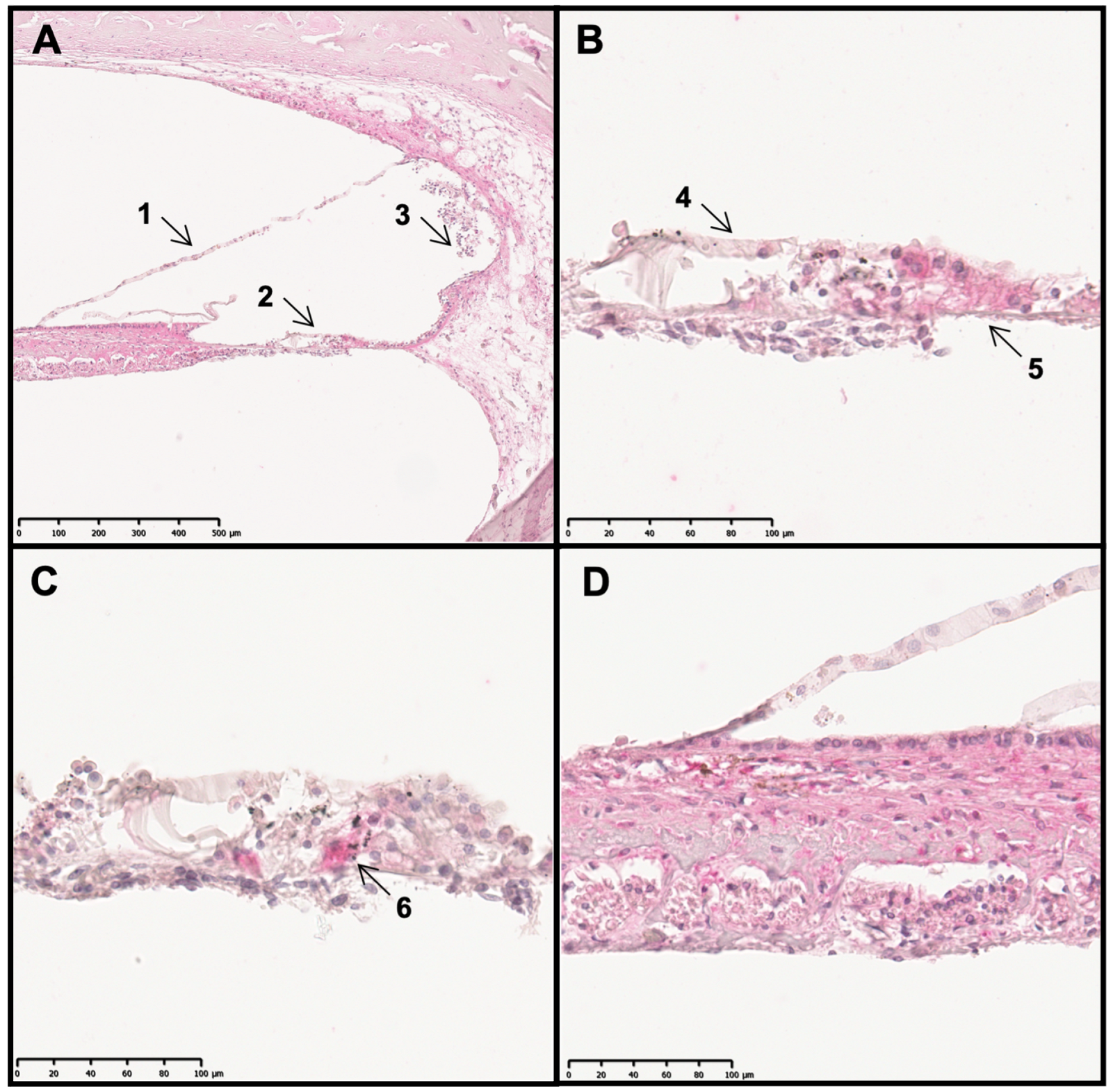
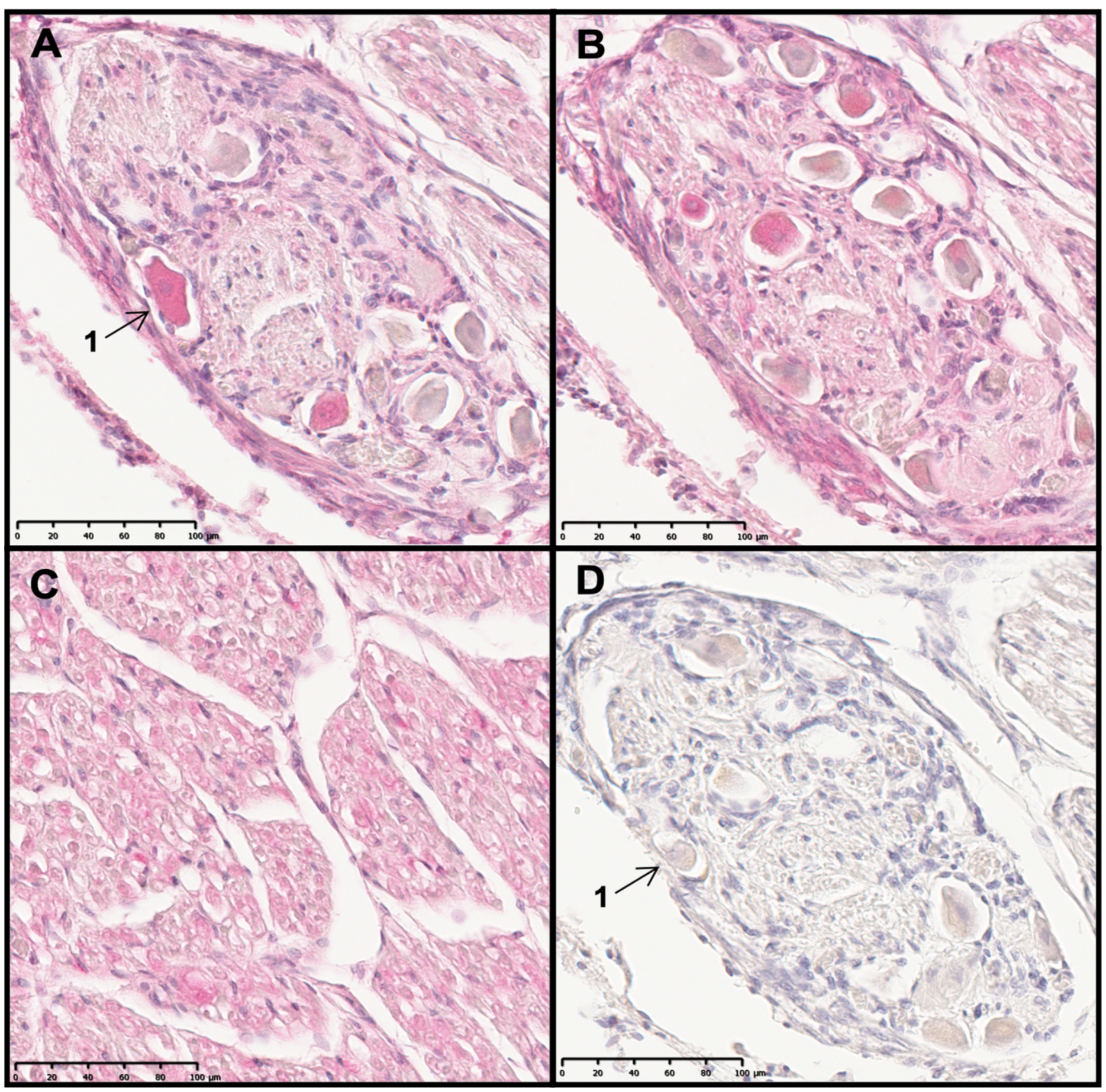
2.1. TRPC6, TRPC5 and TRPC3 Are Revealed in Sections of the Human Cochlea
2.2. The Cochlear Distribution Profile of TRPC Channels Is Similar in Rodents and Humans
2.3. TRPC Channels Are Critical Players in the Hearing Function in Rodents
2.4. TRPC Channels Are Potential Key Structures in Human Cochlear Health and Disease
3. Materials and Methods
3.1. Body Donors: Approval, Sex, Age, Embalming and Preparation
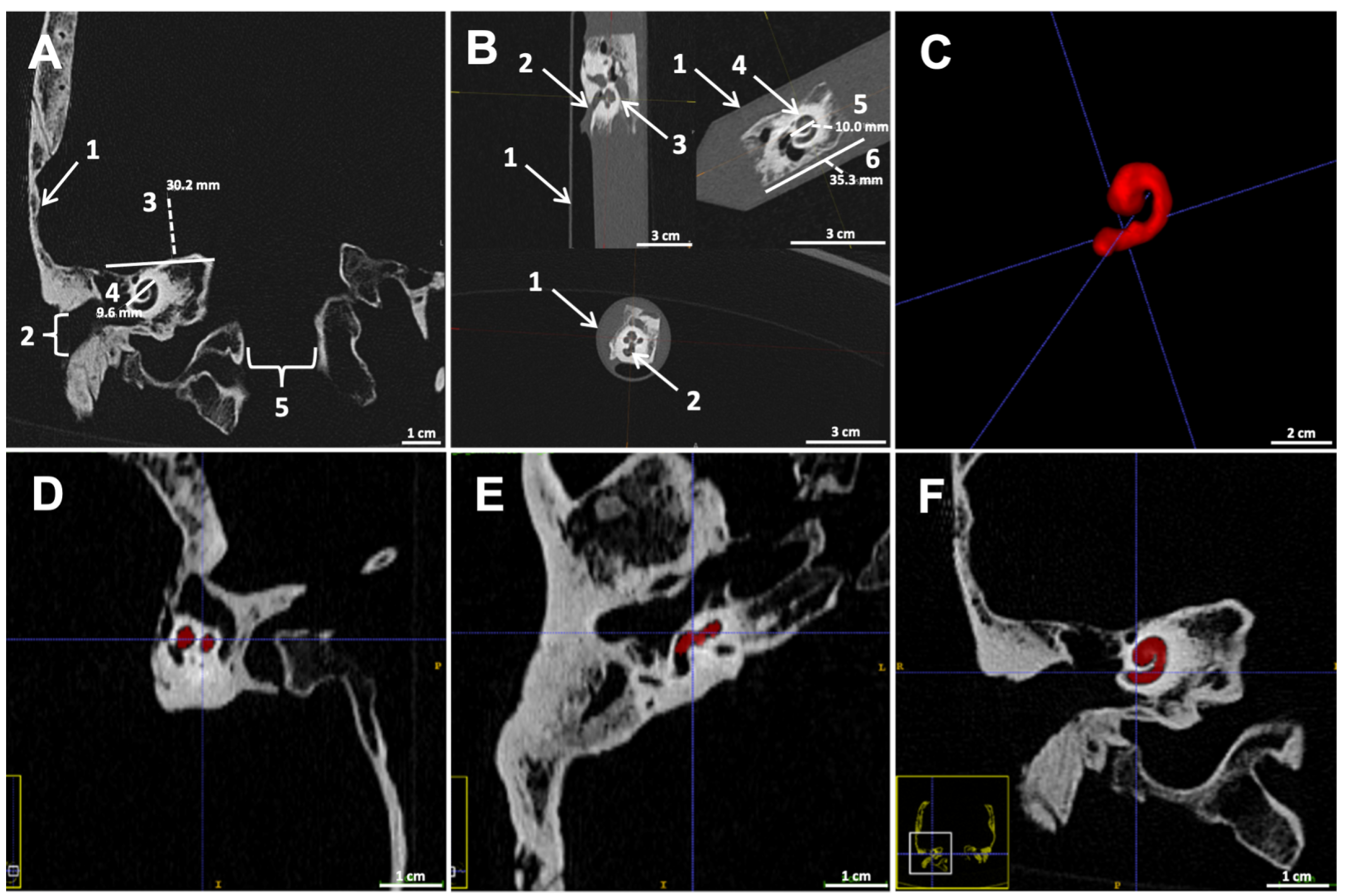
3.2. Imaging of Specimens
3.3. Decalcification Procedure—Challenges and Improvements
3.4. Immunohistochemical Tracing of TRPC6, TRPC5, TRPC3 and Myosin VIIa
3.5. Evaluation and Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morton, N.E. Genetic Epidemiology of Hearing Impairment. Ann. N. Y. Acad. Sci. 1991, 630, 16–31. [Google Scholar] [CrossRef] [PubMed]
- Angeli, S.; Lin, X.; Liu, X.Z. Genetics of Hearing and Deafness. Anat. Rec. 2012, 295, 1812–1829. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L.; Müller, U. Molecular Structure of the Hair Cell Mechanoelectrical Transduction Complex. Cold Spring Harb. Perspect. Med. 2019, 9, a033167. [Google Scholar] [CrossRef]
- Huber, T.B.; Schermer, B.; Müller, R.U.; Höhne, M.; Bartram, M.; Calixto, A.; Hagmann, H.; Reinhardt, C.; Koos, F.; Kunzelmann, K.; et al. Podocin and MEC-2 bind cholesterol to regulate the activity of associated ion channels. Proc. Natl. Acad. Sci. USA 2006, 103, 17079–17086. [Google Scholar] [CrossRef] [PubMed]
- Garrison, S.R.; Dietrich, A.; Stucky, C.L. TRPC1 contributes to light-touch sensation and mechanical responses in low-threshold cutaneous sensory neurons. J. Neurophysiol. 2012, 107, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Quick, K.; Zhao, J.; Eijkelkamp, N.; Linley, J.E.; Rugiero, F.; Cox, J.J.; Raouf, R.; Gringhuis, M.; Sexton, J.E.; Abramowitz, J.; et al. TRPC3 and TRPC6 are essential for normal mechanotransduction in subsets of sensory neurons and cochlear hair cells. Open Biol. 2012, 2, 120068. [Google Scholar] [CrossRef]
- Gao, Y.-Y.; Tian, W.; Zhang, H.-N.; Sun, Y.; Meng, J.-R.; Cao, W.; Li, X.-Q. Canonical transient receptor potential channels and their modulators: Biology, pharmacology and therapeutic potentials. Arch. Pharmacal Res. 2021, 44, 354–377. [Google Scholar] [CrossRef]
- Asghar, M.Y.; Törnquist, K. Transient Receptor Potential Canonical (TRPC) Channels as Modulators of Migration and Invasion. Int. J. Mol. Sci. 2020, 21, 1739. [Google Scholar] [CrossRef]
- Wang, H.; Cheng, X.; Tian, J.; Xiao, Y.; Tian, T.; Xu, F.; Hong, X.; Zhu, M.X. TRPC channels: Structure, function, regulation and recent advances in small molecular probes. Pharmacol. Ther. 2020, 209, 107497. [Google Scholar] [CrossRef]
- Fan, C.; Choi, W.; Sun, W.; Du, J.; Lü, W. Structure of the human lipid-gated cation channel TRPC3. Elife 2018, 7, e36852. [Google Scholar] [CrossRef]
- Englisch, C.N.; Paulsen, F.; Tschernig, T. TRPC Channels in the Physiology and Pathophysiology of the Renal Tubular System: What Do We Know? Int. J. Mol. Sci. 2023, 24, 181. [Google Scholar] [CrossRef] [PubMed]
- Ong, H.L.; de Souza, L.B.; Cheng, K.T.; Ambudkar, I.S. Physiological Functions and Regulation of TRPC Channels. Handb. Exp. Pharm. 2014, 223, 1005–1034. [Google Scholar] [CrossRef]
- Riccio, A.; Medhurst, A.D.; Mattei, C.; Kelsell, R.E.; Calver, A.R.; Randall, A.D.; Benham, C.D.; Pangalos, M.N. mRNA distribution analysis of human TRPC family in CNS and peripheral tissues. Brain Res. Mol. Brain Res. 2002, 109, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Falcón, D.; Galeano-Otero, I.; Martín-Bórnez, M.; Fernández-Velasco, M.; Gallardo-Castillo, I.; Rosado, J.A.; Ordóñez, A.; Smani, T. TRPC Channels: Dysregulation and Ca2+ Mishandling in Ischemic Heart Disease. Cells 2020, 9, 173. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.-C.; Zheng, S.-Y.; Zhang, Z.-G.; Luo, J.-H.; Zhu, Z.-L.; Li, L.; Chen, L.-S.; Lin, X.; Sham, J.S.K.; Lin, M.-J.; et al. TRPC3 promotes tumorigenesis of gastric cancer via the CNB2/GSK3β/NFATc2 signaling pathway. Cancer Lett. 2021, 519, 211–225. [Google Scholar] [CrossRef] [PubMed]
- Fettiplace, R. Hair Cell Transduction, Tuning, and Synaptic Transmission in the Mammalian Cochlea. Compr. Physiol. 2017, 7, 1197–1227. [Google Scholar] [CrossRef]
- Thulasiram, M.R.; Ogier, J.M.; Dabdoub, A. Hearing Function, Degeneration, and Disease: Spotlight on the Stria Vascularis. Front. Cell Dev. Biol. 2022, 10, 841708. [Google Scholar] [CrossRef]
- Klotz-Weigand, L.; Enz, R. Metabotropic Glutamate Receptors at Ribbon Synapses in the Retina and Cochlea. Cells 2022, 11, 1097. [Google Scholar] [CrossRef]
- Reichenbach, T.; Hudspeth, A.J. The physics of hearing: Fluid mechanics and the active process of the inner ear. Rep. Prog. Phys. 2014, 77, 076601. [Google Scholar] [CrossRef]
- Lütjen-Drecoll, E. Funktionnelle Histologie; Volume Hrsg, Version 1.0; Thieme: Stuttgart, Germany, 2021. [Google Scholar]
- Sirko, P.; Kozlov, A.S. Immunohistochemistry localises myosin-7a to cochlear efferent boutons. Wellcome Open Res. 2022, 7, 1. [Google Scholar] [CrossRef]
- Raybould, N.P.; Jagger, D.J.; Kanjhan, R.; Greenwood, D.; Laslo, P.; Hoya, N.; Soeller, C.; Cannell, M.B.; Housley, G.D. TRPC-like conductance mediates restoration of intracellular Ca2+ in cochlear outer hair cells in the guinea pig and rat. J. Physiol. 2007, 579, 101–113. [Google Scholar] [CrossRef]
- Takumida, M.; Anniko, M. Expression of canonical transient receptor potential channel (TRPC) 1–7 in the mouse inner ear. Acta Oto-Laryngologica 2009, 129, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Tadros, S.F.; Kim, Y.; Phan, P.A.; Birnbaumer, L.; Housley, G.D. TRPC3 ion channel subunit immunolocalization in the cochlea. Histochem. Cell Biol. 2010, 133, 137–147. [Google Scholar] [CrossRef]
- Kress, M.; Karasek, J.; Ferrer-Montiel, A.V.; Scherbakov, N.; Haberberger, R.V. TRPC channels and diacylglycerol dependent calcium signaling in rat sensory neurons. Histochem. Cell Biol. 2008, 130, 655–667. [Google Scholar] [CrossRef]
- Tai, Y.; Jia, Y. TRPC Channels and Neuron Development, Plasticity, and Activities. Adv. Exp. Med. Biol. 2017, 976, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.C.; Birnbaumer, L.; Housley, G.D. Canonical transient receptor potential channel subtype 3-mediated hair cell Ca2+ entry regulates sound transduction and auditory neurotransmission. Eur. J. Neurosci. 2013, 37, 1478–1486. [Google Scholar] [CrossRef]
- Zhang, Z.; Tang, J.; Tikunova, S.; Johnson, J.D.; Chen, Z.; Qin, N.; Dietrich, A.; Stefani, E.; Birnbaumer, L.; Zhu, M.X. Activation of Trp3 by inositol 1,4,5-trisphosphate receptors through displacement of inhibitory calmodulin from a common binding domain. Proc. Natl. Acad. Sci. USA 2001, 98, 3168–3173. [Google Scholar] [CrossRef] [PubMed]
- Raybould, N.P.; Housley, G.D. Variation in expression of the outer hair cell P2X receptor conductance along the guinea-pig cochlea. J. Physiol. 1997, 498 Pt 3, 717–727. [Google Scholar] [CrossRef]
- Sexton, J.E.; Desmonds, T.; Quick, K.; Taylor, R.; Abramowitz, J.; Forge, A.; Kros, C.J.; Birnbaumer, L.; Wood, J.N. The contribution of TRPC1, TRPC3, TRPC5 and TRPC6 to touch and hearing. Neurosci. Lett. 2016, 610, 36–42. [Google Scholar] [CrossRef]
- Wu, X.; Indzhykulian, A.A.; Niksch, P.D.; Webber, R.M.; Garcia-Gonzalez, M.; Watnick, T.; Zhou, J.; Vollrath, M.A.; Corey, D.P. Hair-Cell Mechanotransduction Persists in TRP Channel Knockout Mice. PLoS ONE 2016, 11, e0155577. [Google Scholar] [CrossRef]
- Hosoya, M.; Fujioka, M.; Okano, H.; Ogawa, K. Distinct Expression Pattern of a Deafness Gene, KIAA1199, in a Primate Cochlea. BioMed Res. Int. 2016, 2016, 1781894. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, M.; Fujioka, M.; Ogawa, K.; Okano, H. Distinct Expression Patterns of Causative Genes Responsible for Hereditary Progressive Hearing Loss In Non-Human Primate Cochlea. Sci. Rep. 2016, 6, 22250. [Google Scholar] [CrossRef] [PubMed]
- Waschke, J.; Bergmann, M.; Bräuer, L.; Brenner, E.; Buchhorn, A.; Deutsch, A.; Dokter, M.; Egu, D.T.; Ergün, S.; Fassnacht, U.; et al. Recommendations of the working group of the Anatomische Gesellschaft on reduction of formaldehyde exposure in anatomical curricula and institutes. Ann. Anat. 2019, 221, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Yushkevich, P.A.; Piven, J.; Hazlett, H.C.; Smith, R.G.; Ho, S.; Gee, J.C.; Gerig, G. User-guided 3D active contour segmentation of anatomical structures: Significantly improved efficiency and reliability. Neuroimage 2006, 31, 1116–1128. [Google Scholar] [CrossRef] [PubMed]
- Lam, Y.M.; Chen, X.; Pearson, O. Intertaxonomic Variability in Patterns of Bone Density and the Differential Representation of Bovid, Cervid, and Equid Elements in the Archaeological Record. Am. Antiq. 1999, 64, 343–362. [Google Scholar] [CrossRef]
- Gawlitza, J.; Steinhäuser, J.; Bücker, A.; Krasteva-Christ, G.; Tschernig, T. Time is bone—Quantitative comparison of decalcification solvents in human femur samples using dual-X-ray-absorptiometry and computed tomography. Ann. Anat. 2021, 235, 151696. [Google Scholar] [CrossRef]
- Lopez, I.A., 3rd; Ishiyama, G.; Hosokawa, S.; Hosokawa, K.; Acuna, D.; Linthicum, F.H.; Ishiyama, A. Immunohistochemical techniques for the human inner ear. Histochem. Cell Biol. 2016, 146, 367–387. [Google Scholar] [CrossRef]
- Cunningham, C.D., 3rd; Schulte, B.A.; Bianchi, L.M.; Weber, P.C.; Schmiedt, B.N. Microwave Decalcification of Human Temporal Bones. Laryngoscope 2001, 111, 278–282. [Google Scholar] [CrossRef]
- Ghosh, S.; Lewis, M.B.; Walters, B.J. Comparison of ethylenediaminetetraacetic acid and rapid decalcificier solution for studying human temporal bones by immunofluorescence. Laryngoscope Investig. Otolaryngol. 2020, 5, 919–927. [Google Scholar] [CrossRef]
- Englisch, C.N.; Röhricht, D.; Walz, M.; Junker, K.; Beckmann, A.; Meier, C.; Paulsen, F.; Jung, M.; Tschernig, T. TRPC6 Is Found in Distinct Compartments of the Human Kidney. Int. J. Transl. Med. 2022, 2, 156–163. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Sex | Age (Years) | Embalming |
|---|---|---|---|
| 1 | female | 86 | Weigner |
| 2 | female | 84 | Basler |
| 3 | female | 71 | Basler |
| 4 | male | 70 | Weigner |
| 5 | female | 90 | Weigner |
| 6 | female | 93 | Weigner |
| 7 | female | 79 | Basler |
| 8 | female | 89 | Basler |
| 9 | male | 89 | Basler |
| 10 | male | 91 | Basler |
| Number | Total Count Decalcification Solution Replacements | Total Time of Decalcification (Days) |
|---|---|---|
| 1 left (l) | 27× | 93 |
| 1 right (r) | 53× | 270 |
| 2 l/r | 51× | 190 |
| 3 r | 51× | 193 |
| 4 r | 40× | 169 |
| 5 l | 41× | 172 |
| 6 l/r | 30× | 172 |
| 7 l | 23× | 211 |
| 8 l/r | 23× | 211 |
| 9 l | 27× | 253 |
| 10 l/r | 27× | 253 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Englisch, C.N.; Steinhäuser, J.; Wemmert, S.; Jung, M.; Gawlitza, J.; Wenzel, G.; Schick, B.; Tschernig, T. Immunohistochemistry Reveals TRPC Channels in the Human Hearing Organ—A Novel CT-Guided Approach to the Cochlea. Int. J. Mol. Sci. 2023, 24, 9290. https://doi.org/10.3390/ijms24119290
Englisch CN, Steinhäuser J, Wemmert S, Jung M, Gawlitza J, Wenzel G, Schick B, Tschernig T. Immunohistochemistry Reveals TRPC Channels in the Human Hearing Organ—A Novel CT-Guided Approach to the Cochlea. International Journal of Molecular Sciences. 2023; 24(11):9290. https://doi.org/10.3390/ijms24119290
Chicago/Turabian StyleEnglisch, Colya N., Jakob Steinhäuser, Silke Wemmert, Martin Jung, Joshua Gawlitza, Gentiana Wenzel, Bernhard Schick, and Thomas Tschernig. 2023. "Immunohistochemistry Reveals TRPC Channels in the Human Hearing Organ—A Novel CT-Guided Approach to the Cochlea" International Journal of Molecular Sciences 24, no. 11: 9290. https://doi.org/10.3390/ijms24119290