
The ATP Synthase γ Subunit ATPC1 Regulates RNA Editing in Chloroplasts


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Phenotypic and Genetic Characterization of the atpc1 Mutant
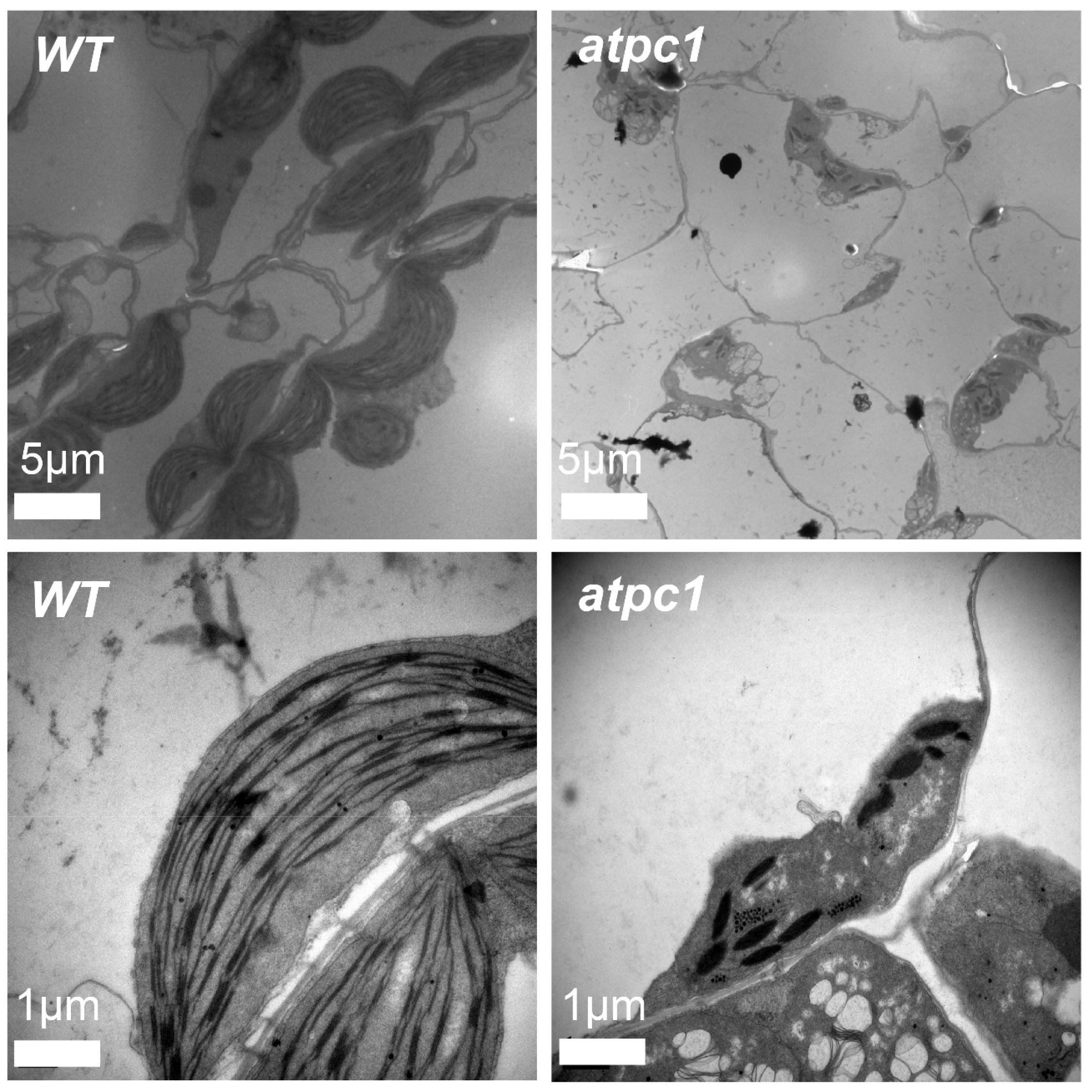
2.2. Disruption of atpC1 Results in Abnormal Chloroplast Development in Arabidopsis
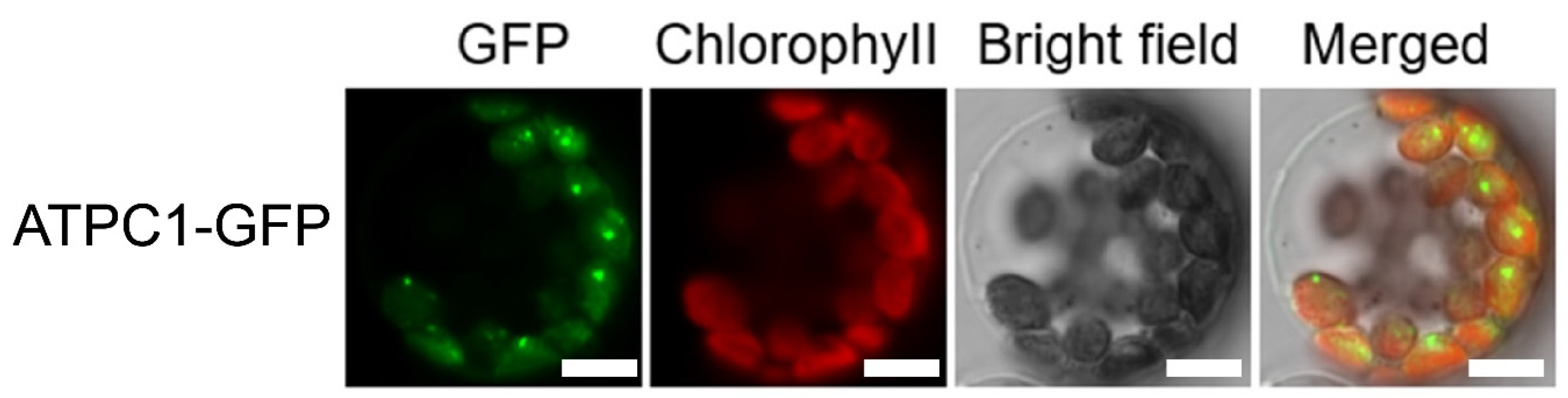
2.3. The ATPC1 Protein Is Localized in Chloroplasts
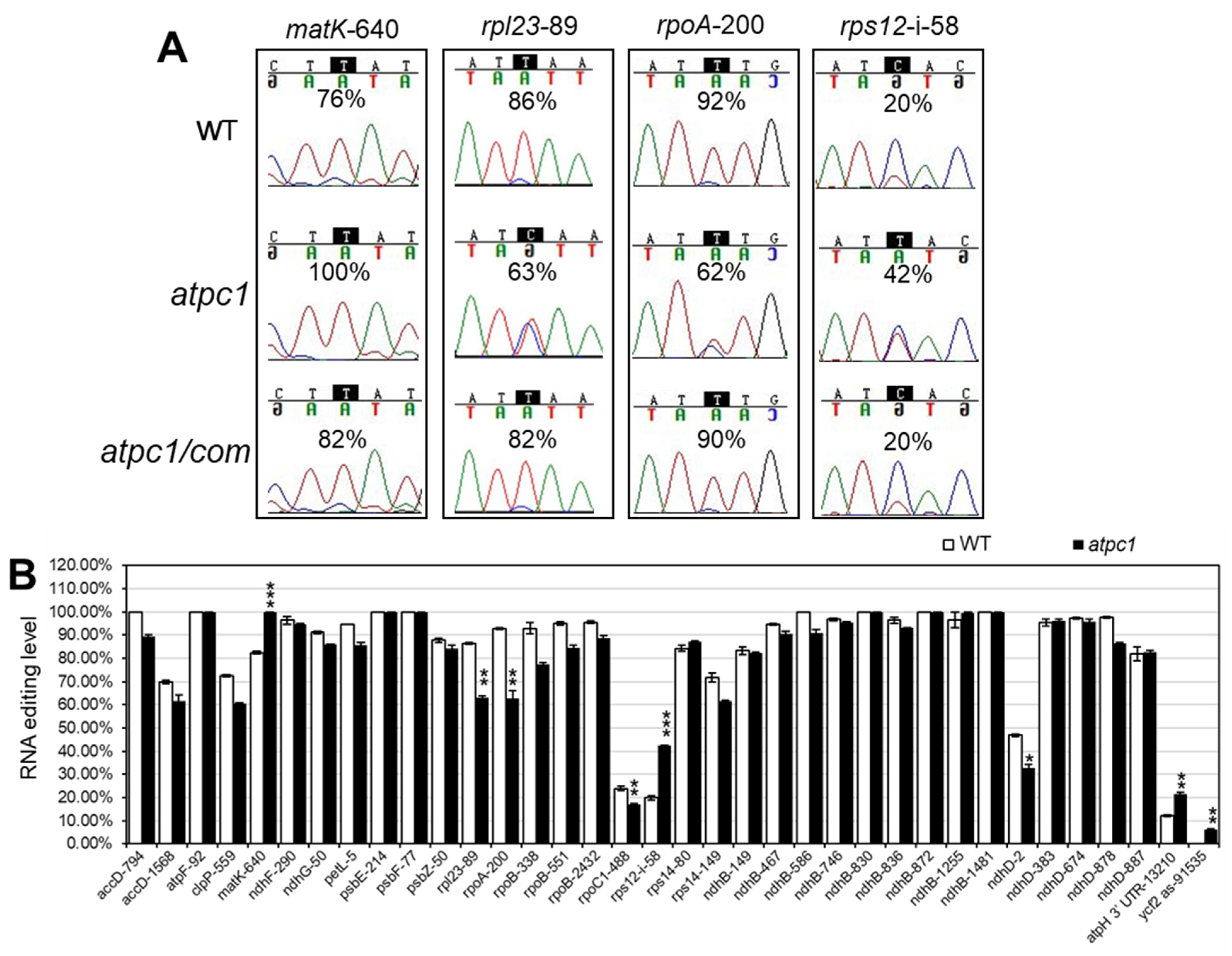
2.4. The Loss of Function of atpC1 Significantly Affects the RNA Editing of Multiple Sites in Chloroplasts
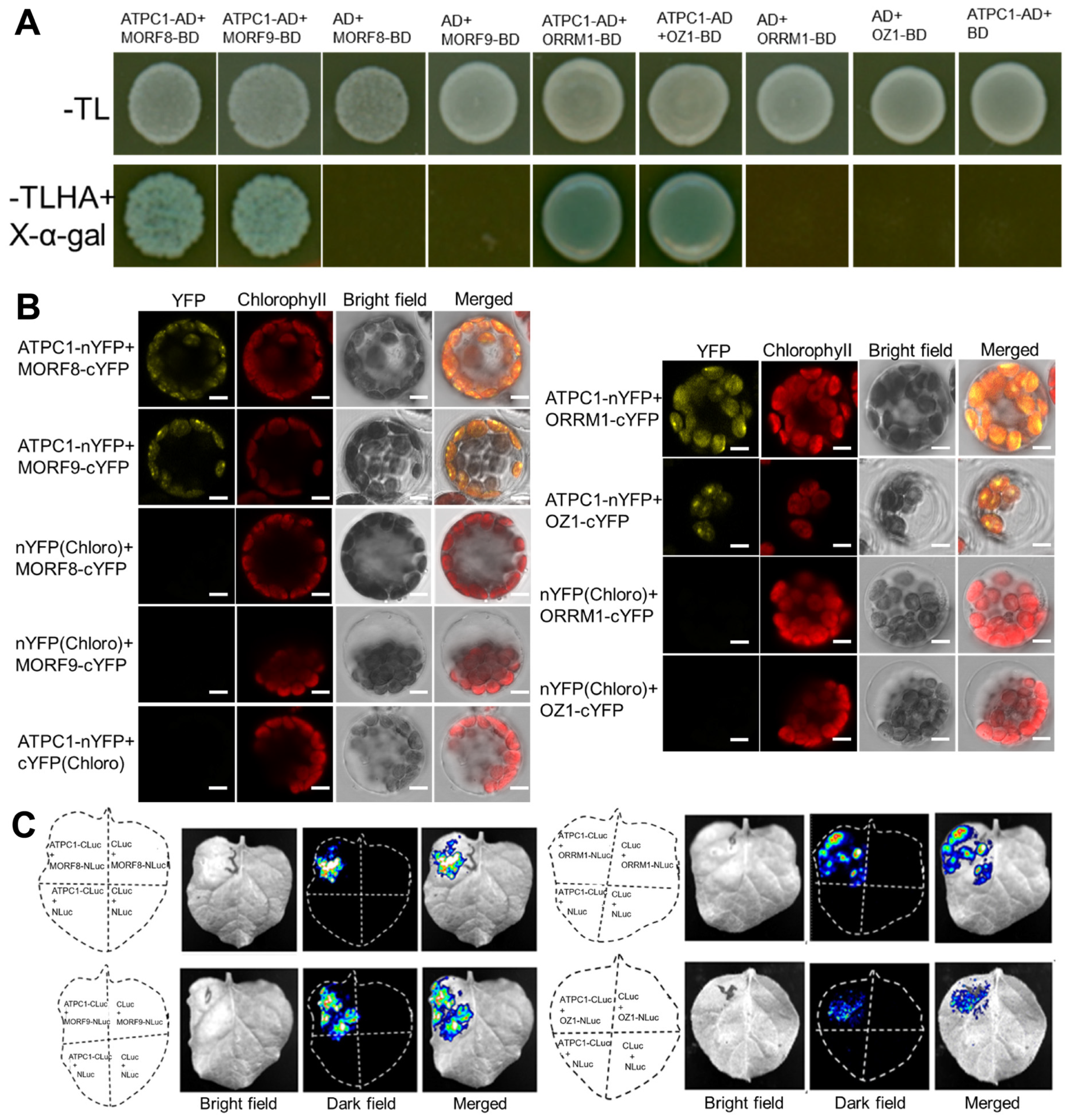
2.5. ATPC1 Interacts with Essential Chloroplast RNA Editosome Component MORF2
2.6. ATPC1 Also Interacts with Other Known Multiple-Site Chloroplast RNA Editing Factors
2.7. Differential Expression of Photosynthesis Genes and Chloroplast Genes in the atpc1 Mutant
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Condition
4.2. Mutant Genotyping
4.3. Transmission Electron Microscopy Analysis
4.4. Plant Transformation and Expression Level Detection
4.5. RNA Isolation and RNA Editing Analysis
4.6. Subcellular Localization
4.7. Yeast Two-Hybrid Assay
4.8. Bimolecular Fluorescence Complementation Assay
4.9. Firefly Luciferase Complementation Imaging (LCI) Assay
4.10. RNA Sequencing and Data Analysis
4.11. Immunoblot Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Takenaka, M.; Zehrmann, A.; Verbitskiy, D.; Härtel, B.; Brennicke, A. RNA Editing in Plants and Its Evolution. Annu. Rev. Genet. 2013, 47, 335–352. [Google Scholar] [CrossRef] [PubMed]
- Barkan, A.; Small, I. Pentatricopeptide Repeat Proteins in Plants. Annu. Rev. Plant. Biol. 2014, 65, 415–442. [Google Scholar] [CrossRef] [PubMed]
- Stern, D.B.; Goldschmidt-Clermont, M.; Hanson, M.R. Chloroplast RNA Metabolism. Annu. Rev. Plant. Biol. 2010, 61, 125–155. [Google Scholar] [CrossRef]
- Sun, T.; Bentolila, S.; Hanson, M.R. The Unexpected Diversity of Plant Organelle RNA Editosomes. Trends Plant. Sci. 2016, 21, 962–973. [Google Scholar] [CrossRef] [PubMed]
- Nishikura, K. Functions and Regulation of RNA Editing by ADAR Deaminases. Annu. Rev. Biochem. 2010, 79, 321–349. [Google Scholar] [CrossRef]
- Takenaka, M.; Zehrmann, A.; Verbitskiy, D.; Kugelmann, M.; Härtel, B.; Brennicke, A. Multiple Organellar RNA Editing Factor (MORF) Family Proteins Are Required for RNA Editing in Mitochondria and Plastids of Plants. Proc. Natl. Acad. Sci. USA 2012, 109, 5104–5109. [Google Scholar] [CrossRef]
- Yan, J.; Zhang, Q.; Yin, P. RNA Editing Machinery in Plant Organelles. Sci. China Life Sci. 2018, 61, 162–169. [Google Scholar] [CrossRef]
- Bentolila, S.; Heller, W.P.; Sun, T.; Babina, A.M.; Friso, G.; van Wijk, K.J.; Hanson, M.R. RIP1, a Member of an Arabidopsis Protein Family, Interacts with the Protein RARE1 and Broadly Affects RNA Editing. Proc. Natl. Acad. Sci. USA 2012, 109, E1453–E1461. [Google Scholar] [CrossRef]
- Ichinose, M.; Sugita, M. RNA Editing and Its Molecular Mechanism in Plant Organelles. Genes 2016, 8, 5. [Google Scholar] [CrossRef]
- Sun, Y.; Law, Y.-S.; Cheng, S.; Lim, B.L. RNA Editing of Cytochrome c Maturation Transcripts Is Responsive to the Energy Status of Leaf Cells in Arabidopsis thaliana . Mitochondrion 2017, 35, 23–34. [Google Scholar] [CrossRef]
- Chateigner-Boutin, A.-L.; Ramos-Vega, M.; Guevara-García, A.; Andrés, C.; De La Luz Gutiérrez-Nava, M.; Cantero, A.; Delannoy, E.; Jiménez, L.F.; Lurin, C.; Small, I.; et al. CLB19, a Pentatricopeptide Repeat Protein Required for Editing of RpoA and ClpP Chloroplast Transcripts. Plant. J. 2008, 56, 590–602. [Google Scholar] [CrossRef]
- Hammani, K.; Okuda, K.; Tanz, S.K.; Chateigner-Boutin, A.-L.; Shikanai, T.; Small, I. A Study of New Arabidopsis Chloroplast RNA Editing Mutants Reveals General Features of Editing Factors and Their Target Sites. Plant. Cell 2009, 21, 3686–3699. [Google Scholar] [CrossRef]
- Okuda, K.; Chateigner-Boutin, A.-L.; Nakamura, T.; Delannoy, E.; Sugita, M.; Myouga, F.; Motohashi, R.; Shinozaki, K.; Small, I.; Shikanai, T. Pentatricopeptide Repeat Proteins with the DYW Motif Have Distinct Molecular Functions in RNA Editing and RNA Cleavage in Arabidopsis Chloroplasts. Plant. Cell 2009, 21, 146–156. [Google Scholar] [CrossRef]
- Cheng, S.; Gutmann, B.; Zhong, X.; Ye, Y.; Fisher, M.F.; Bai, F.; Castleden, I.; Song, Y.; Song, B.; Huang, J.; et al. Redefining the Structural Motifs That Determine RNA Binding and RNA Editing by Pentatricopeptide Repeat Proteins in Land Plants. Plant. J. 2016, 85, 532–547. [Google Scholar] [CrossRef]
- Shi, X.; Bentolila, S.; Hanson, M.R. Organelle RNA Recognition Motif-Containing (ORRM) Proteins Are Plastid and Mitochondrial Editing Factors in Arabidopsis. Plant. Signal. Behav. 2016, 11, e1167299. [Google Scholar] [CrossRef]
- Sun, T.; Germain, A.; Giloteaux, L.; Hammani, K.; Barkan, A.; Hanson, M.R.; Bentolila, S. An RNA Recognition Motif-Containing Protein Is Required for Plastid RNA Editing in Arabidopsis and Maize. Proc. Natl. Acad. Sci. USA 2013, 110, E1169–E1178. [Google Scholar] [CrossRef]
- Sun, T.; Shi, X.; Friso, G.; Van Wijk, K.; Bentolila, S.; Hanson, M.R. A Zinc Finger Motif-Containing Protein Is Essential for Chloroplast RNA Editing. PLoS Genet. 2015, 11, e1005028. [Google Scholar] [CrossRef]
- Zhang, F.; Tang, W.; Hedtke, B.; Zhong, L.; Liu, L.; Peng, L.; Lu, C.; Grimm, B.; Lin, R. Tetrapyrrole Biosynthetic Enzyme Protoporphyrinogen IX Oxidase 1 Is Required for Plastid RNA Editing. Proc. Natl. Acad. Sci. USA 2014, 111, 2023–2028. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Huang, J.; Chory, J. GUN1 Interacts with MORF2 to Regulate Plastid RNA Editing during Retrograde Signaling. Proc. Natl. Acad. Sci. USA 2019, 116, 10162–10167. [Google Scholar] [CrossRef] [PubMed]
- Zehrmann, A.; Härtel, B.; Glass, F.; Bayer-Császár, E.; Obata, T.; Meyer, E.; Brennicke, A.; Takenaka, M. Selective Homo- and Heteromer Interactions between the Multiple Organellar RNA Editing Factor (MORF) Proteins in Arabidopsis thaliana . J. Biol. Chem. 2015, 290, 6445–6456. [Google Scholar] [CrossRef]
- Cui, X.; Wang, Y.; Wu, J.; Han, X.; Gu, X.; Lu, T.; Zhang, Z. The RNA Editing Factor DUA1 Is Crucial to Chloroplast Development at Low Temperature in Rice. New Phytol. 2019, 221, 834–849. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Ren, Y.; Duan, E.; Zhu, X.; Hao, Y.; Zhu, J.; Chen, R.; Lei, J.; Teng, X.; et al. White Panicle2 Encoding Thioredoxin z, Regulates Plastid RNA Editing by Interacting with Multiple Organellar RNA Editing Factors in Rice. New Phytol. 2021, 229, 2693–2706. [Google Scholar] [CrossRef] [PubMed]
- McCarty, R.E.; Evron, Y.; Johnson, E.A. THE CHLOROPLAST ATP SYNTHASE: A Rotary Enzyme? Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 2000, 51, 83–109. [Google Scholar] [CrossRef] [PubMed]
- Hahn, A.; Vonck, J.; Mills, D.J.; Meier, T.; Kühlbrandt, W. Structure, Mechanism, and Regulation of the Chloroplast ATP Synthase. Science 2018, 360, eaat4318. [Google Scholar] [CrossRef]
- Bosco, C.D.; Lezhneva, L.; Biehl, A.; Leister, D.; Strotmann, H.; Wanner, G.; Meurer, J. Inactivation of the Chloroplast ATP Synthase γ Subunit Results in High Non-Photochemical Fluorescence Quenching and Altered Nuclear Gene Expression in Arabidopsis thaliana . J. Biol. Chem. 2004, 279, 1060–1069. [Google Scholar] [CrossRef]
- Inohara, N.; Iwamoto, A.; Moriyama, Y.; Shimomura, S.; Maeda, M.; Futai, M. Two Genes, AtpC1 and AtpC2, for the γ Subunit of Arabidopsis thaliana Chloroplast ATP Synthase. J. Biol. Chem. 1991, 266, 7333–7338. [Google Scholar] [CrossRef]
- Stock, D.; Leslie, A.G.; Walker, J.E. Molecular Architecture of the Rotary Motor in ATP Synthase. Science 1999, 286, 1700–1705. [Google Scholar] [CrossRef]
- Ruwe, H.; Castandet, B.; Schmitz-Linneweber, C.; Stern, D.B. Arabidopsis Chloroplast Quantitative Editotype. FEBS Lett. 2013, 587, 1429–1433. [Google Scholar] [CrossRef]
- Smith, D.R.; Sanitá Lima, M. Unraveling Chloroplast Transcriptomes with ChloroSeq, an Organelle RNA-Seq Bioinformatics Pipeline. Brief Bioinform. 2017, 18, 1012–1016. [Google Scholar] [CrossRef]
- Michel, E.J.S.; Hotto, A.M.; Strickler, S.R.; Stern, D.B.; Castandet, B. A Guide to the Chloroplast Transcriptome Analysis Using RNA-Seq. Methods Mol. Biol. 2018, 1829, 295–313. [Google Scholar] [CrossRef]
- Sánchez-Baracaldo, P.; Cardona, T. On the Origin of Oxygenic Photosynthesis and Cyanobacteria. New Phytol. 2020, 225, 1440–1446. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll Fluorescence: A Probe of Photosynthesis in Vivo. Annu. Rev. Plant. Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef]
- Yan, J.; Zhang, Q.; Guan, Z.; Wang, Q.; Li, L.; Ruan, F.; Lin, R.; Zou, T.; Yin, P. MORF9 Increases the RNA-Binding Activity of PLS-Type Pentatricopeptide Repeat Protein in Plastid RNA Editing. Nat. Plants 2017, 3, 17037. [Google Scholar] [CrossRef]
- Yuan, J.; Ma, T.; Ji, S.; Hedtke, B.; Grimm, B.; Lin, R. Two Chloroplast-Localized MORF Proteins Act as Chaperones to Maintain Tetrapyrrole Biosynthesis. New Phytol. 2022, 235, 1868–1883. [Google Scholar] [CrossRef]
- Alonso, J.M.; Stepanova, A.N.; Leisse, T.J.; Kim, C.J.; Chen, H.; Shinn, P.; Stevenson, D.K.; Zimmerman, J.; Barajas, P.; Cheuk, R.; et al. Genome-Wide Insertional Mutagenesis of Arabidopsis thaliana . Science 2003, 301, 653–657. [Google Scholar] [CrossRef]
- Earley, K.W.; Haag, J.R.; Pontes, O.; Opper, K.; Juehne, T.; Song, K.; Pikaard, C.S. Gateway-Compatible Vectors for Plant Functional Genomics and Proteomics. Plant. J. 2006, 45, 616–629. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral Dip: A Simplified Method for Agrobacterium-Mediated Transformation of Arabidopsis thaliana . Plant. J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Czechowski, T.; Stitt, M.; Altmann, T.; Udvardi, M.K.; Scheible, W.-R. Genome-Wide Identification and Testing of Superior Reference Genes for Transcript Normalization in Arabidopsis. Plant. Physiol. 2005, 139, 5–17. [Google Scholar] [CrossRef]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-Performance Genomics Data Visualization and Exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef]
- Chen, H.; Zou, Y.; Shang, Y.; Lin, H.; Wang, Y.; Cai, R.; Tang, X.; Zhou, J.-M. Firefly Luciferase Complementation Imaging Assay for Protein-Protein Interactions in Plants. Plant. Physiol. 2008, 146, 368–376. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-Level Expression Analysis of RNA-Seq Experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. AgriGO v2.0: A GO Analysis Toolkit for the Agricultural Community, 2017 Update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ni, J.; Song, W.; Ali, N.A.; Zhang, Y.; Xing, J.; Su, K.; Sun, X.; Zhao, X. The ATP Synthase γ Subunit ATPC1 Regulates RNA Editing in Chloroplasts. Int. J. Mol. Sci. 2023, 24, 9203. https://doi.org/10.3390/ijms24119203
Ni J, Song W, Ali NA, Zhang Y, Xing J, Su K, Sun X, Zhao X. The ATP Synthase γ Subunit ATPC1 Regulates RNA Editing in Chloroplasts. International Journal of Molecular Sciences. 2023; 24(11):9203. https://doi.org/10.3390/ijms24119203
Chicago/Turabian StyleNi, Jia, Wenjian Song, Nadia Ahmed Ali, Yayi Zhang, Jiani Xing, Kexing Su, Xingxing Sun, and Xiaobo Zhao. 2023. "The ATP Synthase γ Subunit ATPC1 Regulates RNA Editing in Chloroplasts" International Journal of Molecular Sciences 24, no. 11: 9203. https://doi.org/10.3390/ijms24119203