
Genetic Diversity of a Heat Activated Channel—TRPV1 in Two Desert Gerbil Species with Different Heat Sensitivity
 , and
, and 
Abstract
:1. Introduction
2. Results
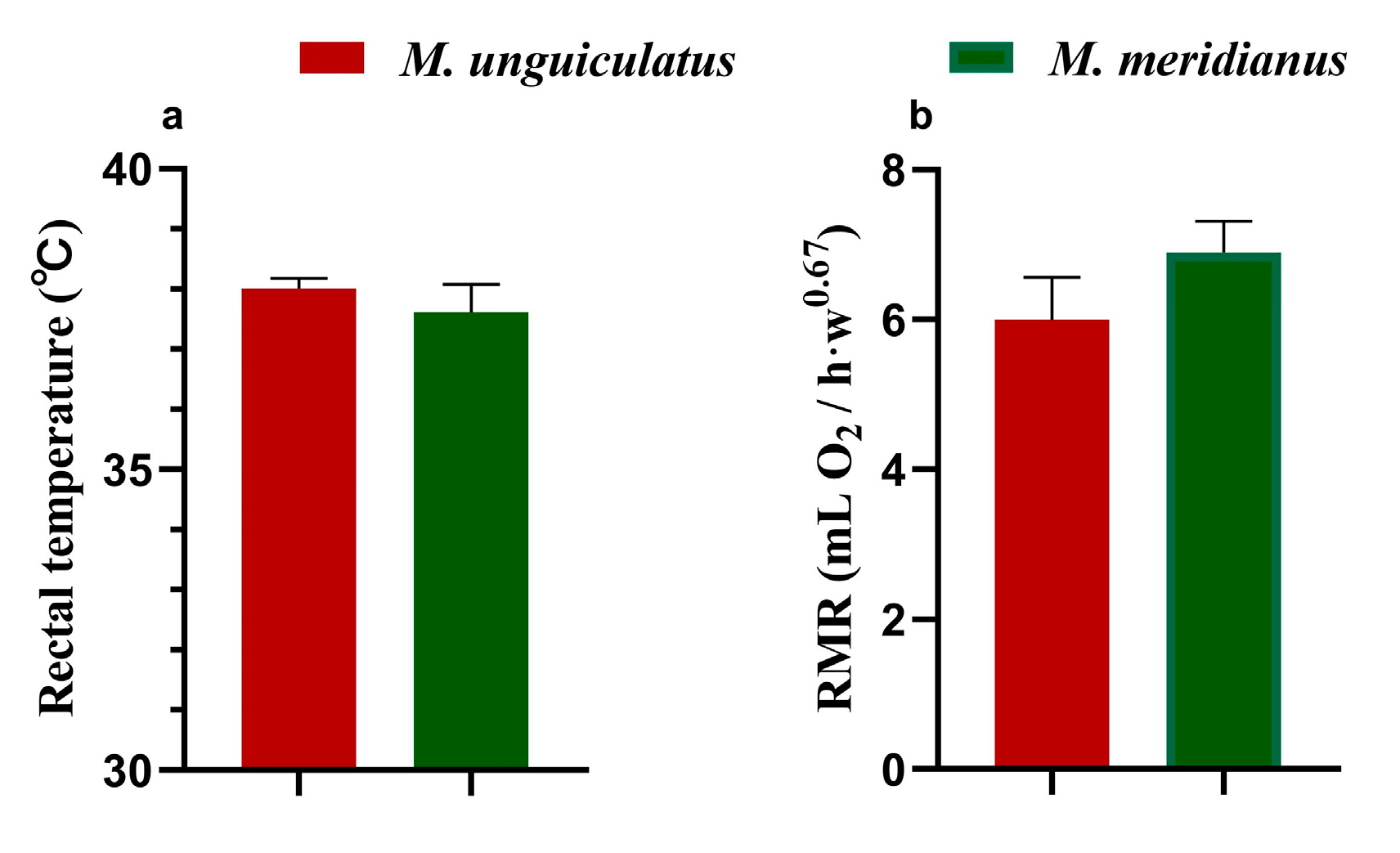
2.1. Morphological and Physiological Traits of M. unguiculatus and M. meridianus
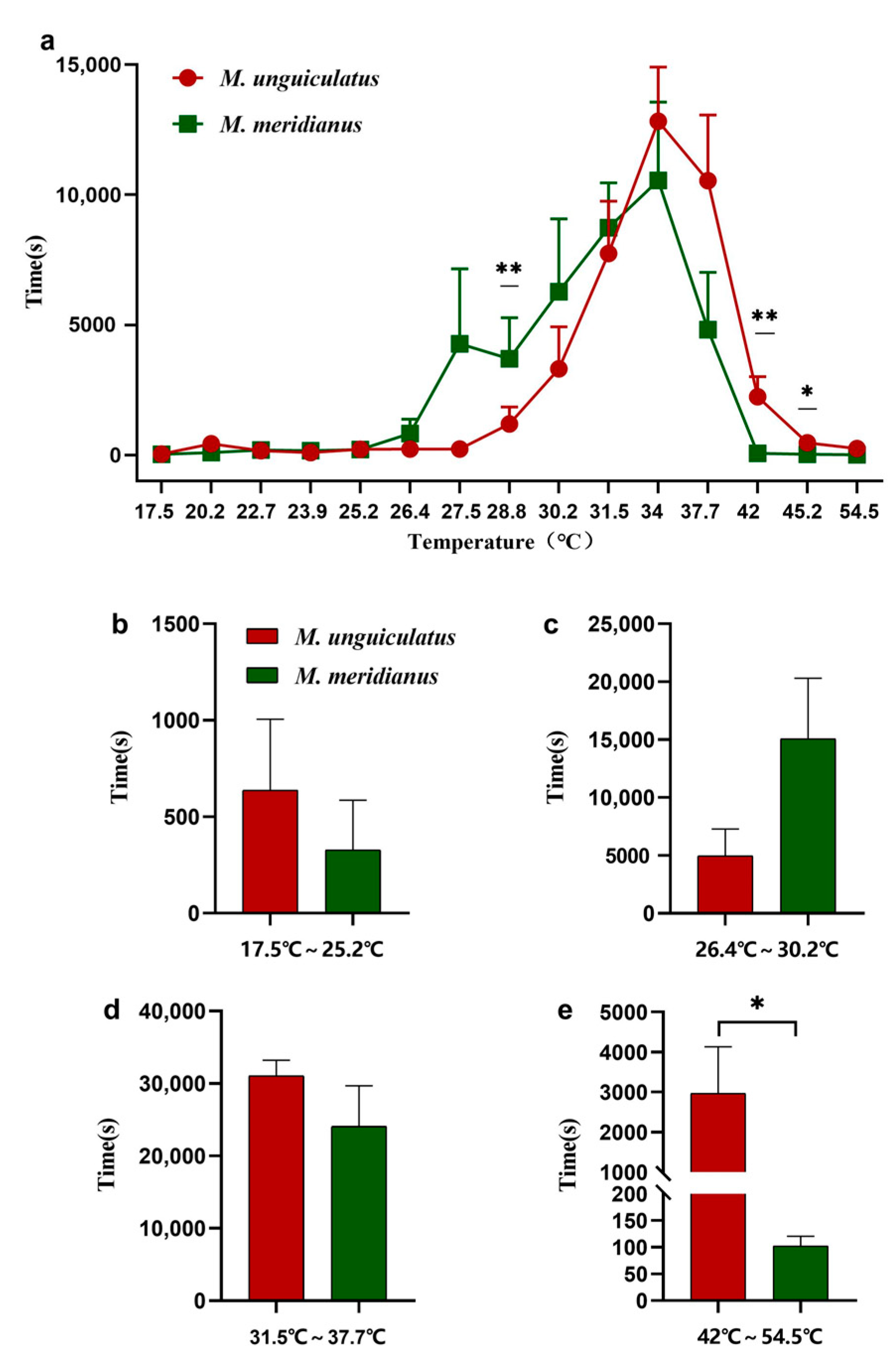
2.2. Temperature Preference Trials
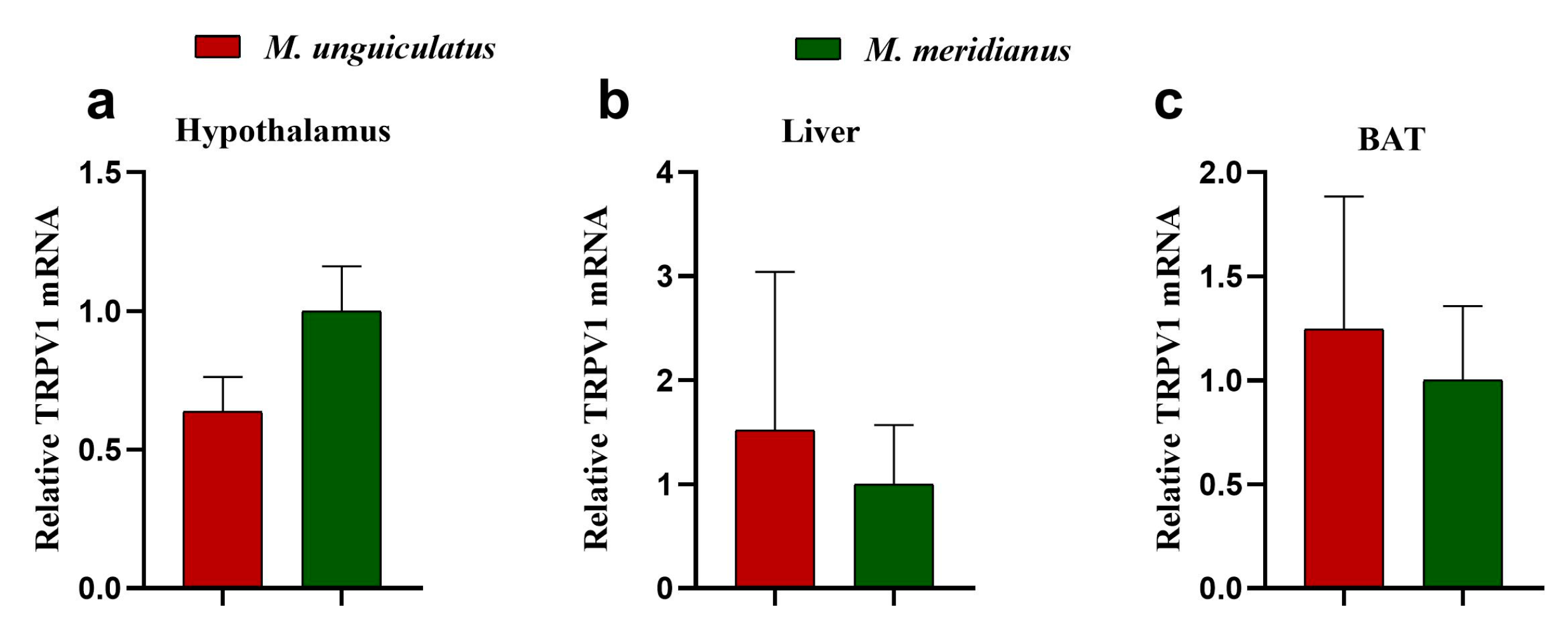
2.3. The mRNA Expression Pattern of TRPV1 Gene in Two Species
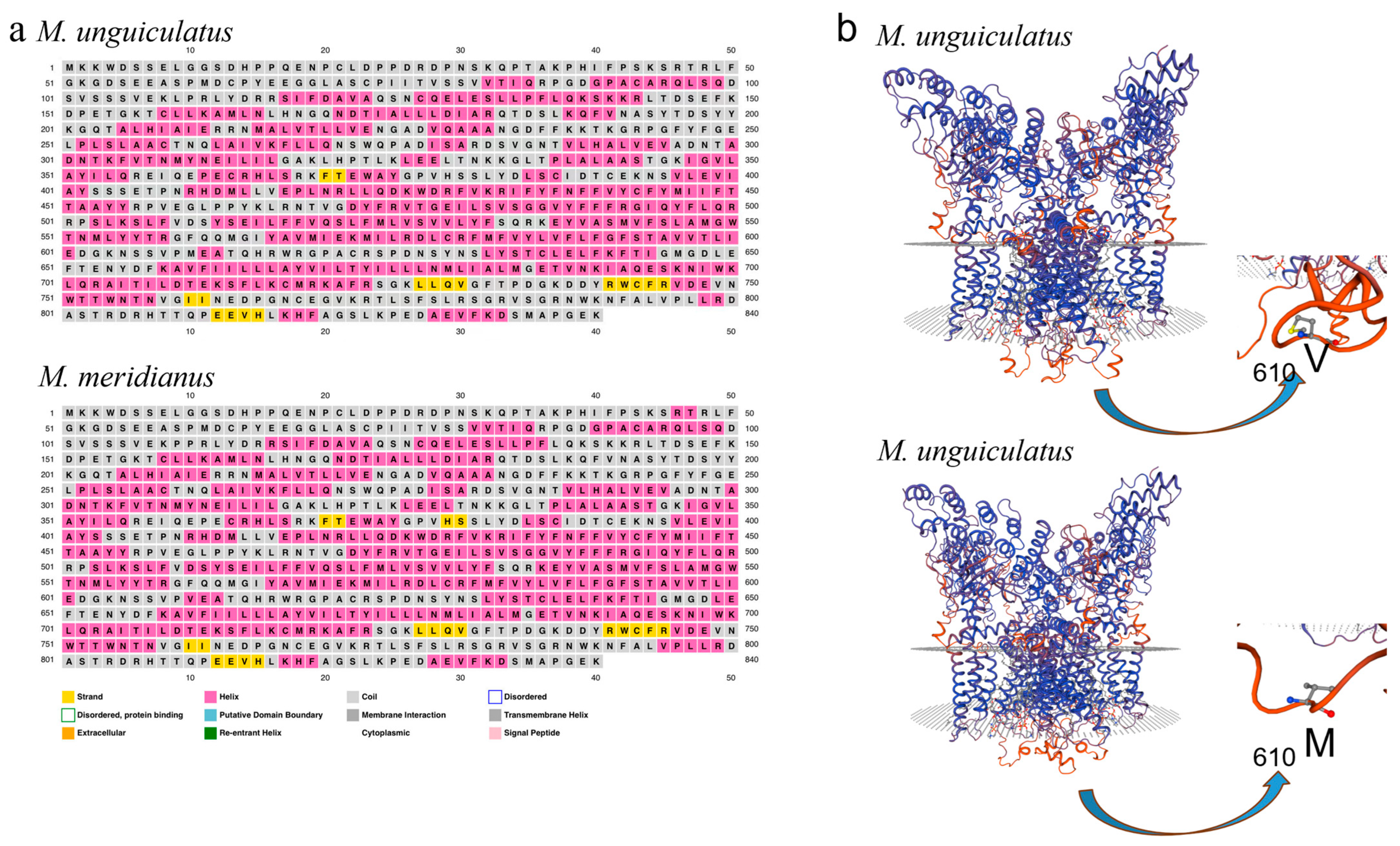
2.4. Homology Analysis of utrpv1 (trpv1 of M. unguiculatus) and mtrpv1 (trpv1 of M. meridianus)
2.5. Structural Prediction of uTRPV1 and mTRPV1
2.6. Cloning of TRPV1 and Ectopic Expression in E. coli System
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Measurement of Rectal Temperature and Metabolic Rate
4.3. Temperature Preference Test
4.4. qPCR Analysis of TRPV1
4.5. Next-Generation Sequencing and TRPV1 Gene Analysis of M. meridianus
4.6. Validation of Missense Mutation Sites and Prediction of Protein Structure
4.7. TRPV1 Cloning and Ectopic Expression in E. coli System
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trisos, C.H.; Merow, C.; Pigot, A. The projected timing of abrupt ecological disruption from climate change. Nature 2020, 580, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.H.; Zhao, Z.J.; Zhang, X.Y.; Zhang, Z.Q.; Xu, D.L.; Xing, X.; Yang, S.M.; Wang, Z.K.; Gao, Y.F.; Yang, M. Research advances and perspectives in mammal physiological ecology in China. Acta Theriol. Sin. 2021, 41, 537–555. [Google Scholar]
- Zub, K.; Szafrańska, P.A.; Konarzewski, M.; Speakman, J.R. Effect of energetic constraints on distribution and winter survival of weasel males. J. Anim. Ecol. 2011, 80, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Tattersall, G.J.; Sinclair, B.J.; Withers, P.C.; Fields, P.A.; Seebacher, F.; Cooper, C.E.; Maloney, S.K. Coping with thermal challenges: Physiological adaptations to environmental temperatures. Compr. Physiol. 2012, 2, 2151–2202. [Google Scholar] [PubMed]
- Morrison, S.F.; Madden, C.J. Central nervous system regulation of brown adipose tissue. Compr. Physiol. 2014, 4, 1677–1713. [Google Scholar]
- Boulant, J.A. Counterpoint: Heat-induced membrane depolarization of hypothalamic neurons: An unlikely mechanism of central thermosensitivity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R1481–R1484. [Google Scholar]
- Cheuvront, S.N.; Kenefick, R.W.; Montain, S.J.; Sawka, M.N. Mechanisms of aerobic performance impairment with heat stress and dehydration. J. Appl. Physiol. 2010, 109, 1989–1995. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Chi, Q.S.; Zhang, X.Y.; Liu, W.; Hao, S.Y.; Wang, D.H. Brown adipose tissue plays thermoregulatory role within the thermoneutral zone in Mongolian gerbils (Meriones unguiculatus). J. Therm. Biol. 2019, 81, 137–145. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Hao, S.Y.; Zhang, M.M.; Zhang, X.Y.; Wang, D.H. Aquaporins, evaporative water loss and thermoregulation in heat-acclimated Mongolian gerbils (Meriones unguiculatus). J. Therm. Biol. 2020, 91, 102641. [Google Scholar] [CrossRef]
- Pan, Q.; Li, M.; Shi, Y.L.; Liu, H.W.; Speakman, J.R.; Wang, D.H. Lipidomics reveals mitochondrial membrane remodeling associated with acute thermoregulation in a rodent with a wide thermoneutral zone. Lipids 2014, 49, 715–730. [Google Scholar] [CrossRef]
- Gladkikh, I.N.; Sintsova, O.V.; Leychenko, E.V.; Kozlov, S.A. TRPV1 ion channel: Structural features, activity modulators, and therapeutic potential. Biochemistry 2021, 86 (Suppl. 1), S50–S70. [Google Scholar] [CrossRef] [PubMed]
- Moran, M.M. TRP ion channels in the nervous system. Curr. Opin. Neurobiol. 2004, 14, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Romanovsky, A.A. Thermoregulation: Some concepts have changed: Functional architecture of the thermoregulatory system. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R37–R46. [Google Scholar] [CrossRef] [PubMed]
- Montell, C.; Rubin, G.M. Molecular characterization of the Drosophila TRP locus: A putative integral membrane protein required for phototransduction. Neuron 1989, 2, 1313–1323. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y. TRP channel classification. Adv. Exp. Med. Biol. 2017, 976, 1–8. [Google Scholar]
- Rosenbaum, T.; Islas, L.D. Molecular physiology of TRPV channels: Controversies and future challenges. Annu. Rev. Physiol. 2023, 85, 293–316. [Google Scholar] [CrossRef]
- Nilius, B.; Owsianik, G. The transient receptor potential family of ion channels. Genome Biol. 2011, 12, 218. [Google Scholar] [CrossRef]
- Díaz-Franulic, I.; Caceres-Molina, J.; Sepulveda, R.V.; Gonzalez-Nilo, F.; Latorre, R. Structure-driven pharmacology of transient receptor potential channel vanilloid 1. Mol. Pharmacol. 2016, 90, 300–308. [Google Scholar] [CrossRef]
- Benítez-Angeles, M.; Morales-Lázaro, S.L.; Juárez-González, E.; Rosenbaum, T. TRPV1: Structure, endogenous agonists, and mechanisms. Int. J. Mol. Sci. 2020, 21, 3421. [Google Scholar] [CrossRef]
- Bal, S.; Sharangi, A.B.; Upadhyay, T.K.; Khan, F.; Pandey, P.; Siddiqui, S.; Saeed, M.; Lee, H.J.; Yadav, D.K. Biomedical and antioxidant potentialities in chilli: Perspectives and way forward. Molecules 2022, 27, 6380. [Google Scholar] [CrossRef]
- Zhang, X.; El-Demerdash, N.; Falck, J.R.; Munnuri, S.; Koehler, R.C.; Yang, Z.J. The contribution of TRPV1 channel to 20-HETE-aggravated ischemic neuronal injury. Prostaglandins Other Lipid Mediat. 2018, 137, 63–68. [Google Scholar] [CrossRef]
- Pang, Z.X.; Sakamoto, T.; Tiwari, V.; Kim, Y.S.; Yang, F.; Dong, X.Z.; Güler, A.D.; Guan, Y.; Caterina, M.J. Selective keratinocyte stimulation is sufficient to evoke nociception in mice. Pain 2015, 156, 656–665. [Google Scholar] [CrossRef]
- Davis, J.B.; Gray, J.; Gunthorpe, M.J.; Hatcher, J.P.; Sheardown, S.A. Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature 2000, 405, 183–187. [Google Scholar] [CrossRef]
- Caterina, M.J.; Leffler, A.; Malmberg, A.B.; Martin, W.J.; Trafton, J.; Petersen-Zeitz, K.R.; Koltzenburg, M.; Basbaum, A.I.; Julius, D. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 2000, 288, 306–313. [Google Scholar] [CrossRef]
- Laursen, W.J.; Schneider, E.R.; Merriman, D.K.; Bagriantsev, S.N.; Gracheva, E.O. Low-cost functional plasticity of TRPV1 supports heat tolerance in squirrels and camels. Proc. Natl. Acad. Sci. USA 2016, 113, 11342–11347. [Google Scholar] [CrossRef]
- Masamoto, Y.; Kawabata, F.; Fushiki, T. Intragastric administration of TRPV1, TRPV3, TRPM8, and TRPA1 agonists modulates autonomic thermoregulation in different manners in mice. Biosci. Biotechnol. Biochem. 2009, 73, 1021–1027. [Google Scholar] [CrossRef]
- Jordt, S.E.; Julius, D. Molecular basis for species-specific sensitivity to “Hot” chili peppers. Cell 2002, 108, 421–430. [Google Scholar] [CrossRef]
- Jordt, S.E.; Tominaga, M.; Julius, D. Acid potentiation of the capsaicin receptor determined by a key extracellular site. Proc. Natl. Acad. Sci. USA 2000, 97, 8134–8139. [Google Scholar] [CrossRef]
- Yang, F.; Xiao, X.; Cheng, W.; Yang, W.; Yu, P.L.; Song, Z.Z.; Vladimir, Y.Y.; Zheng, J. Structural mechanism underlying capsaicin binding and activation of the TRPV1 ion channel. Nat. Chem. Biol. 2015, 11, 518–524. [Google Scholar] [CrossRef]
- Ito, M.; Jiang, W.; Sato, J.J.; Zhen, Q.; Kamiya, H. Molecular phylogeny of the subfamily Gerbillinae (Muridae, Rodentia) with emphasis on species living in the Xinjiang-Uygur Autonomous Region of China and based on the mitochondrial cytochrome b and cytochrome c oxidase subunit II genes. Zool. Sci. 2010, 27, 269–278. [Google Scholar] [CrossRef]
- Ding, B.Y.; Chi, Q.S.; Liu, W.; Shi, Y.L.; Wang, D.H. Thermal biology of two sympatric gerbil species: The physiological basis of temporal partitioning. J. Therm. Biol. 2018, 74, 241–248. [Google Scholar] [CrossRef]
- Lou, S.L.; Zhang, X.Y.; Wang, D.H. HSP70 plays a role in the defense of acute and chronic heat stress in Mongolian gerbils (Meriones unguiculatus). J. Therm. Biol. 2019, 86, 102452. [Google Scholar] [CrossRef]
- Wen, J.; Bo, T.B.; Zhang, X.Y.; Wang, Z.X.; Wang, D.H. Thermo-TRPs and gut microbiota are involved in thermogenesis and energy metabolism during low temperature exposure of obese mice. J. Exp. Biol. 2020, 223 Pt 11, jeb218974. [Google Scholar] [CrossRef]
- Wen, J.; Bo, T.B.; Zhao, Z.J.; Wang, D.H. Role of transient receptor potential vanilloid-1 in behavioral thermoregulation of the Mongolian gerbil Meriones unguiculatus (Milne-Edwards, 1867). Integr. Zool. 2021, 17, 608–618. [Google Scholar] [CrossRef]
- Dale, V.H.; Beyeler, S.C. Challenges in the development and use of ecological indicators. Ecol. Indic. 2001, 1, 3–10. [Google Scholar] [CrossRef]
- Myers, P.; Lundrigan, B.L.; Hoffman, S.; Haraminac, A.P.; Seto, S.H. Climate-induced changes in the small mammal communities of the northern great lakes region. Glob. Chang. Biol. 2010, 15, 1434–1454. [Google Scholar] [CrossRef]
- Jarema, S.I.; Jason, S.; Mcgill, B.J.; Humphries, M.M. Variation in abundance across a species’ range predicts climate change responses in the range interior will exceed those at the edge: A case study with north American beaver. Glob. Chang. Biol. 2009, 15, 508–522. [Google Scholar] [CrossRef]
- Ma, R.J.; Jiang, Z.G. Impact of global climate change on wildlife. Acta Ecol. Sin. 2005, 11, 269–274. [Google Scholar]
- Wu, X.D.; Yuan, S.; Fu, H.P.; Zhang, X.D.; Zhang, F.S.; Gao, Q.R. Responses of dominant rodentspecies to climate change in different disturbed habitats in the Alashan desert. Acta Ecol. Sin. 2016, 36, 1765–1773. [Google Scholar]
- Wang, D.H.; Wang, Y.S.; Wang, Z.W. Metabolism and thermoregulation in the Mongolian gerbil Meriones unguiculatus. Acta Theriol. 2000, 45, 183–192. [Google Scholar] [CrossRef]
- Zoltán, W.; Andrea, B.; Ferenc, Ö.; Katalin, J.; Csaba, V.; György, D.; Gerda, S.; Zoltán, O. Functionally important amino acid residues in the transient receptor potential vanilloid 1 (TRPV1) ion channel—An overview of the current mutational data. Mol. Pain 2013, 9, 1–29. [Google Scholar]
- Miki, T.; Kaori, H.; Nozomi, O.; Tatsuki, K.; Kenichi, K.; Kenji, I.; Toshitaka, M.; Yasuo, M.; Masaki, U. Structure determination of the human TRPV1 ankyrin-repeat domain under nonreducing conditions. Acta Crystallogr. F. Struct. Biol. Commun. 2020, 76 Pt 3, 130–137. [Google Scholar]
- Dosey, T.L.; Wang, Z.; Fan, G.Z.; Zhang, Z.X.; Serysheva, I.I.; Chiu, W.; Wensel, T.G. Structures of TRPV2 in distinct conformations provide insight into role of the pore turret. Nat. Struct. Mol. Biol. 2019, 26, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.L.; Lu, X.C.; Wang, Y.F.; Xu, L.Z.; Chen, X.Y.; Yang, F.; Lai, R. A paradigm of thermal adaptation in penguins and elephants by tuning cold activation in TRPM8. Proc. Natl. Acad. Sci. USA 2020, 117, 8633–8638. [Google Scholar] [CrossRef] [PubMed]
- Frankham, R. Stress and adaptation in conservation genetics. J. Evol. Biol. 2005, 18, 750–755. [Google Scholar] [CrossRef]
- Zalapa, J.E.; Brunet, J.; Guries, R.P. The extent of hybridization and its impact on the genetic diversity and population structure of an invasive tree, Ulmus pumila (Ulmaceae). Evol. Appl. 2010, 3, 157–168. [Google Scholar] [CrossRef]
- Merilä, J.; Laurila, A.; Lindgren, B. Variation in the degree and costs of adaptive phenotypic plasticity among Rana temporaria populations. J. Evol. Biol. 2004, 17, 1132–1140. [Google Scholar] [CrossRef]
- Birch, J. Natural selection and the maximization of fitness. Biol. Rev. Camb. Philos. Soc. 2016, 91, 712–727. [Google Scholar] [CrossRef]
- Khakisahneh, S.; Zhang, X.Y.; Zouri, Z.; Wang, D.H. Gut microbiota and host thermoregulation in response to ambient temperature fluctuations. mSystems 2020, 5, e00514–e00520. [Google Scholar] [CrossRef]
- Wang, D.H.; Wang, Z.W. Seasonal variations in thermogenesis and energy requirements of plateau pikas (Ochotona curzoniae) and root voles (Microtus oeconomus). Acta Theriol. 1996, 41, 225–236. [Google Scholar] [CrossRef]
- Li, X.S.; Wang, D.H. Photoperiod and temperature can regulate body mass, serum leptin concentration, and uncoupling protein 1 in Brandt’s voles (Lasiopodomys brandtii) and Mongolian gerbils (Meriones unguiculatus). Physiol. Biochem. Zool. 2007, 80, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Chi, Q.S.; Wan, X.R.; Geiser, F.; Wang, D.H. Fasting-induced daily torpor in desert hamsters (Phodopus roborovskii). Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2016, 199, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.G.; Yan, Z.C.; Wang, D.H. Physiological and biochemical basis of basal metabolic rates in Brandt's voles (Lasiopodomys brandtii) and Mongolian gerbils (Meriones unguiculatus). Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2010, 157, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, P.C.; Zhang, S.S.; Yang, J.; Li, G.C.; Huang, L.Q.; Wang, C.Z. Functional analysis of a bitter gustatory receptor highly expressed in the larval maxillary galea of Helicoverpa armigera. PLoS Genet. 2022, 18, e1010455. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Species | Statistical Summary | |
|---|---|---|---|
| M. unguiculatus | M. meridianus | p-Value | |
| Body mass (g) | 38.7 ± 1.5 | 48.2 ± 2.7 | <0.01 |
| Body length (mm) | 101.7 ± 1.8 | 107.4 ± 2.5 | ns |
| Ear length (mm) | 11.9 ± 0.54 | 15.7 ± 1.27 | <0.01 |
| Tail length (mm) | 93.8 ± 2.68 | 100.1 ± 4.33 | ns |
| Metapodial length (mm) | 22.6 ± 0.34 | 26.1 ± 0.83 | <0.05 |
| Species | Total Length (bp) | Total Variants | Nonsense Variants | Homology of cDNA | Protein Length | Homology of Protein |
|---|---|---|---|---|---|---|
| M. unguiculatus | 2523 | 18 | 2 | 99.28% | 840 | 99.76% |
| M. meridianus |
| Mutation 1 | Mutation 2 | |
|---|---|---|
| M. unguiculatus | 100DSVSSSVEKPPRLYDRRSIF … | … 597VTLIEDGKNSSVPVEATQHR |
| M. meridianus | 100DSVSSSVEKLPRLYDRRSIF … | … 597VTLIEDGKNSSVPMEATQHR |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, B.; Zhang, X.-Y.; Yuan, S.; Fu, H.-P.; Wang, C.-Z.; Wang, D.-H. Genetic Diversity of a Heat Activated Channel—TRPV1 in Two Desert Gerbil Species with Different Heat Sensitivity. Int. J. Mol. Sci. 2023, 24, 9123. https://doi.org/10.3390/ijms24119123
Wang B, Zhang X-Y, Yuan S, Fu H-P, Wang C-Z, Wang D-H. Genetic Diversity of a Heat Activated Channel—TRPV1 in Two Desert Gerbil Species with Different Heat Sensitivity. International Journal of Molecular Sciences. 2023; 24(11):9123. https://doi.org/10.3390/ijms24119123
Chicago/Turabian StyleWang, Bing, Xue-Ying Zhang, Shuai Yuan, He-Ping Fu, Chen-Zhu Wang, and De-Hua Wang. 2023. "Genetic Diversity of a Heat Activated Channel—TRPV1 in Two Desert Gerbil Species with Different Heat Sensitivity" International Journal of Molecular Sciences 24, no. 11: 9123. https://doi.org/10.3390/ijms24119123