
Upregulation of the Oct3/4 Network in Basal Breast Cancer Is Associated with Its Metastatic Potential and Shows Tissue Dependent Variability
 , ,
, , 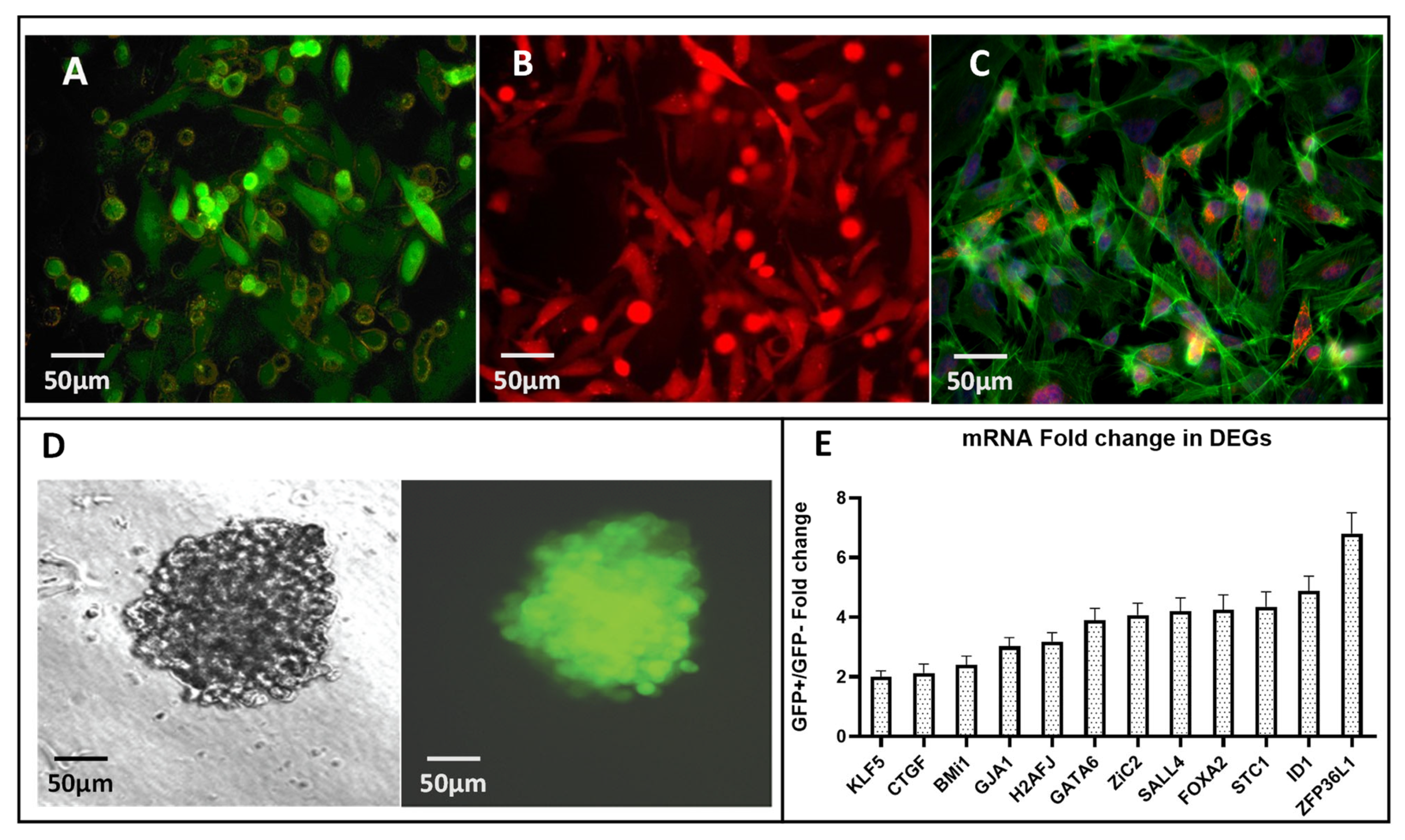{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Oct3/4-Transfected MDA-MB-231 Cells Demonstrate Positive Correlation with Oct3/4 Expression and Cancer Stem Cell Phenotype with an Upregulated Transcriptome of Pioneer Factors
2.2. Oct3/4 Expression Is Associated with High Tumorigenic Potential of MDA-MB-231 Cells
2.3. Metastatic Potential of MDA-MB-231 Tumor Cells Is Associated with an Upregulated Oct3/4 Network Specific for Cancer Stem Cell Phenotype and Tumor Growth
2.4. Oct3/4-Transfected MDA-MB-231 Tumors in Mice Demonstrate Organ-Specific Heterogeneity in Upregulation of Oct3/4 Transcriptome
2.5. MDA-MB-231 with Activated Oct3/4 Cells Evade Paclitaxel-Induced Cell Death and Generate Tumors with Significantly Upregulated Stemness Genotype (CD44+/CD24-)
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Generation of Oct3/4-eGFP Transgenic MDA-MB-231 Cell Sublines and Clones by Stable Transfection
4.3. Flow Cytometry-Based Cell Sorting (FACS) and Establishing Monoclonal Colonies
4.4. RNA Isolation and RT-PCR Analyses of Gene Expression
4.5. Monoclonal Oncosphere Culture System Stem Cell Culture (SCC) Medium
4.6. Cell Proliferation Assay in the Presence of Paclitaxel
4.7. Generating Orthotopic Primary Tumors and Spontaneous Lung Metastases Using BALB/C Mouse Xenograft Model
4.7.1. Primary Tumor Growth Model
4.7.2. Lung Metastasis Model
4.7.3. Flow Cytometry and Single Cell Sorting of CD44+/CD24-Populations
4.7.4. Flow Cytometry Staining
4.7.5. Microarray Gene Expression Analysis
4.7.6. mRNA Synthesis and Labelling
4.7.7. Oligonucleotide Array Hybridization and Analysis
4.7.8. Validation of Affimetrix Gene Chip Results by Using Real Time Semi-Quantitative Reverse Transcriptase PCR
4.7.9. Quantitative Real-Time Reverse Transcription PCR Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eng, L.G.; Dawood, S.; Sopik, V.; Haaland, B.; Tan, P.S.; Bhoo-Pathy, N.; Warner, E.; Iqbal, J.; Narod, S.A.; Dent, R. Ten-year survival in women with primary stage IV breast cancer. Breast Cancer Res. Treat. 2016, 160, 145–152. [Google Scholar] [CrossRef]
- Jemel, I.; Jellali, K.; Elloumi, J.; Aifa, S. The offer of chemistry to targeted therapy in cancer. Recent Pat. Biotechnol. 2011, 5, 174–182. [Google Scholar] [CrossRef]
- Marusyk, A.; Janiszewska, M.; Polyak, K. Intratumor Heterogeneity: The Rosetta Stone of Therapy Resistance. Cancer Cell 2020, 37, 471–484. [Google Scholar] [CrossRef]
- Huang, T.; Song, X.; Xu, D.; Tiek, D.; Goenka, A.; Wu, B.; Sastry, N.; Hu, B.; Cheng, S.Y. Stem cell programs in cancer initiation, progression, and therapy resistance. Theranostics 2020, 10, 8721–8743. [Google Scholar] [CrossRef]
- Yang, J.; Chai, L.; Fowles, T.C.; Alipio, Z.; Xu, D.; Fink, L.M.; Ward, D.C.; Ma, Y. Genome-wide analysis reveals Sall4 to be a major regulator of pluripotency in murine-embryonic stem cells. Proc. Natl. Acad. Sci. USA 2008, 105, 19756–19761. [Google Scholar] [CrossRef]
- Abboud, N.; Morris, T.M.; Hiriart, E.; Yang, H.; Bezerra, H.; Gualazzi, M.G.; Stefanovic, S.; Guénantin, A.C.; Evans, S.M.; Pucéat, M. A cohesin-OCT4 complex mediates Sox enhancers to prime an early embryonic lineage. Nat. Commun. 2015, 6, 6749. [Google Scholar] [CrossRef]
- Robinson, E.L.; Anene-Nzelu, C.G.; Rosa-Garrido, M.; Foo, R.S.Y. Cardiac epigenetics: Driving signals to the cardiac epigenome in development and disease. J. Mol. Cell. Cardiol. 2021, 151, 88. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Walsh, L.A.; Alvarez, M.J.; Sabio, E.Y.; Reyngold, M.; Makarov, V.; Mukherjee, S.; Lee, K.W.; Desrichard, A.; Turcan, Ş.; Dalin, M.G.; et al. An Integrated Systems Biology Approach Identifies TRIM25 as a Key Determinant of Breast Cancer Metastasis. Cell Rep. 2017, 20, 1623–1640. [Google Scholar] [CrossRef]
- Liu, M.C.; Oxnard, G.R.; Klein, E.A.; Swanton, C.; Seiden, M.V.; Consortium, C. Sensitive and specific multi-cancer detection and localization using methylation signatures in cell-free DNA. Ann. Oncol. 2020, 31, 745–759. [Google Scholar] [CrossRef]
- Himanen, S.V.; Sistonen, L. New insights into transcriptional reprogramming during cellular stress. J. Cell Sci. 2019, 132, jcs238402. [Google Scholar] [CrossRef]
- Prager, B.C.; Bhargava, S.; Mahadev, V.; Hubert, C.G.; Rich, J.N. Glioblastoma Stem Cells: Driving Resilience through Chaos. Trends Cancer 2020, 6, 223–235. [Google Scholar] [CrossRef]
- Ramos, E.K.; Hoffmann, A.D.; Gerson, S.L.; Liu, H. New Opportunities and Challenges to Defeat Cancer Stem Cells. Trends Cancer 2017, 3, 780–796. [Google Scholar] [CrossRef]
- Dirkse, A.; Golebiewska, A.; Buder, T.; Nazarov, P.V.; Muller, A.; Poovathingal, S.; Brons, N.H.C.; Leite, S.; Sauvageot, N.; Sarkisjan, D.; et al. Stem cell-associated heterogeneity in Glioblastoma results from intrinsic tumor plasticity shaped by the microenvironment. Nat. Commun. 2019, 10, 1787. [Google Scholar] [CrossRef]
- Gerrard, L.; Zhao, D.; Clark, A.J.; Cui, W. Stably transfected human embryonic stem cell clones express OCT4-specific green fluorescent protein and maintain self-renewal and pluripotency. Stem Cells 2005, 23, 124–133. [Google Scholar] [CrossRef]
- Gibbs, C.P.; Kukekov, V.G.; Reith, J.D.; Tchigrinova, O.; Suslov, O.N.; Scott, E.W.; Ghivizzani, S.C.; Ignatova, T.N.; Steindler, D.A. Stem-like cells in bone sarcomas: Implications for tumorigenesis. Neoplasia 2005, 7, 967–976. [Google Scholar] [CrossRef]
- Ignatova, T.N.; Chaitin, H.J.; Kukekov, N.V.; Suslov, O.N.; Dulatova, G.I.; Hanafy, K.A.; Vrionis, F.D. Gliomagenesis is orchestrated by the Oct3/4 regulatory network. J. Neurosurg. Sci. 2021. [Google Scholar] [CrossRef]
- Basati, G.; Mohammadpour, H.; Emami Razavi, A. Association of High Expression Levels of SOX2, NANOG, and OCT4 in Gastric Cancer Tumor Tissues with Progression and Poor Prognosis. J. Gastrointest Cancer 2020, 51, 41–47. [Google Scholar] [CrossRef]
- Clemente-Periván, S.I.; Gómez-Gómez, Y.; Leyva-Vázquez, M.A.; Lagunas-Martínez, A.; Organista-Nava, J.; Illades-Aguiar, B. Role of Oct3/4 in Cervical Cancer Tumorigenesis. Front. Oncol. 2020, 10, 247. [Google Scholar] [CrossRef]
- Ignatova, T.N.; Kukekov, V.G.; Laywell, E.D.; Suslov, O.N.; Vrionis, F.D.; Steindler, D.A. Human cortical glial tumors contain neural stem-like cells expressing astroglial and neuronal markers in vitro. Glia 2002, 39, 193–206. [Google Scholar] [CrossRef]
- Boyer, L.A.; Lee, T.I.; Cole, M.F.; Johnstone, S.E.; Levine, S.S.; Zucker, J.P.; Guenther, M.G.; Kumar, R.M.; Murray, H.L.; Jenner, R.G.; et al. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell 2005, 122, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 3983–3988. [Google Scholar] [CrossRef] [PubMed]
- Zasadil, L.M.; Andersen, K.A.; Yeum, D.; Rocque, G.B.; Wilke, L.G.; Tevaarwerk, A.J.; Raines, R.T.; Burkard, M.E.; Weaver, B.A. Cytotoxicity of paclitaxel in breast cancer is due to chromosome missegregation on multipolar spindles. Sci. Transl. Med. 2014, 6, 229ra243. [Google Scholar] [CrossRef] [PubMed]
- Hassn Mesrati, M.; Syafruddin, S.E.; Mohtar, M.A.; Syahir, A. CD44: A Multifunctional Mediator of Cancer Progression. Biomolecules 2021, 11, 1850. [Google Scholar] [CrossRef] [PubMed]
- Castelli, V.; Giordano, A.; Benedetti, E.; Giansanti, F.; Quintiliani, M.; Cimini, A.; d’Angelo, M. The Great Escape: The Power of Cancer Stem Cells to Evade Programmed Cell Death. Cancers 2021, 13, 328. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.Q.; Misra, Y.; Li, D.B.; Wadsworth, M.P.; Krag, D.; Weaver, D.; Tessitore, J.; Li, D.W.; Zhang, G.; Tian, Q.; et al. Differential expression of Oct3/4 in human breast cancer and normal tissues. Int. J. Oncol. 2018, 52, 2069–2078. [Google Scholar] [CrossRef]
- Chen, M.-T.; Sun, H.-F.; Zhao, Y.; Fu, W.-Y.; Yang, L.-P.; Gao, S.-P.; Li, L.-D.; Jiang, H.-l.; Jin, W. Comparison of patterns and prognosis among distant metastatic breast cancer patients by age groups: A SEER population-based analysis. Sci. Rep. 2017, 7, 9254. [Google Scholar] [CrossRef]
- Gidekel, S.; Pizov, G.; Bergman, Y.; Pikarsky, E. Oct-3/4 is a dose-dependent oncogenic fate determinant. Cancer Cell 2003, 4, 361–370. [Google Scholar] [CrossRef]
- Kadauke, S.; Blobel, G.A. Mitotic bookmarking by transcription factors. Epigenet. Chromatin 2013, 6, 6. [Google Scholar] [CrossRef]
- Soujanya, M.; Bihani, A.; Hajirnis, N.; Pathak, R.U.; Mishra, R.K. Nuclear architecture and the structural basis of mitotic memory. Chromosome Res. 2023, 31, 8. [Google Scholar] [CrossRef]
- Wang, Y.J.; Herlyn, M. The emerging roles of Oct4 in tumor-initiating cells. Am. J. Physiol. Cell. Physiol. 2015, 309, C709–C718. [Google Scholar] [CrossRef] [PubMed]
- Anatskaya, O.V.; Vinogradov, A.E. Polyploidy and Myc Proto-Oncogenes Promote Stress Adaptation via Epigenetic Plasticity and Gene Regulatory Network Rewiring. Int. J. Mol. Sci. 2022, 23, 9691. [Google Scholar] [CrossRef]
- Chew, J.L.; Loh, Y.H.; Zhang, W.; Chen, X.; Tam, W.L.; Yeap, L.S.; Li, P.; Ang, Y.S.; Lim, B.; Robson, P.; et al. Reciprocal transcriptional regulation of Pou5f1 and Sox2 via the Oct4/Sox2 complex in embryonic stem cells. Mol. Cell. Biol. 2005, 25, 6031–6046. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Jiang, P.; Chen, J.; Li, J.; Li, D.; He, Y.; Jiang, Y.; Zhang, Y.; Xu, S.; Li, X.; et al. GATA6 promotes epithelial-mesenchymal transition and metastasis through MUC1/β-catenin pathway in cholangiocarcinoma. Cell Death Dis. 2020, 11, 860. [Google Scholar] [CrossRef] [PubMed]
- Bryzgalov, L.O.; Ershov, N.I.; Oshchepkov, D.Y.; Kaledin, V.I.; Merkulova, T.I. Detection of target genes of FOXA transcription factors involved in proliferation control. Biochemistry 2008, 73, 70–75. [Google Scholar] [CrossRef]
- Kagey, M.H.; He, X. Rationale for targeting the Wnt signalling modulator Dickkopf-1 for oncology. Br. J. Pharmacol. 2017, 174, 4637–4650. [Google Scholar] [CrossRef]
- Boyle, M.J.; Seaver, E.C. Developmental expression of foxA and gata genes during gut formation in the polychaete annelid, Capitella sp. I. Evol. Dev. 2008, 10, 89–105. [Google Scholar] [CrossRef]
- Guo, Y.; Costa, R.; Ramsey, H.; Starnes, T.; Vance, G.; Robertson, K.; Kelley, M.; Reinbold, R.; Scholer, H.; Hromas, R. The embryonic stem cell transcription factors Oct-4 and FoxD3 interact to regulate endodermal-specific promoter expression. Proc. Natl. Acad. Sci. USA 2002, 99, 3663–3667. [Google Scholar] [CrossRef]
- van der Sluis, M.; Vincent, A.; Bouma, J.; Korteland-Van Male, A.; van Goudoever, J.B.; Renes, I.B.; Van Seuningen, I. Forkhead box transcription factors Foxa1 and Foxa2 are important regulators of Muc2 mucin expression in intestinal epithelial cells. Biochem. Biophys. Res. Commun. 2008, 369, 1108–1113. [Google Scholar] [CrossRef]
- Chen, T.; Tsang, J.Y.S.; Su, X.C.; Li, P.; Sun, W.Q.; Wong, I.L.K.; Choy, K.Y.; Yang, Q.; Tse, G.M.K.; Chan, T.H.; et al. SALL4 promotes tumor progression in breast cancer by targeting EMT. Mol. Carcinog. 2020, 59, 1209–1226. [Google Scholar] [CrossRef]
- Lim, C.Y.; Tam, W.L.; Zhang, J.; Ang, H.S.; Jia, H.; Lipovich, L.; Ng, H.H.; Wei, C.L.; Sung, W.K.; Robson, P.; et al. Sall4 regulates distinct transcription circuitries in different blastocyst-derived stem cell lineages. Cell Stem Cell 2008, 3, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Nicolè, L.; Sanavia, T.; Veronese, N.; Cappellesso, R.; Luchini, C.; Dabrilli, P.; Fassina, A. Oncofetal gene SALL4 and prognosis in cancer: A systematic review with meta-analysis. Oncotarget 2017, 8, 22968–22979. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Jeong, H.W.; Kong, N.; Yang, Y.; Carroll, J.; Luo, H.R.; Silberstein, L.E.; Chai, Y.L. Stem cell factor SALL4 represses the transcriptions of PTEN and SALL1 through an epigenetic repressor complex. PLoS ONE 2009, 4, e5577. [Google Scholar] [CrossRef]
- Yang, J.; Corsello, T.R.; Ma, Y. Stem cell gene SALL4 suppresses transcription through recruitment of DNA methyltransferases. J. Biol. Chem. 2012, 287, 1996–2005. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, J.; Yang, X.; Fang, C.; Xu, H.; Xi, X. SALL4 as an Epithelial-Mesenchymal Transition and Drug Resistance Inducer through the Regulation of c-Myc in Endometrial Cancer. PLoS ONE 2015, 10, e0138515. [Google Scholar] [CrossRef] [PubMed]
- Erenpreisa, J.; Salmina, K.; Anatskaya, O.; Cragg, M.S. Paradoxes of cancer: Survival at the brink. Semin. Cancer Biol. 2022, 81, 119–131. [Google Scholar] [CrossRef]
- Altevogt, P.; Sammar, M.; Hüser, L.; Kristiansen, G. Novel insights into the function of CD24: A driving force in cancer. Int. J. Cancer 2021, 148, 546–559. [Google Scholar] [CrossRef]
- Irizarry, R.A.; Hobbs, B.; Collin, F.; Beazer-Barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics 2003, 4, 249–264. [Google Scholar] [CrossRef]
- Niwa, H.; Miyazaki, J.-i.; Smith, A. Quantitative expression of Oct-3/4 defines differentiation, dedifferentiation, or self-renewal of ES cells. Nat. Genet. 2000, 24, 372–376. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajan, R.G.; Krutilina, R.I.; Ignatova, T.N.; Pavicevich, Z.S.; Dulatova, G.M.; Lane, M.A.; Chatterjee, A.R.; Rooney, R.J.; Antony, M.; Hagerty, V.R.; et al. Upregulation of the Oct3/4 Network in Basal Breast Cancer Is Associated with Its Metastatic Potential and Shows Tissue Dependent Variability. Int. J. Mol. Sci. 2023, 24, 9142. https://doi.org/10.3390/ijms24119142
Rajan RG, Krutilina RI, Ignatova TN, Pavicevich ZS, Dulatova GM, Lane MA, Chatterjee AR, Rooney RJ, Antony M, Hagerty VR, et al. Upregulation of the Oct3/4 Network in Basal Breast Cancer Is Associated with Its Metastatic Potential and Shows Tissue Dependent Variability. International Journal of Molecular Sciences. 2023; 24(11):9142. https://doi.org/10.3390/ijms24119142
Chicago/Turabian StyleRajan, Robin G., Raisa I. Krutilina, Tatyana N. Ignatova, Zoran S. Pavicevich, Galina M. Dulatova, Maria A. Lane, Arindam R. Chatterjee, Robert J. Rooney, Mymoon Antony, Vivian R. Hagerty, and et al. 2023. "Upregulation of the Oct3/4 Network in Basal Breast Cancer Is Associated with Its Metastatic Potential and Shows Tissue Dependent Variability" International Journal of Molecular Sciences 24, no. 11: 9142. https://doi.org/10.3390/ijms24119142