
Kidney-Related Function of Mitochondrial Protein Mitoregulin

 , , and
, , and 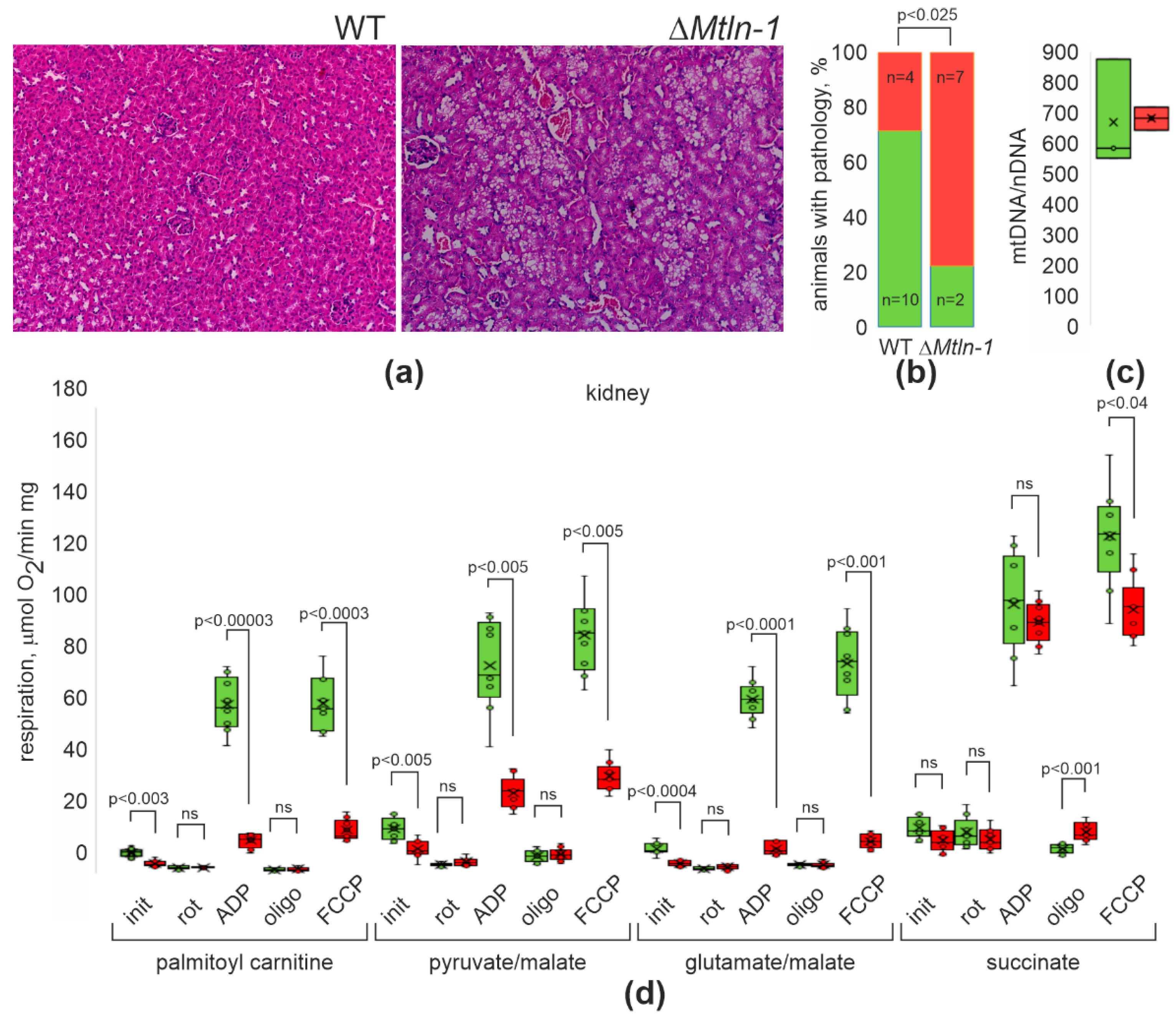{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Kidney Morphology Changes upon Mtln Inactivation
2.2. Respiration of Kidney Mitochondria upon Mtln Inactivation
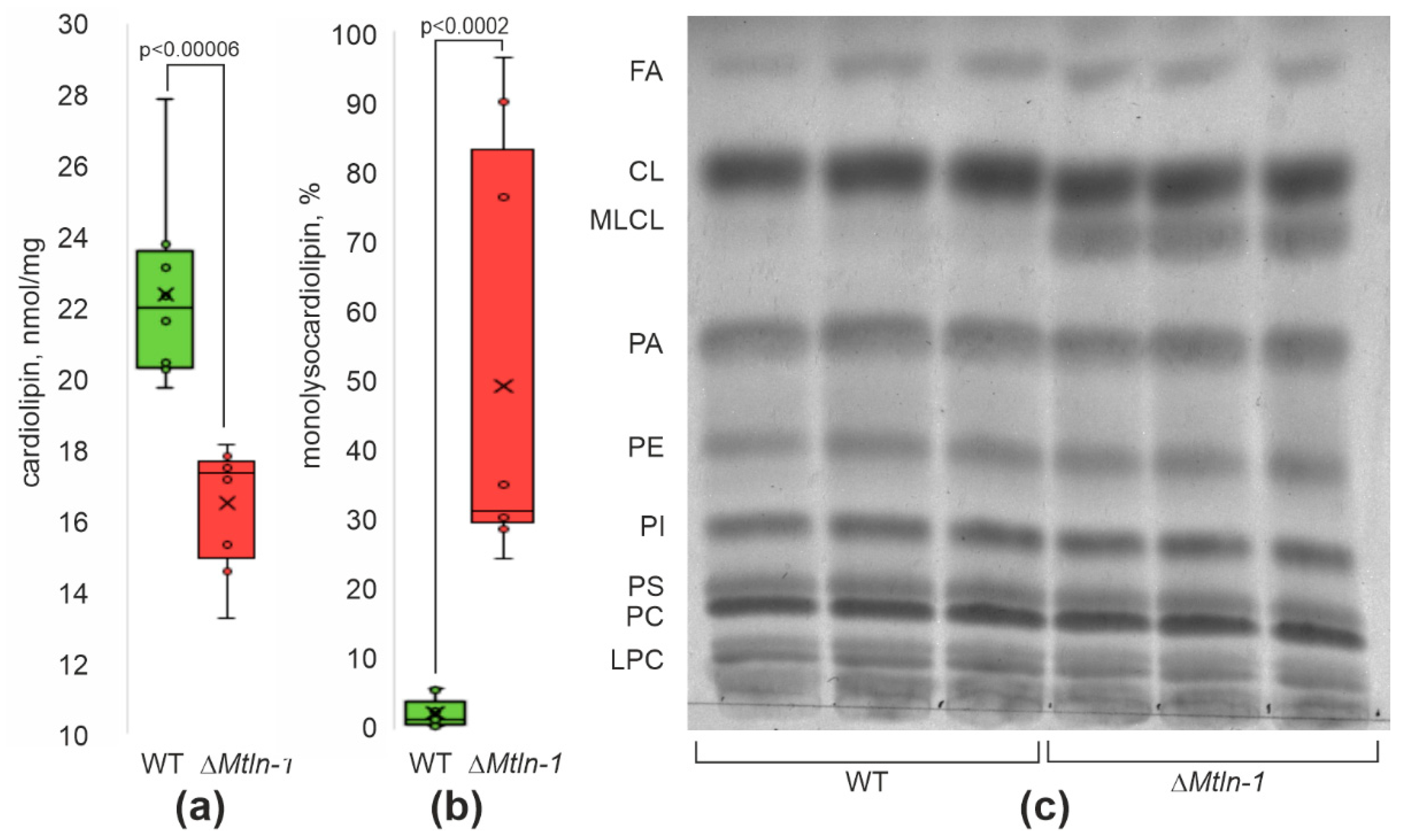
2.3. Cardiolipin Damage in Kidney Mitochondria upon Mtln Inactivation
2.4. Kidney Mitochondrial Creatine Kinase Oligomerization and Activity Depend on Mtln Functionality
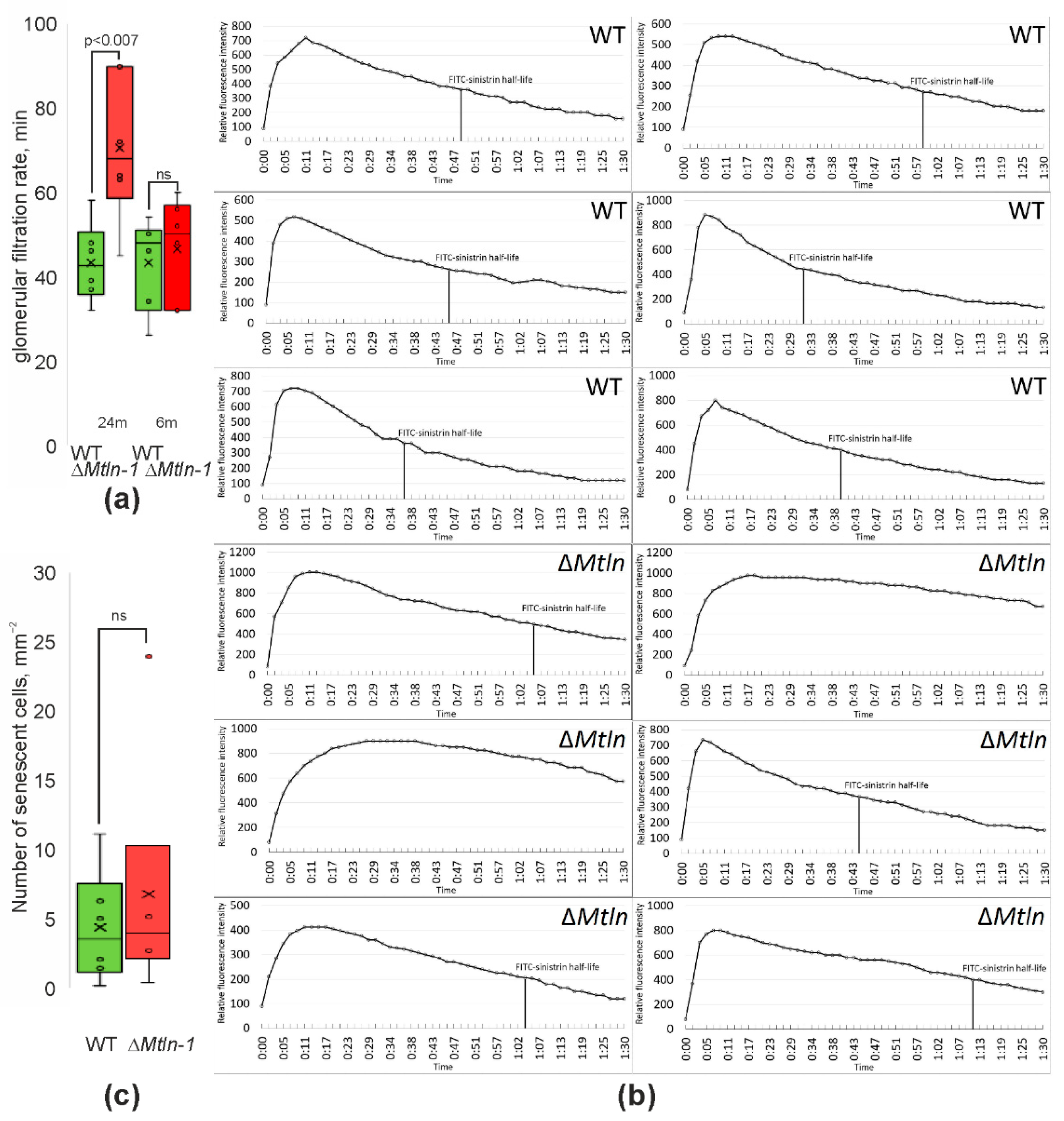
2.5. Reduction of Glomerular Filtration Rates upon Mtln Inactivation
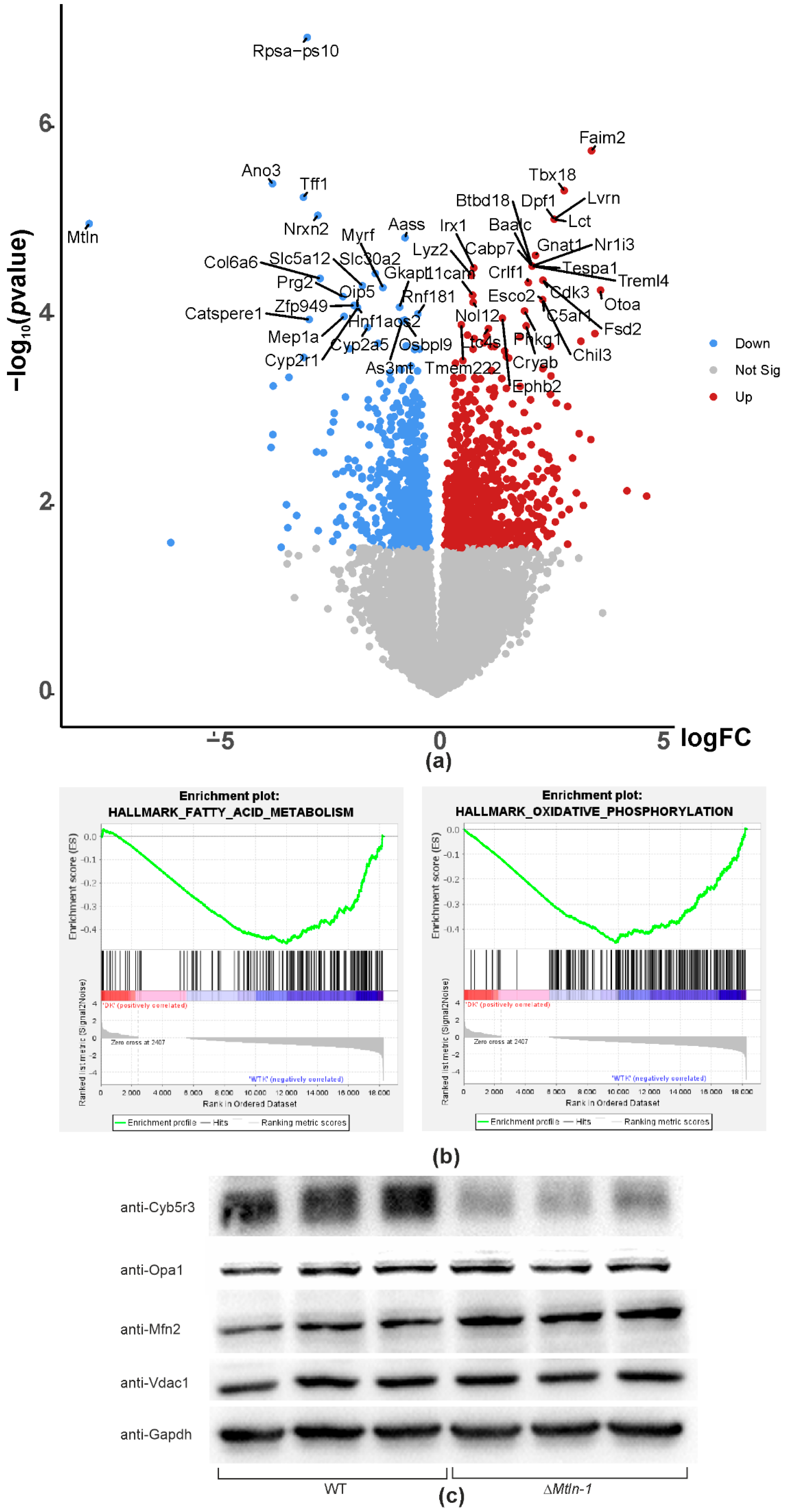
2.6. Mtln Inactivation Perturbs Kidney Gene Expression
3. Discussion
4. Materials and Methods
4.1. Mice Housing and Breeding
4.2. Glomerular Filtration Rate
4.3. Histology
4.4. Tissue Processing, Mitochondria Preparation, and Respiration Analysis
4.5. Lipid Analysis
4.6. Whole Transcriptome Assay
4.7. Immunoblotting
4.8. qPCR
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ruiz-Orera, J.; Albà, M.M. Translation of Small Open Reading Frames: Roles in Regulation and Evolutionary Innovation. Trends Genet. 2019, 35, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Makarewich, C.A.; Olson, E.N. Mining for Micropeptides. Trends Cell Biol. 2017, 27, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Couso, J.-P.; Patraquim, P. Classification and Function of Small Open Reading Frames. Nat. Rev. Mol. Cell Biol. 2017, 18, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Chugunova, A.; Navalayeu, T.; Dontsova, O.; Sergiev, P. Mining for Small Translated ORFs. J. Proteome Res. 2018, 17, 1–11. [Google Scholar] [CrossRef]
- Rath, S.; Sharma, R.; Gupta, R.; Ast, T.; Chan, C.; Durham, T.J.; Goodman, R.P.; Grabarek, Z.; Haas, M.E.; Hung, W.H.W.; et al. MitoCarta3.0: An Updated Mitochondrial Proteome Now with Sub-Organelle Localization and Pathway Annotations. Nucleic Acids Res. 2021, 49, D1541–D1547. [Google Scholar] [CrossRef] [PubMed]
- Rathore, A.; Chu, Q.; Tan, D.; Martinez, T.F.; Donaldson, C.J.; Diedrich, J.K.; Yates, J.R.; Saghatelian, A. MIEF1 Microprotein Regulates Mitochondrial Translation. Biochemistry 2018, 57, 5564–5575. [Google Scholar] [CrossRef]
- Brown, A.; Rathore, S.; Kimanius, D.; Aibara, S.; Bai, X.-C.; Rorbach, J.; Amunts, A.; Ramakrishnan, V. Structures of the Human Mitochondrial Ribosome in Native States of Assembly. Nat. Struct. Mol. Biol. 2017, 24, 866–869. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Brunner, A.-D.; Cogan, J.Z.; Nuñez, J.K.; Fields, A.P.; Adamson, B.; Itzhak, D.N.; Li, J.Y.; Mann, M.; Leonetti, M.D.; et al. Pervasive Functional Translation of Noncanonical Human Open Reading Frames. Science 2020, 367, 1140–1146. [Google Scholar] [CrossRef]
- Zhang, S.; Reljić, B.; Liang, C.; Kerouanton, B.; Francisco, J.C.; Peh, J.H.; Mary, C.; Jagannathan, N.S.; Olexiouk, V.; Tang, C.; et al. Mitochondrial Peptide BRAWNIN Is Essential for Vertebrate Respiratory Complex III Assembly. Nat. Commun. 2020, 11, 1312. [Google Scholar] [CrossRef]
- Chu, Q.; Martinez, T.F.; Novak, S.W.; Donaldson, C.J.; Tan, D.; Vaughan, J.M.; Chang, T.; Diedrich, J.K.; Andrade, L.; Kim, A.; et al. Regulation of the ER Stress Response by a Mitochondrial Microprotein. Nat. Commun. 2019, 10, 4883. [Google Scholar] [CrossRef]
- Sergiev, P.V.; Rubtsova, M.P. Little but Loud. The Diversity of Functions of Small Proteins and Peptides—Translational Products of Short Reading Frames. Biochem. Moscow. 2021, 86, 1139–1150. [Google Scholar] [CrossRef] [PubMed]
- Chugunova, A.; Loseva, E.; Mazin, P.; Mitina, A.; Navalayeu, T.; Bilan, D.; Vishnyakova, P.; Marey, M.; Golovina, A.; Serebryakova, M.; et al. LINC00116 Codes for a Mitochondrial Peptide Linking Respiration and Lipid Metabolism. Proc. Natl. Acad. Sci. USA 2019, 116, 4940–4945. [Google Scholar] [CrossRef] [PubMed]
- Stein, C.S.; Jadiya, P.; Zhang, X.; McLendon, J.M.; Abouassaly, G.M.; Witmer, N.H.; Anderson, E.J.; Elrod, J.W.; Boudreau, R.L. Mitoregulin: A LncRNA-Encoded Microprotein That Supports Mitochondrial Supercomplexes and Respiratory Efficiency. Cell Rep. 2018, 23, 3710–3720.e8. [Google Scholar] [CrossRef] [PubMed]
- Makarewich, C.A.; Baskin, K.K.; Munir, A.Z.; Bezprozvannaya, S.; Sharma, G.; Khemtong, C.; Shah, A.M.; McAnally, J.R.; Malloy, C.R.; Szweda, L.I.; et al. MOXI Is a Mitochondrial Micropeptide That Enhances Fatty Acid β-Oxidation. Cell Rep. 2018, 23, 3701–3709. [Google Scholar] [CrossRef]
- Friesen, M.; Warren, C.R.; Yu, H.; Toyohara, T.; Ding, Q.; Florido, M.H.C.; Sayre, C.; Pope, B.D.; Goff, L.A.; Rinn, J.L.; et al. Mitoregulin Controls β-Oxidation in Human and Mouse Adipocytes. Stem Cell Rep. 2020, 14, 590–602. [Google Scholar] [CrossRef]
- Averina, O.A.; Permyakov, O.A.; Emelianova, M.A.; Grigoryeva, O.O.; Gulyaev, M.V.; Pavlova, O.S.; Mariasina, S.S.; Frolova, O.Y.; Kurkina, M.V.; Baydakova, G.V.; et al. Mitochondrial Peptide Mtln Contributes to Oxidative Metabolism in Mice. Biochimie 2023, 204, 136–139. [Google Scholar] [CrossRef]
- Choi, M.; Kang, K.W. Mitoregulin Controls Mitochondrial Function and Stress-Adaptation Response during Early Phase of Endoplasmic Reticulum Stress in Breast Cancer Cells. Biochim. Biophys. Acta (BBA.)-Mol. Basis Dis. 2023, 1869, 166570. [Google Scholar] [CrossRef]
- Lin, Y.-F.; Xiao, M.-H.; Chen, H.-X.; Meng, Y.; Zhao, N.; Yang, L.; Tang, H.; Wang, J.-L.; Liu, X.; Zhu, Y.; et al. A Novel Mitochondrial Micropeptide MPM Enhances Mitochondrial Respiratory Activity and Promotes Myogenic Differentiation. Cell Death Dis. 2019, 10, 528. [Google Scholar] [CrossRef]
- Wang, L.; Fan, J.; Han, L.; Qi, H.; Wang, Y.; Wang, H.; Chen, S.; Du, L.; Li, S.; Zhang, Y.; et al. The Micropeptide LEMP Plays an Evolutionarily Conserved Role in Myogenesis. Cell Death Dis. 2020, 11, 357. [Google Scholar] [CrossRef]
- Averina, O.A.; Permyakov, O.A.; Emelianova, M.A.; Grigoryeva, O.O.; Lovat, M.L.; Egorova, A.E.; Grinchenko, A.V.; Kumeiko, V.V.; Marey, M.V.; Manskikh, V.N.; et al. Mitoregulin Contributes to Creatine Shuttling and Cardiolipin Protection in Mice Muscle. Int. J. Mol. Sci. 2023, 24, 7589. [Google Scholar] [CrossRef]
- Galvan, D.L.; Green, N.H.; Danesh, F.R. The Hallmarks of Mitochondrial Dysfunction in Chronic Kidney Disease. Kidney Int. 2017, 92, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, J. Renal Manifestations of Genetic Mitochondrial Disease. Int. J. Nephrol. Renov. Dis. 2014, 7, 57. [Google Scholar] [CrossRef] [PubMed]
- Finsterer, J.; Scorza, F. Renal Manifestations of Primary Mitochondrial Disorders. Biomed. Rep. 2017, 6, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Gidlund, E.-K.; Witasp, A.; Qureshi, A.R.; Söderberg, M.; Thorell, A.; Nader, G.A.; Barany, P.; Stenvinkel, P.; Von Walden, F. Reduced Skeletal Muscle Expression of Mitochondrial-Derived Peptides Humanin and MOTS-C and Nrf2 in Chronic Kidney Disease. Am. J. Physiol.-Ren. Physiol. 2019, 317, F1122–F1131. [Google Scholar] [CrossRef]
- Li, J.; Qu, X.; Guan, C.; Luo, N.; Chen, H.; Li, A.; Zhuang, H.; Yang, J.; Diao, H.; Zeng, S.; et al. Mitochondrial Micropeptide MOXI Promotes Fibrotic Gene Transcription by Translocation to the Nucleus and Bridging N-Acetyltransferase 14 with Transcription Factor c-Jun. Kidney Int. 2023, 103, 886–902. [Google Scholar] [CrossRef]
- Scarfe, L.; Schock-Kusch, D.; Ressel, L.; Friedemann, J.; Shulhevich, Y.; Murray, P.; Wilm, B.; de Caestecker, M. Transdermal Measurement of Glomerular Filtration Rate in Mice. JoVE 2018, 140, 58520. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Leboucher, G.P.; Tsai, Y.C.; Yang, M.; Shaw, K.C.; Zhou, M.; Veenstra, T.D.; Glickman, M.H.; Weissman, A.M. Stress-Induced Phosphorylation and Proteasomal Degradation of Mitofusin 2 Facilitates Mitochondrial Fragmentation and Apoptosis. Mol. Cell 2012, 47, 547–557. [Google Scholar] [CrossRef]
- Keyes, S.R.; Cinti, D.L. Biochemical Properties of Cytochrome B5-Dependent Microsomal Fatty Acid Elongation and Identification of Products. J. Biol. Chem. 1980, 255, 11357–11364. [Google Scholar] [CrossRef]
- Reddy, V.V.; Kupfer, D.; Caspi, E. Mechanism of C-5 Double Bond Introduction in the Biosynthesis of Cholesterol by Rat Liver Microsomes. J. Biol. Chem. 1977, 252, 2797–2801. [Google Scholar] [CrossRef]
- Forbes, J.M.; Thorburn, D.R. Mitochondrial Dysfunction in Diabetic Kidney Disease. Nat. Rev. Nephrol. 2018, 14, 291–312. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Sheu, S.-S. (Eds.) Pharmacology of Mitochondria; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2017; ISBN 978-3-319-57311-3. [Google Scholar]
- Crawford, S.O.; Hoogeveen, R.C.; Brancati, F.L.; Astor, B.C.; Ballantyne, C.M.; Schmidt, M.I.; Young, J.H. Association of Blood Lactate with Type 2 Diabetes: The Atherosclerosis Risk in Communities Carotid MRI Study. Int. J. Epidemiol. 2010, 39, 1647–1655. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.C. A Mitochondrial Paradigm of Metabolic and Degenerative Diseases, Aging, and Cancer: A Dawn for Evolutionary Medicine. Annu. Rev. Genet. 2005, 39, 359–407. [Google Scholar] [CrossRef]
- Miller, B.; Kim, S.-J.; Kumagai, H.; Yen, K.; Cohen, P. Mitochondria-Derived Peptides in Aging and Healthspan. J. Clin. Investig. 2022, 132, e158449. [Google Scholar] [CrossRef] [PubMed]
- Sultanova, R.F.; Schibalski, R.; Yankelevich, I.A.; Stadler, K.; Ilatovskaya, D.V. Sex Differences in Renal Mitochondrial Function: A Hormone-Gous Opportunity for Research. Am. J. Physiol.-Ren. Physiol. 2020, 319, F1117–F1124. [Google Scholar] [CrossRef] [PubMed]
- Maronpot, R.R. (Ed.) Pathology of the Mouse: Reference and Atlas, 1st ed; Cache River Press: Saint Louis, MO, USA, 1999; ISBN 978-1-889899-02-2. [Google Scholar]
- Hard, C.C.; Alden, C.L.; Bruner, R.H.G.; Frith, C.H.; Lewis, R.M.; Owen, R.A.; Krieg, K.; Durchfeld-Meyer, B. Non-Proliferative Lesions of the Kidney and Lower Urinary Tract in the Rat. In URG-1, Guides for Toxicological Pathology; STP/ARP/AFIP: Washington, DC, USA, 1999. [Google Scholar]
- Averina, O.A.; Laptev, I.G.; Emelianova, M.A.; Permyakov, O.A.; Mariasina, S.S.; Nikiforova, A.I.; Manskikh, V.N.; Grigorieva, O.O.; Bolikhova, A.K.; Kalabin, G.A.; et al. Mitochondrial RRNA Methylation by Mettl15 Contributes to the Exercise and Learning Capability in Mice. Int. J. Mol. Sci. 2022, 23, 6056. [Google Scholar] [CrossRef] [PubMed]
- Luna, L. Histopathologic Methods and. Color Atlas of Special Stains and Tissue Artifacts; American Histolabs: Gaithersburg MD, USA, 1993. [Google Scholar]
- Lojda, Z.; Gossrau, R.; Schiebler, T.H. Enzyme Histochemistry: A Laboratory Manual; Springer: Berlin/Heidelberg, Germany, 1979; ISBN 978-3-540-09269-8. [Google Scholar]
- Debacq-Chainiaux, F.; Erusalimsky, J.D.; Campisi, J.; Toussaint, O. Protocols to Detect Senescence-Associated Beta-Galactosidase (SA-Βgal) Activity, a Biomarker of Senescent Cells in Culture and in Vivo. Nat. Protoc. 2009, 4, 1798–1806. [Google Scholar] [CrossRef] [PubMed]
- Vyssokikh, M.Y.; Holtze, S.; Averina, O.A.; Lyamzaev, K.G.; Panteleeva, A.A.; Marey, M.V.; Zinovkin, R.A.; Severin, F.F.; Skulachev, M.V.; Fasel, N.; et al. Mild Depolarization of the Inner Mitochondrial Membrane Is a Crucial Component of an Anti-Aging Program. Proc. Natl. Acad. Sci. USA 2020, 117, 6491–6501. [Google Scholar] [CrossRef]
- Gnaiger, E.; Kuznetsov, A.V.; Schneeberger, S.; Seiler, R.; Brandacher, G.; Steurer, W.; Margreiter, R. Mitochondria in the Cold. In Life in the Cold; Heldmaier, G., Klingenspor, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 431–442. ISBN 978-3-642-08682-3. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Pinault, M.; Guimaraes, C.; Dumas, J.; Servais, S.; Chevalier, S.; Besson, P.; Goupille, C. A 1D High Performance Thin Layer Chromatography Method Validated to Quantify Phospholipids Including Cardiolipin and Monolysocardiolipin from Biological Samples. Eur. J. Lipid. Sci. Technol. 2020, 122, 1900240. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Averina, O.A.; Permyakov, O.A.; Emelianova, M.A.; Guseva, E.A.; Grigoryeva, O.O.; Lovat, M.L.; Egorova, A.E.; Grinchenko, A.V.; Kumeiko, V.V.; Marey, M.V.; et al. Kidney-Related Function of Mitochondrial Protein Mitoregulin. Int. J. Mol. Sci. 2023, 24, 9106. https://doi.org/10.3390/ijms24109106
Averina OA, Permyakov OA, Emelianova MA, Guseva EA, Grigoryeva OO, Lovat ML, Egorova AE, Grinchenko AV, Kumeiko VV, Marey MV, et al. Kidney-Related Function of Mitochondrial Protein Mitoregulin. International Journal of Molecular Sciences. 2023; 24(10):9106. https://doi.org/10.3390/ijms24109106
Chicago/Turabian StyleAverina, Olga A., Oleg A. Permyakov, Mariia A. Emelianova, Ekaterina A. Guseva, Olga O. Grigoryeva, Maxim L. Lovat, Anna E. Egorova, Andrei V. Grinchenko, Vadim V. Kumeiko, Maria V. Marey, and et al. 2023. "Kidney-Related Function of Mitochondrial Protein Mitoregulin" International Journal of Molecular Sciences 24, no. 10: 9106. https://doi.org/10.3390/ijms24109106