

Plasmatic MicroRNAs and Treatment Outcomes of Patients with Metastatic Castration-Resistant Prostate Cancer: A Hospital-Based Cohort Study and In Silico Analysis
 , ,
, ,
Abstract
:1. Introduction
2. Results
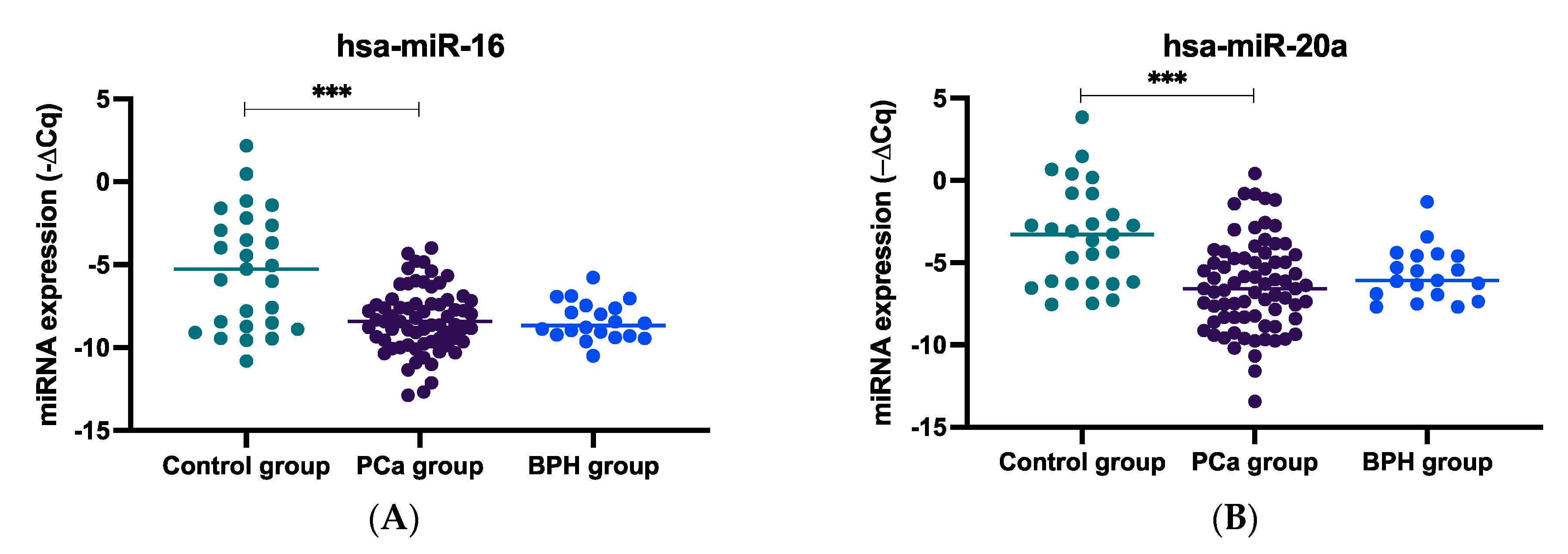
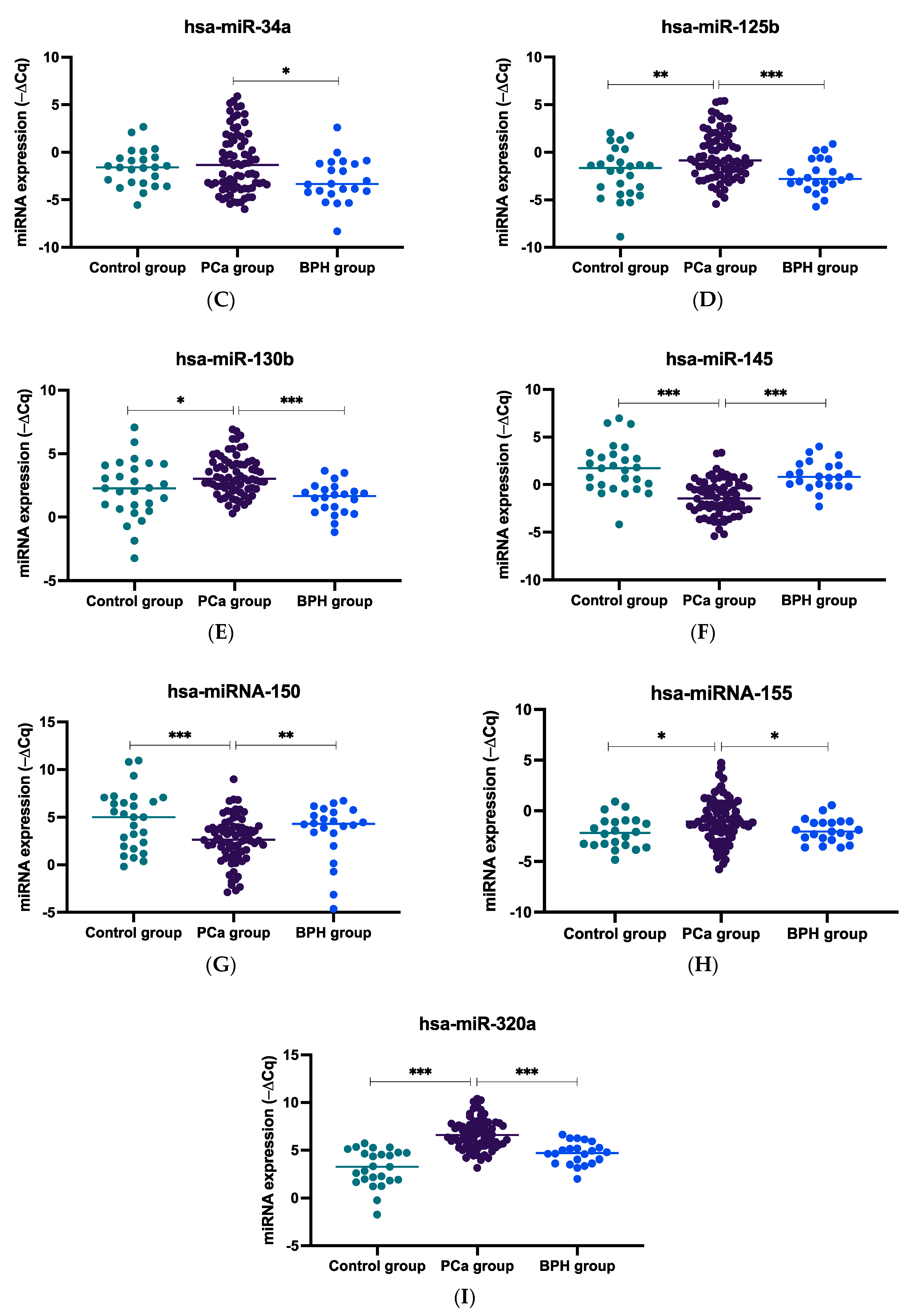
2.1. miRNA Expression Levels in Plasma Samples of mCRPC, BPH, and Healthy Controls
2.2. Associations between the miRNA Expression and Patient Demographic and Clinicopathological Factors
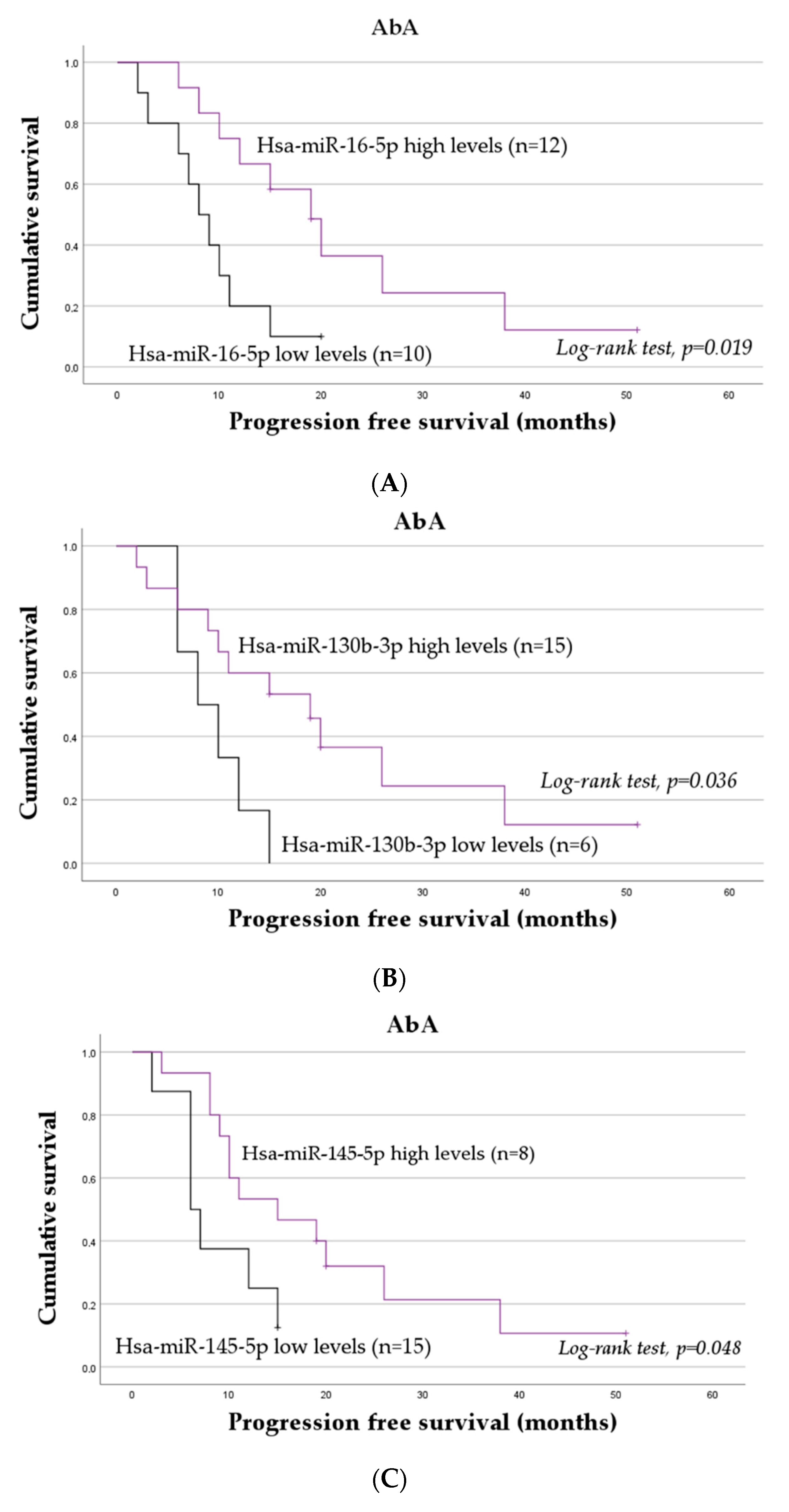
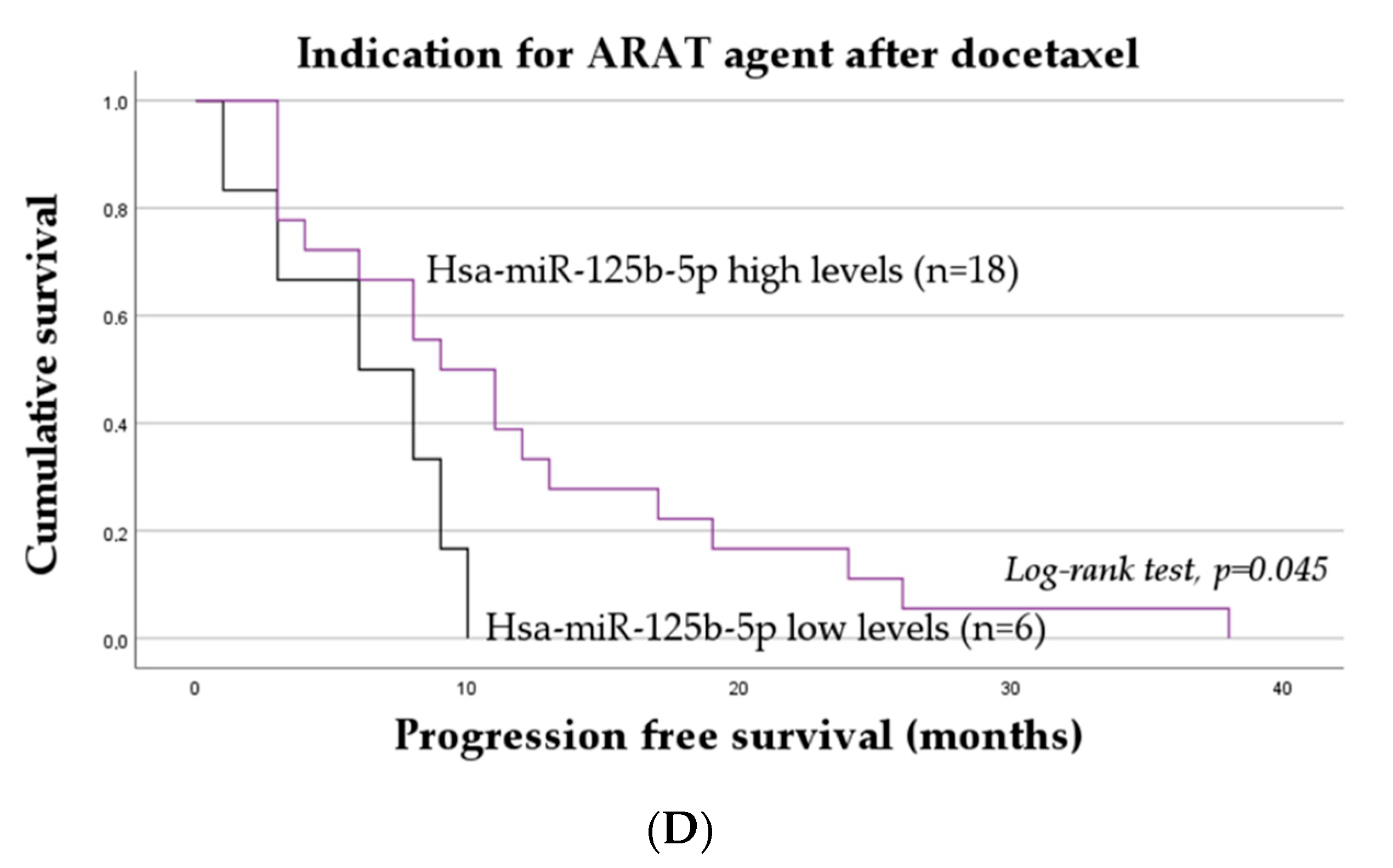
2.3. The Impact of miRNAs on the Progression-Free Survival of mCRPC Patients
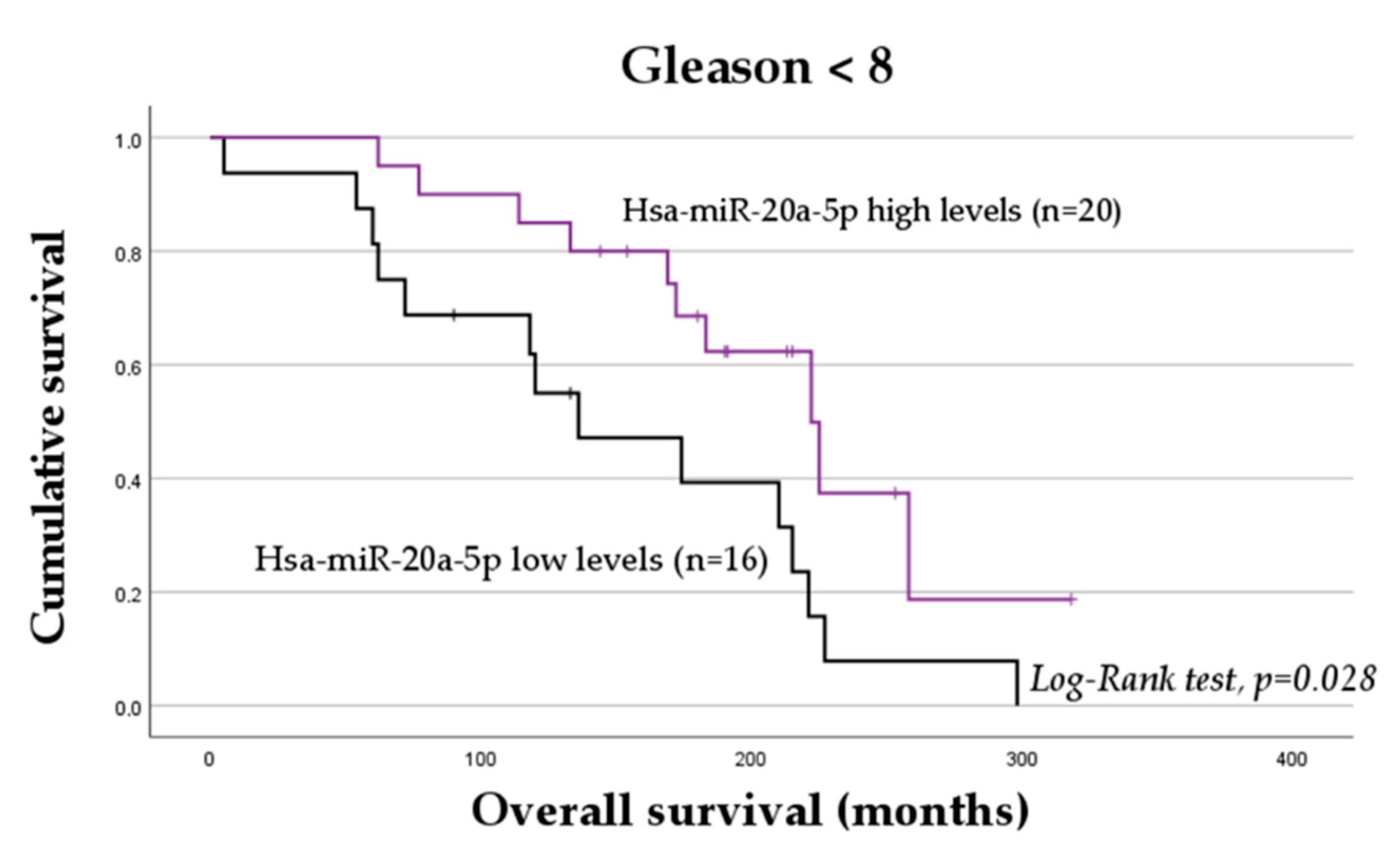
2.4. The Impact of miRNAs on the Overall Survival of mCRPC Patients
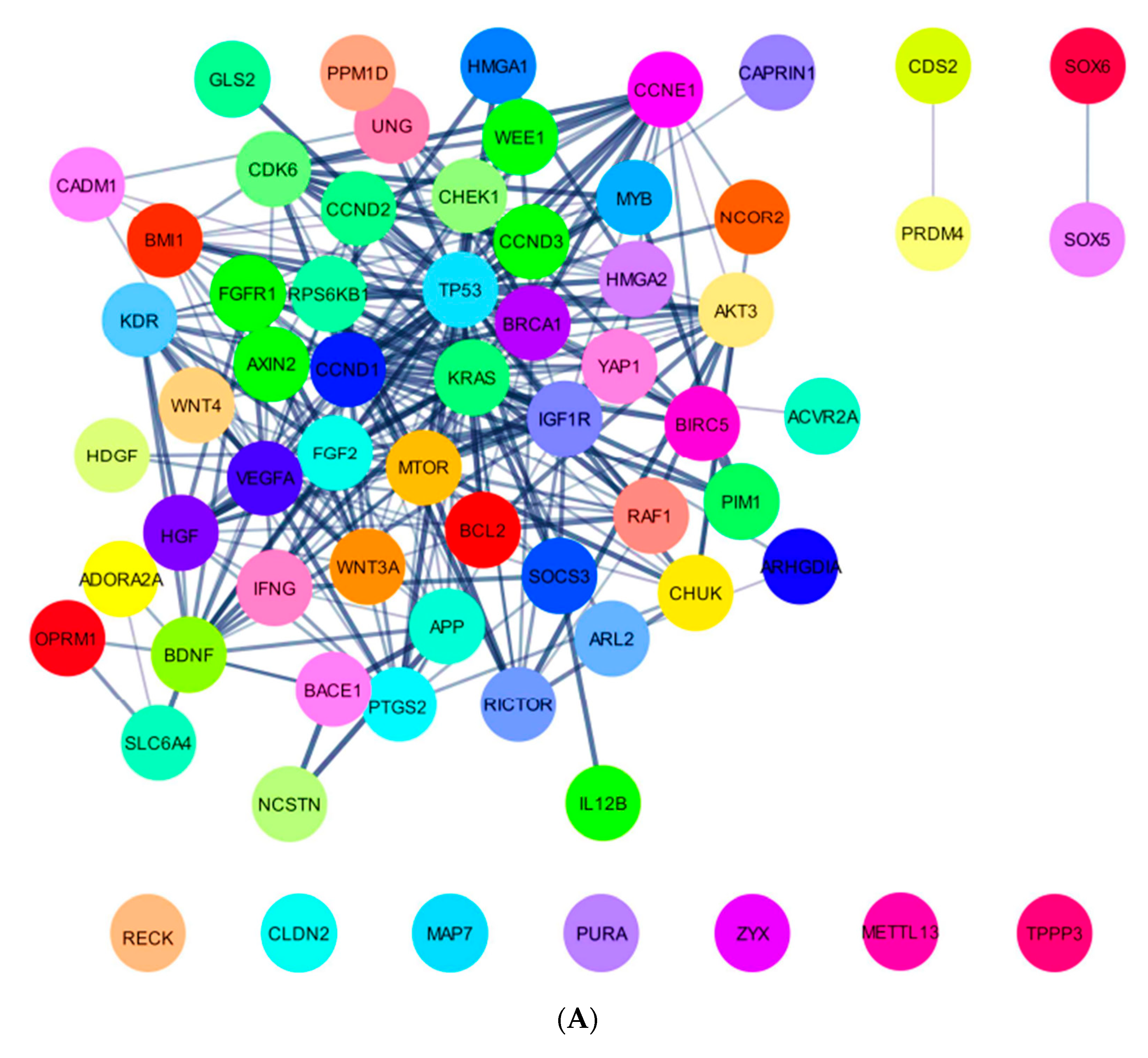
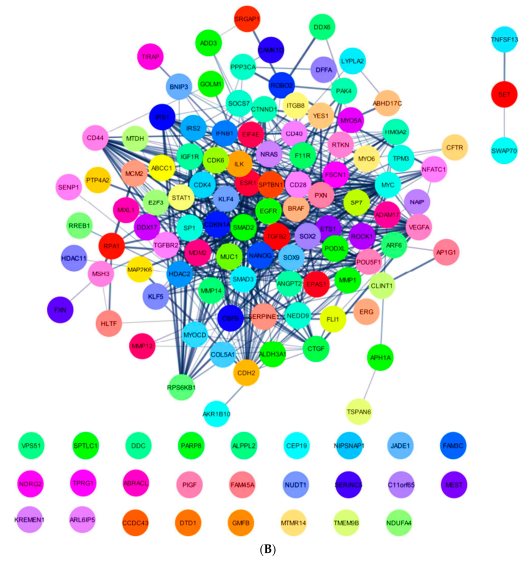
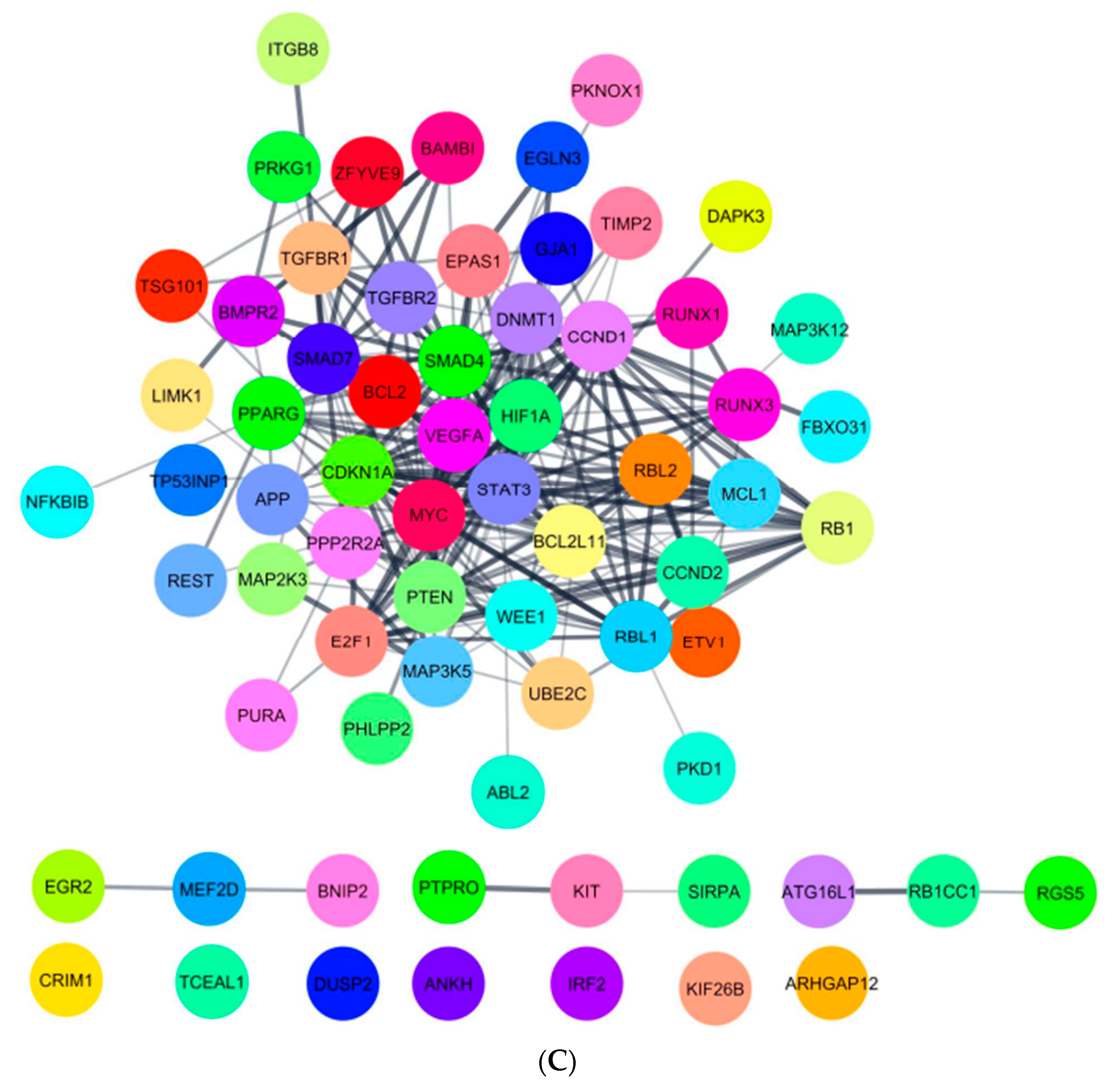
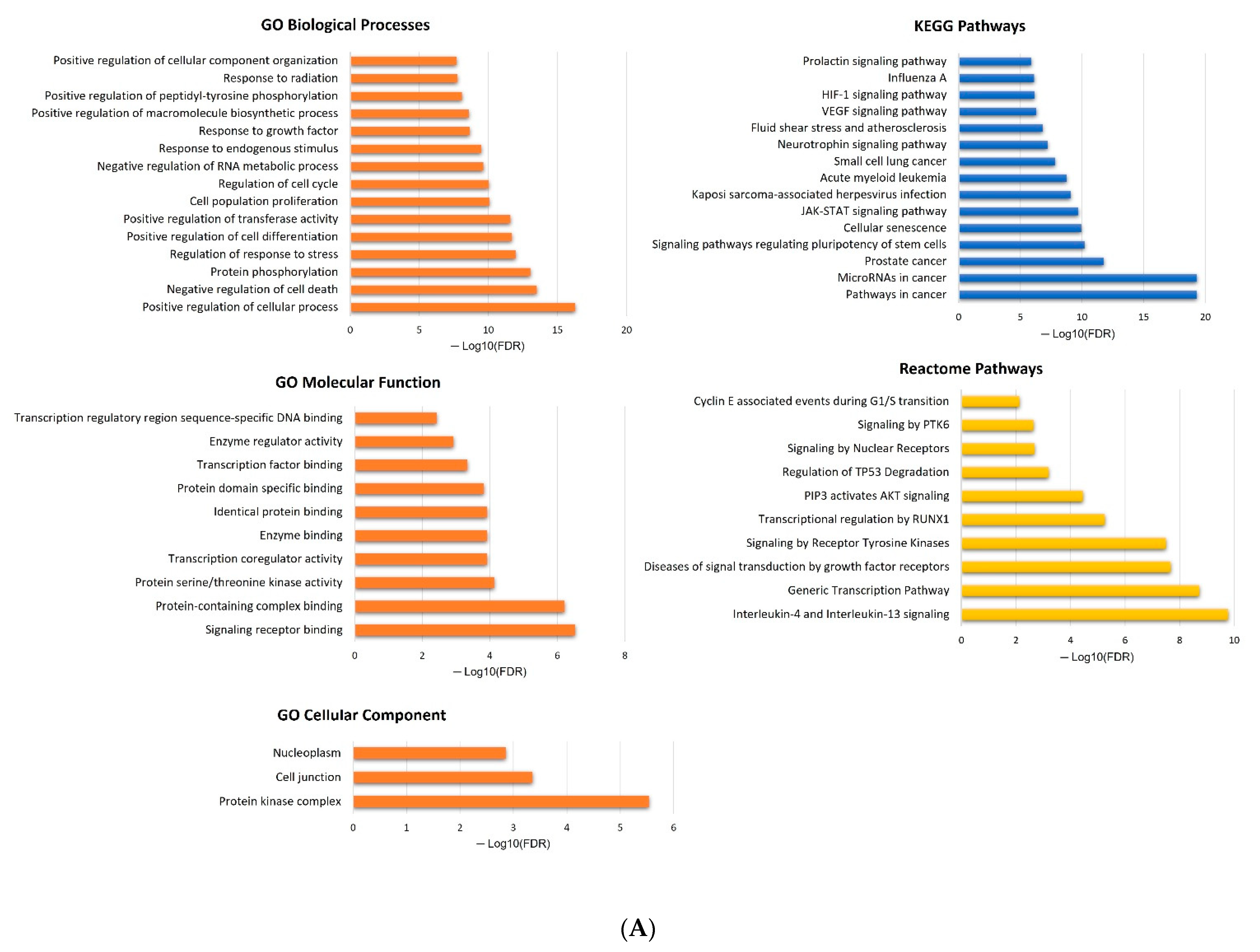
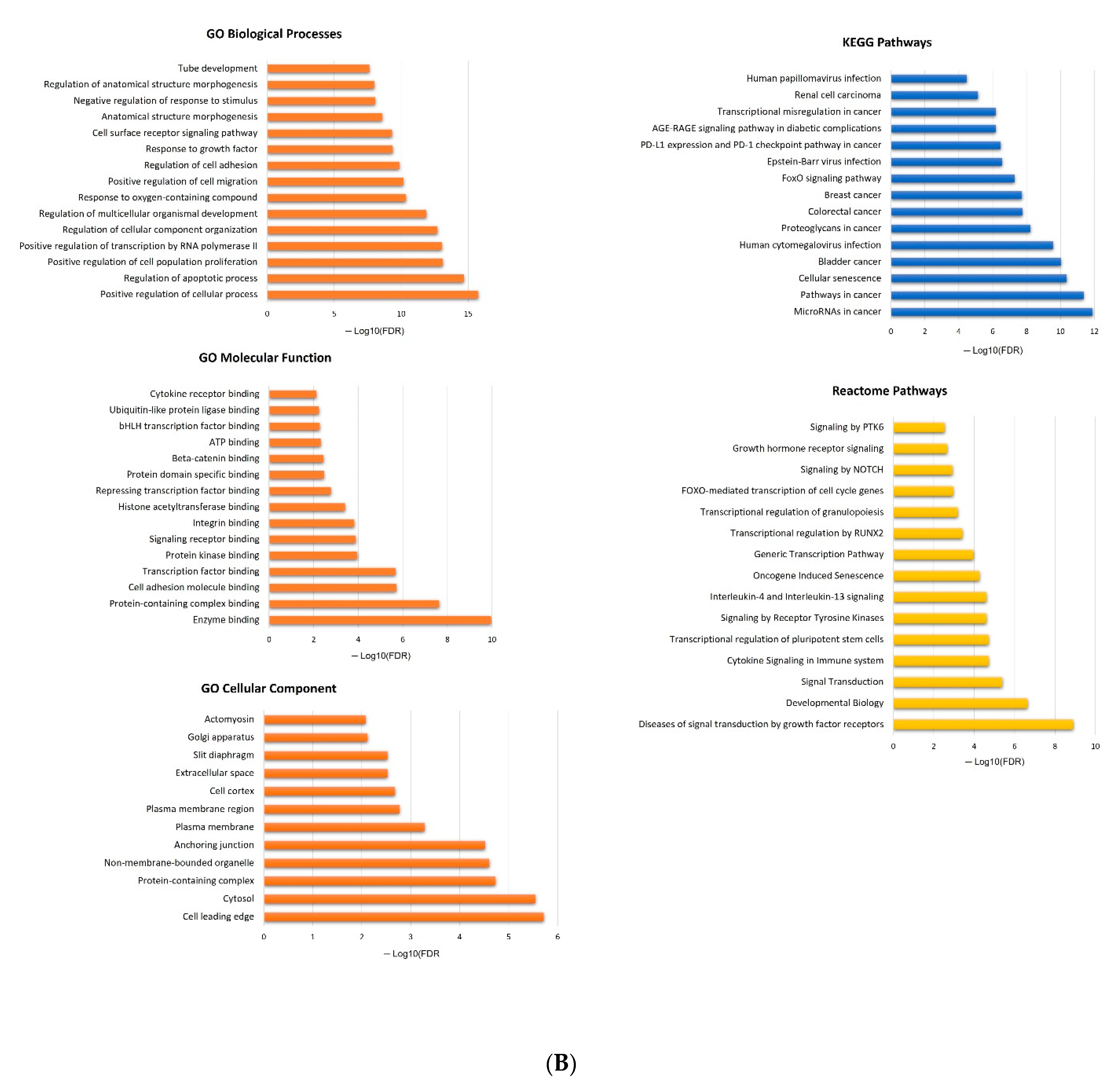
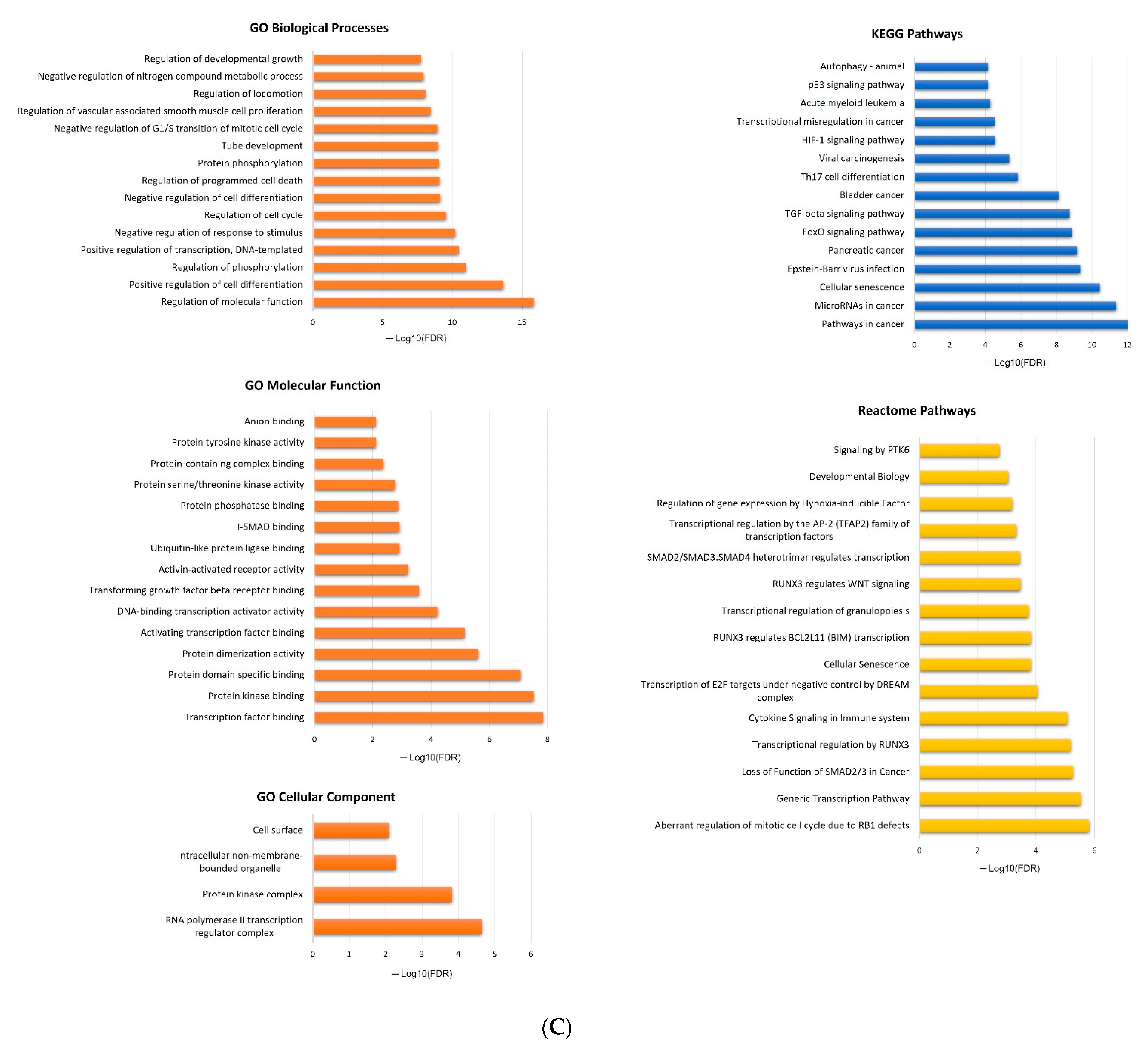
2.5. In Silico Analysis of hsa-miR-16-5p, hsa-miR-145-5p, and hsa-miR-20a-5p Targets
3. Discussion
4. Materials and Methods
4.1. Patients, Controls, and Sample Collection
4.2. miRNA Selection: In Silico Analysis and Literature-Based Approach
4.3. miRNA Extraction, cDNA Synthesis, and miRNA Relative Quantification
4.4. Statistical Analysis
4.5. In Silico Analyses
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Barsouk, A.; Padala, S.A.; Vakiti, A.; Mohammed, A.; Saginala, K.; Thandra, K.C.; Rawla, P.; Barsouk, A. Epidemiology, Staging and Management of Prostate Cancer. Med. Sci. 2020, 8, 28. [Google Scholar] [CrossRef]
- Hou, Z.; Huang, S.; Li, Z. Androgens in prostate cancer: A tale that never ends. Cancer Lett. 2021, 516, 1–12. [Google Scholar] [CrossRef]
- Parker, C.C.; James, N.D.; Brawley, C.D.; Clarke, N.W.; Hoyle, A.P.; Ali, A.; Ritchie, A.W.S.; Attard, G.; Chowdhury, S.; Cross, W.; et al. Radiotherapy to the primary tumour for newly diagnosed, metastatic prostate cancer (STAMPEDE): A randomised controlled phase 3 trial. Lancet 2018, 392, 2353–2366. [Google Scholar] [CrossRef] [PubMed]
- Mansinho, A.; Macedo, D.; Fernandes, I.; Costa, L. Castration-Resistant Prostate Cancer: Mechanisms, Targets and Treatment. Adv. Exp. Med. Biol. 2018, 1096, 117–133. [Google Scholar] [CrossRef]
- Chung, M.S.; Lee, S.H. Side Effects and Management of ADT for Prostate Cancer. Manag. Adv. Prostate Cancer 2018, 149–156. [Google Scholar] [CrossRef]
- Aurilio, G.; Cimadamore, A.; Mazzucchelli, R.; Lopez-Beltran, A.; Verri, E.; Scarpelli, M.; Massari, F.; Cheng, L.; Santoni, M.; Montironi, R. Androgen Receptor Signaling Pathway in Prostate Cancer: From Genetics to Clinical Applications. Cells 2020, 9, 2653. [Google Scholar] [CrossRef]
- Cornford, P.; Bellmunt, J.; Bolla, M.; Briers, E.; De Santis, M.; Gross, T.; Henry, A.M.; Joniau, S.; Lam, T.B.; Mason, M.D.; et al. EAU-ESTRO-SIOG Guidelines on Prostate Cancer. Part II: Treatment of Relapsing, Metastatic, and Castration-Resistant Prostate Cancer. Eur. Urol. 2017, 71, 630–642. [Google Scholar] [CrossRef]
- Kushwaha, P.P.; Gupta, S. New insights for drug resistance in metastatic castration-resistant prostate cancer. Cancer Drug Resist. 2022, 5, 846–849. [Google Scholar] [CrossRef]
- Hussain, A.; Dawson, N. Management of advanced/metastatic prostate cancer: 2000 update. Oncology 2000, 14, discussion 1688, 1691–1674. [Google Scholar]
- Mostaghel, E.A.; Montgomery, B.; Nelson, P.S. Castration-resistant prostate cancer: Targeting androgen metabolic pathways in recurrent disease. Urol. Oncol. 2009, 27, 251–257. [Google Scholar] [CrossRef]
- Seruga, B.; Ocana, A.; Tannock, I.F. Drug resistance in metastatic castration-resistant prostate cancer. Nat. Rev. Clin. Oncol. 2011, 8, 12–23. [Google Scholar] [CrossRef]
- Zedan, A.H.; Osther, P.J.S.; Assenholt, J.; Madsen, J.S.; Hansen, T.F. Circulating miR-141 and miR-375 are associated with treatment outcome in metastatic castration resistant prostate cancer. Sci. Rep. 2020, 10, 227. [Google Scholar] [CrossRef] [PubMed]
- Turco, F.; Gillessen, S.; Cathomas, R.; Buttigliero, C.; Vogl, U.M. Treatment Landscape for Patients with Castration-Resistant Prostate Cancer: Patient Selection and Unmet Clinical Needs. Res. Rep. Urol. 2022, 14, 339–350. [Google Scholar] [CrossRef] [PubMed]
- de Bono, J.S.; Logothetis, C.J.; Molina, A.; Fizazi, K.; North, S.; Chu, L.; Chi, K.N.; Jones, R.J.; Goodman, O.B., Jr.; Saad, F.; et al. Abiraterone and increased survival in metastatic prostate cancer. N. Engl. J. Med. 2011, 364, 1995–2005. [Google Scholar] [CrossRef] [PubMed]
- Scher, H.I.; Fizazi, K.; Saad, F.; Taplin, M.E.; Sternberg, C.N.; Miller, K.; de Wit, R.; Mulders, P.; Chi, K.N.; Shore, N.D.; et al. Increased survival with enzalutamide in prostate cancer after chemotherapy. N. Engl. J. Med. 2012, 367, 1187–1197. [Google Scholar] [CrossRef] [PubMed]
- Ryan, C.J.; Smith, M.R.; de Bono, J.S.; Molina, A.; Logothetis, C.J.; de Souza, P.; Fizazi, K.; Mainwaring, P.; Piulats, J.M.; Ng, S.; et al. Abiraterone in metastatic prostate cancer without previous chemotherapy. N. Engl. J. Med. 2013, 368, 138–148. [Google Scholar] [CrossRef]
- Beer, T.M.; Armstrong, A.J.; Rathkopf, D.E.; Loriot, Y.; Sternberg, C.N.; Higano, C.S.; Iversen, P.; Bhattacharya, S.; Carles, J.; Chowdhury, S.; et al. Enzalutamide in metastatic prostate cancer before chemotherapy. N. Engl. J. Med. 2014, 371, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, G.; Buonerba, L.; Ingenito, C.; Crocetto, F.; Buonerba, C.; Libroia, A.; Sciarra, A.; Ragone, G.; Sanseverino, R.; Iaccarino, S.; et al. Clinical Characteristics of Metastatic Prostate Cancer Patients Infected with COVID-19 in South Italy. Oncology 2020, 98, 743–747. [Google Scholar] [CrossRef] [PubMed]
- Payne, H.; Cornford, P. Prostate-specific antigen: An evolving role in diagnosis, monitoring, and treatment evaluation in prostate cancer. Urol. Oncol. 2011, 29, 593–601. [Google Scholar] [CrossRef]
- Pezaro, C.; Omlin, A.; Lorente, D.; Rodrigues, D.N.; Ferraldeschi, R.; Bianchini, D.; Mukherji, D.; Riisnaes, R.; Altavilla, A.; Crespo, M.; et al. Visceral disease in castration-resistant prostate cancer. Eur. Urol. 2014, 65, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [PubMed]
- Filella, X.; Foj, L. miRNAs as novel biomarkers in the management of prostate cancer. Clin. Chem. Lab. Med. 2017, 55, 715–736. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Volinia, S.; Calin, G.A.; Liu, C.G.; Ambs, S.; Cimmino, A.; Petrocca, F.; Visone, R.; Iorio, M.; Roldo, C.; Ferracin, M.; et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc. Natl. Acad. Sci. USA 2006, 103, 2257–2261. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Bidarra, D.; Constancio, V.; Barros-Silva, D.; Ramalho-Carvalho, J.; Moreira-Barbosa, C.; Antunes, L.; Mauricio, J.; Oliveira, J.; Henrique, R.; Jeronimo, C. Circulating MicroRNAs as Biomarkers for Prostate Cancer Detection and Metastasis Development Prediction. Front. Oncol. 2019, 9, 900. [Google Scholar] [CrossRef]
- Brase, J.C.; Johannes, M.; Schlomm, T.; Falth, M.; Haese, A.; Steuber, T.; Beissbarth, T.; Kuner, R.; Sultmann, H. Circulating miRNAs are correlated with tumor progression in prostate cancer. Int. J. Cancer 2011, 128, 608–616. [Google Scholar] [CrossRef]
- Haldrup, C.; Kosaka, N.; Ochiya, T.; Borre, M.; Hoyer, S.; Orntoft, T.F.; Sorensen, K.D. Profiling of circulating microRNAs for prostate cancer biomarker discovery. Drug Deliv. Transl. Res. 2014, 4, 19–30. [Google Scholar] [CrossRef]
- Saini, S. PSA and beyond: Alternative prostate cancer biomarkers. Cell. Oncol. 2016, 39, 97–106. [Google Scholar] [CrossRef]
- Lone, S.N.; Nisar, S.; Masoodi, T.; Singh, M.; Rizwan, A.; Hashem, S.; El-Rifai, W.; Bedognetti, D.; Batra, S.K.; Haris, M.; et al. Liquid biopsy: A step closer to transform diagnosis, prognosis and future of cancer treatments. Mol. Cancer 2022, 21, 79. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Silveira, D.A.; Mombach, J.C.M. Towards DNA-damage induced autophagy: A Boolean model of p53-induced cell fate mechanisms. DNA Repair 2020, 96, 102971. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yang, S.; Ren, C.; Liu, S.; Zhang, X.; Sui, A. Deciphering the roles of miR-16-5p in malignant solid tumors. Biomed. Pharm. 2022, 148, 112703. [Google Scholar] [CrossRef]
- Krell, A.; Wolter, M.; Stojcheva, N.; Hertler, C.; Liesenberg, F.; Zapatka, M.; Weller, M.; Malzkorn, B.; Reifenberger, G. MiR-16-5p is frequently down-regulated in astrocytic gliomas and modulates glioma cell proliferation, apoptosis and response to cytotoxic therapy. Neuropathol. Appl. Neurobiol. 2019, 45, 441–458. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Ding, F.; Huang, C.; Xiao, H.; Fei, F.; Li, J. Role of miR-16-5p in the proliferation and metastasis of hepatocellular carcinoma. Eur. Rev. Med. Pharm. Sci. 2019, 23, 137–145. [Google Scholar]
- Zhang, S.; Wang, W.; Wu, X.; Liu, W.; Ding, F. miR-16-5p modulates the radiosensitivity of cervical cancer cells via regulating coactivator-associated arginine methyltransferase 1. Pathol. Int. 2020, 70, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Di, X.; Bi, Y.; Sun, S.; Wang, T. Long non-coding RNA LINC00649 regulates YES-associated protein 1 (YAP1)/Hippo pathway to accelerate gastric cancer (GC) progression via sequestering miR-16-5p. Bioengineered 2021, 12, 1791–1802. [Google Scholar] [CrossRef]
- He, J.; Qiu, Z.; Zhang, H.; Gao, Z.; Jiang, Y.; Li, Z.; Kong, C.; Man, X. MicroRNA-16-5p/BIMP1/NF-κB axis regulates autophagy to exert a tumor-suppressive effect on bladder cancer. Mol. Med. Rep. 2021, 24, 576. [Google Scholar] [CrossRef]
- Wang, F.; Wang, W.; Lu, L.; Xie, Y.; Yan, J.; Chen, Y.; Di, C.; Gan, L.; Si, J.; Zhang, H. MicroRNA-16-5p regulates cell survival, cell cycle and apoptosis by targeting AKT3 in prostate cancer cells. Oncol. Rep. 2020, 44, 1282–1292. [Google Scholar] [CrossRef] [PubMed]
- Meireles Da Costa, N.; Palumbo, A.; De Martino, M.; Fusco, A.; Ribeiro Pinto, L.F.; Nasciutti, L.E. Interplay between HMGA and TP53 in cell cycle control along tumor progression. Cell. Mol. Life Sci. 2021, 78, 817–831. [Google Scholar] [CrossRef]
- Chen, X.; Wu, Y.; Wang, X.; Xu, C.; Wang, L.; Jian, J.; Wu, D.; Wu, G. CDK6 is upregulated and may be a potential therapeutic target in enzalutamide-resistant castration-resistant prostate cancer. Eur. J. Med. Res. 2022, 27, 105. [Google Scholar] [CrossRef] [PubMed]
- Cham, J.; Venkateswaran, A.R.; Bhangoo, M. Targeting the PI3K-AKT-mTOR Pathway in Castration Resistant Prostate Cancer: A Review Article. Clin. Genitourin. Cancer 2021, 19, 563.e1–563.e7. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, E.V.; Reece, K.M.; Ley, A.M.; Troutman, S.M.; Sissung, T.M.; Price, D.K.; Chau, C.H.; Figg, W.D. Dual targeting of the androgen receptor and hypoxia-inducible factor 1α pathways synergistically inhibits castration-resistant prostate cancer cells. Mol. Pharmacol. 2015, 87, 1006–1012. [Google Scholar] [CrossRef]
- Tang, W.; Zhang, X.; Tan, W.; Gao, J.; Pan, L.; Ye, X.; Chen, L.; Zheng, W. miR-145-5p suppresses breast cancer progression by inhibiting SOX2. J. Surg. Res. 2019, 236, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Zhao, W.; Chen, Z.; Li, M.; Li, S.; Wu, B.; Bu, R. Circ_0058063 regulates CDK6 to promote bladder cancer progression by sponging miR-145-5p. J. Cell. Physiol. 2019, 234, 4812–4824. [Google Scholar] [CrossRef]
- Zhang, Y.; Wen, X.; Hu, X.; Cheng, L.; Yu, J.; Wei, Z. Downregulation of miR-145-5p correlates with poor prognosis in gastric cancer. Eur. Rev. Med. Pharm. Sci. 2016, 20, 3026–3030. [Google Scholar]
- Zhang, X.; Wu, J. Prognostic role of microRNA-145 in prostate cancer: A systems review and meta-analysis. Prostate Int. 2015, 3, 71–74. [Google Scholar] [CrossRef]
- Avgeris, M.; Stravodimos, K.; Fragoulis, E.G.; Scorilas, A. The loss of the tumour-suppressor miR-145 results in the shorter disease-free survival of prostate cancer patients. Br. J. Cancer 2013, 108, 2573–2581. [Google Scholar] [CrossRef]
- Fuse, M.; Nohata, N.; Kojima, S.; Sakamoto, S.; Chiyomaru, T.; Kawakami, K.; Enokida, H.; Nakagawa, M.; Naya, Y.; Ichikawa, T.; et al. Restoration of miR-145 expression suppresses cell proliferation, migration and invasion in prostate cancer by targeting FSCN1. Int. J. Oncol. 2011, 38, 1093–1101. [Google Scholar] [CrossRef]
- Capik, O.; Sanli, F.; Kurt, A.; Ceylan, O.; Suer, I.; Kaya, M.; Ittmann, M.; Karatas, O.F. CASC11 promotes aggressiveness of prostate cancer cells through miR-145/IGF1R axis. Prostate Cancer Prostatic Dis. 2021, 24, 891–902. [Google Scholar] [CrossRef]
- Gaur, S.; Gallick, G. Tumor suppressive miRNA-145 inhibits IGF-1 and cell viability in prostate cancer cells. Cancer Res. 2014, 74, 4356. [Google Scholar] [CrossRef]
- Zeinali, T.; Mansoori, B.; Mohammadi, A.; Baradaran, B. Regulatory mechanisms of miR-145 expression and the importance of its function in cancer metastasis. Biomed. Pharm. 2019, 109, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Wu, X.; Xiang, S.; Qiao, M.; Cen, X.; Pan, X.; Huang, X.; Zhao, Z. Regulatory mechanism of miR-20a-5p expression in Cancer. Cell Death Discov. 2022, 8, 262. [Google Scholar] [CrossRef] [PubMed]
- Sylvestre, Y.; De Guire, V.; Querido, E.; Mukhopadhyay, U.K.; Bourdeau, V.; Major, F.; Ferbeyre, G.; Chartrand, P. An E2F/miR-20a autoregulatory feedback loop. J. Biol. Chem. 2007, 282, 2135–2143. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Geng, D.; Li, S.; Chen, Z.; Sun, M. LncRNA HOTAIR influences cell growth, migration, invasion, and apoptosis via the miR-20a-5p/HMGA2 axis in breast cancer. Cancer Med. 2018, 7, 842–855. [Google Scholar] [CrossRef]
- Qin, X.; Zhou, M.; Lv, H.; Mao, X.; Li, X.; Guo, H.; Li, L.; Xing, H. Long noncoding RNA LINC00657 inhibits cervical cancer development by sponging miR-20a-5p and targeting RUNX3. Cancer Lett. 2021, 498, 130–141. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Ma, H.; Wang, J.; Kang, Y.; Xue, Q. miR-20a-5p inhibits endometrial cancer progression by targeting janus kinase 1. Oncol. Lett. 2021, 21, 427. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Han, J.; Chen, J.; Dong, J.; Xia, Y.; Liu, J.; Jiang, Y.; Dai, J.; Lu, J.; Jin, G.; et al. Plasma miRNAs as early biomarkers for detecting hepatocellular carcinoma. Int. J. Cancer 2015, 137, 1679–1690. [Google Scholar] [CrossRef]
- Daniel, R.; Wu, Q.; Williams, V.; Clark, G.; Guruli, G.; Zehner, Z. A Panel of MicroRNAs as Diagnostic Biomarkers for the Identification of Prostate Cancer. Int. J. Mol. Sci. 2017, 18, 1281. [Google Scholar] [CrossRef]
- Liu, H.; Wu, Z.; Zhou, H.; Cai, W.; Li, X.; Hu, J.; Gao, L.; Feng, T.; Wang, L.; Peng, X.; et al. The SOX4/miR-17-92/RB1 Axis Promotes Prostate Cancer Progression. Neoplasia 2019, 21, 765–776. [Google Scholar] [CrossRef]
- Li, J.; Ye, D.; Shen, P.; Liu, X.; Zhou, P.; Zhu, G.; Xu, Y.; Fu, Y.; Li, X.; Sun, J.; et al. Mir-20a-5p induced WTX deficiency promotes gastric cancer progressions through regulating PI3K/AKT signaling pathway. J. Exp. Clin. Cancer Res. 2020, 39, 212. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhou, D.; Li, S.; Feng, Y.; Li, X.; Chang, W.; Zhang, J.; Sun, Y.; Qing, D.; Chen, G.; et al. Licochalcone A reverses NNK-induced ectopic miRNA expression to elicit in vitro and in vivo chemopreventive effects. Phytomed. Int. J. Phytother. Phytopharm. 2020, 76, 153245. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Qie, J.; Fu, Q.; Chen, J.; Jin, Y.; Ding, Z. miR-20a-5p/TGFBR2 Axis Affects Pro-inflammatory Macrophages and Aggravates Liver Fibrosis. Front. Oncol. 2020, 10, 107. [Google Scholar] [CrossRef]
- Zhu, M.L.; Kyprianou, N. Androgen receptor and growth factor signaling cross-talk in prostate cancer cells. Endocr. Relat. Cancer 2008, 15, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.L.; Partin, J.V.; Bruckheimer, E.M.; Strup, S.E.; Kyprianou, N. TGF-beta signaling and androgen receptor status determine apoptotic cross-talk in human prostate cancer cells. Prostate 2008, 68, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.B.; Xue, L.; Yang, J.; Ma, A.H.; Zhao, J.; Xu, M.; Tepper, C.G.; Evans, C.P.; Kung, H.J.; deVere White, R.W. An androgen-regulated miRNA suppresses Bak1 expression and induces androgen-independent growth of prostate cancer cells. Proc. Natl. Acad. Sci. USA 2007, 104, 19983–19988. [Google Scholar] [CrossRef]
- Shi, X.B.; Xue, L.; Ma, A.H.; Tepper, C.G.; Kung, H.J.; White, R.W. miR-125b promotes growth of prostate cancer xenograft tumor through targeting pro-apoptotic genes. Prostate 2011, 71, 538–549. [Google Scholar] [CrossRef]
- Boll, K.; Reiche, K.; Kasack, K.; Morbt, N.; Kretzschmar, A.K.; Tomm, J.M.; Verhaegh, G.; Schalken, J.; von Bergen, M.; Horn, F.; et al. MiR-130a, miR-203 and miR-205 jointly repress key oncogenic pathways and are downregulated in prostate carcinoma. Oncogene 2013, 32, 277–285. [Google Scholar] [CrossRef]
- Daniunaite, K.; Dubikaityte, M.; Gibas, P.; Bakavicius, A.; Rimantas Lazutka, J.; Ulys, A.; Jankevicius, F.; Jarmalaite, S. Clinical significance of miRNA host gene promoter methylation in prostate cancer. Hum. Mol. Genet. 2017, 26, 2451–2461. [Google Scholar] [CrossRef]
- Sato, S.; Katsushima, K.; Shinjo, K.; Hatanaka, A.; Ohka, F.; Suzuki, S.; Naiki-Ito, A.; Soga, N.; Takahashi, S.; Kondo, Y. Histone Deacetylase Inhibition in Prostate Cancer Triggers miR-320-Mediated Suppression of the Androgen Receptor. Cancer Res. 2016, 76, 4192–4204. [Google Scholar] [CrossRef]
- Chen, X.; Xu, X.; Pan, B.; Zeng, K.; Xu, M.; Liu, X.; He, B.; Pan, Y.; Sun, H.; Wang, S. miR-150-5p suppresses tumor progression by targeting VEGFA in colorectal cancer. Aging 2018, 10, 3421–3437. [Google Scholar] [CrossRef] [PubMed]
- Sugita, B.M.; Rodriguez, Y.; Fonseca, A.S.; Nunes Souza, E.; Kallakury, B.; Cavalli, I.J.; Ribeiro, E.; Aneja, R.; Cavalli, L.R. MiR-150-5p Overexpression in Triple-Negative Breast Cancer Contributes to the In Vitro Aggressiveness of This Breast Cancer Subtype. Cancers 2022, 14, 2156. [Google Scholar] [CrossRef] [PubMed]
- Ostling, P.; Leivonen, S.K.; Aakula, A.; Kohonen, P.; Makela, R.; Hagman, Z.; Edsjo, A.; Kangaspeska, S.; Edgren, H.; Nicorici, D.; et al. Systematic analysis of microRNAs targeting the androgen receptor in prostate cancer cells. Cancer Res. 2011, 71, 1956–1967. [Google Scholar] [CrossRef] [PubMed]
- Rokhlin, O.W.; Scheinker, V.S.; Taghiyev, A.F.; Bumcrot, D.; Glover, R.A.; Cohen, M.B. MicroRNA-34 mediates AR-dependent p53-induced apoptosis in prostate cancer. Cancer Biol. 2008, 7, 1288–1296. [Google Scholar] [CrossRef] [PubMed]
- Afonso, A.; Silva, J.; Lopes, A.R.; Coelho, S.; Patrao, A.S.; Rosinha, A.; Carneiro, F.; Pinto, A.R.; Mauricio, M.J.; Medeiros, R. YB-1 variant and androgen receptor axis-targeted agents in metastatic castration-resistant prostate cancer patients. Pharmacogenomics 2020, 21, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Buyyounouski, M.K.; Choyke, P.L.; McKenney, J.K.; Sartor, O.; Sandler, H.M.; Amin, M.B.; Kattan, M.W.; Lin, D.W. Prostate cancer—Major changes in the American Joint Committee on Cancer eighth edition cancer staging manual. CA Cancer J. Clin. 2017, 67, 245–253. [Google Scholar] [CrossRef]
- Glare, P.; Sinclair, C.; Downing, M.; Stone, P.; Maltoni, M.; Vigano, A. Predicting survival in patients with advanced disease. Eur. J. Cancer 2008, 44, 1146–1156. [Google Scholar] [CrossRef]
- Dewys, W.D.; Begg, C.; Lavin, P.T.; Band, P.R.; Bennett, J.M.; Bertino, J.R.; Cohen, M.H.; Douglass, H.O., Jr.; Engstrom, P.F.; Ezdinli, E.Z.; et al. Prognostic effect of weight loss prior to chemotherapy in cancer patients. Eastern Cooperative Oncology Group. Am. J. Med. 1980, 69, 491–497. [Google Scholar] [CrossRef]
- D’Amico, A.V.; Whittington, R.; Malkowicz, S.B.; Schultz, D.; Blank, K.; Broderick, G.A.; Tomaszewski, J.E.; Renshaw, A.A.; Kaplan, I.; Beard, C.J.; et al. Biochemical outcome after radical prostatectomy, external beam radiation therapy, or interstitial radiation therapy for clinically localized prostate cancer. JAMA J. Am. Med. Assoc. 1998, 280, 969–974. [Google Scholar] [CrossRef]
- Kern, F.; Aparicio-Puerta, E.; Li, Y.; Fehlmann, T.; Kehl, T.; Wagner, V.; Ray, K.; Ludwig, N.; Lenhof, H.P.; Meese, E.; et al. miRTargetLink 2.0-interactive miRNA target gene and target pathway networks. Nucleic Acids Res. 2021, 49, W409–W416. [Google Scholar] [CrossRef]
- Wang, F.; Mao, A.; Tang, J.; Zhang, Q.; Yan, J.; Wang, Y.; Di, C.; Gan, L.; Sun, C.; Zhang, H. microRNA-16-5p enhances radiosensitivity through modulating Cyclin D1/E1-pRb-E2F1 pathway in prostate cancer cells. J. Cell. Physiol. 2019, 234, 13182–13190. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, M.A.; Kreger, B.; Omaruddin, R.A. Transcriptional modulation of micro-RNA in human cells differing in radiation sensitivity. Int. J. Radiat. Biol. 2010, 86, 569–583. [Google Scholar] [CrossRef] [PubMed]
- Bonci, D.; De Maria, R. miR-15/miR-16 loss, miR-21 upregulation, or deregulation of their target genes predicts poor prognosis in prostate cancer patients. Mol. Cell. Oncol. 2016, 3, e1109744. [Google Scholar] [CrossRef]
- Stoen, M.J.; Andersen, S.; Rakaee, M.; Pedersen, M.I.; Ingebriktsen, L.M.; Donnem, T.; Lombardi, A.P.G.; Kilvaer, T.K.; Busund, L.R.; Richardsen, E. Overexpression of miR-20a-5p in Tumor Epithelium Is an Independent Negative Prognostic Indicator in Prostate Cancer-A Multi-Institutional Study. Cancers 2021, 13, 4096. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.M.; Castillo, L.; Mahon, K.L.; Chiam, K.; Lee, B.Y.; Nguyen, Q.; Boyer, M.J.; Stockler, M.R.; Pavlakis, N.; Marx, G.; et al. Circulating microRNAs are associated with docetaxel chemotherapy outcome in castration-resistant prostate cancer. Br. J. Cancer 2014, 110, 2462–2471. [Google Scholar] [CrossRef]
- Yamamura, S.; Saini, S.; Majid, S.; Hirata, H.; Ueno, K.; Deng, G.; Dahiya, R. MicroRNA-34a modulates c-Myc transcriptional complexes to suppress malignancy in human prostate cancer cells. PLoS ONE 2012, 7, e29722. [Google Scholar] [CrossRef]
- Kojima, K.; Fujita, Y.; Nozawa, Y.; Deguchi, T.; Ito, M. MiR-34a attenuates paclitaxel-resistance of hormone-refractory prostate cancer PC3 cells through direct and indirect mechanisms. Prostate 2010, 70, 1501–1512. [Google Scholar] [CrossRef]
- Liu, C.; Kelnar, K.; Liu, B.; Chen, X.; Calhoun-Davis, T.; Li, H.; Patrawala, L.; Yan, H.; Jeter, C.; Honorio, S.; et al. The microRNA miR-34a inhibits prostate cancer stem cells and metastasis by directly repressing CD44. Nat. Med. 2011, 17, 211–215. [Google Scholar] [CrossRef]
- Damodaran, M.; Chinambedu Dandapani, M.; SimonDuraiRaj; SandhyaSundaram; VenkatRamanan, S.; Ramachandran, I.; Venkatesan, V. Differentially expressed miR-20, miR-21, miR-100, miR-125a and miR-146a as a potential biomarker for prostate cancer. Mol. Biol. Rep. 2021, 48, 3349–3356. [Google Scholar] [CrossRef]
- Mu, H.Q.; He, Y.H.; Wang, S.B.; Yang, S.; Wang, Y.J.; Nan, C.J.; Bao, Y.F.; Xie, Q.P.; Chen, Y.H. MiR-130b/TNF-alpha/NF-kappaB/VEGFA loop inhibits prostate cancer angiogenesis. Clin. Transl. Oncol. 2020, 22, 111–121. [Google Scholar] [CrossRef]
- Fort, R.S.; Matho, C.; Oliveira-Rizzo, C.; Garat, B.; Sotelo-Silveira, J.R.; Duhagon, M.A. An integrated view of the role of miR-130b/301b miRNA cluster in prostate cancer. Exp. Hematol. Oncol. 2018, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Ramalho-Carvalho, J.; Martins, J.B.; Cekaite, L.; Sveen, A.; Torres-Ferreira, J.; Graca, I.; Costa-Pinheiro, P.; Eilertsen, I.A.; Antunes, L.; Oliveira, J.; et al. Epigenetic disruption of miR-130a promotes prostate cancer by targeting SEC23B and DEPDC1. Cancer Lett. 2017, 385, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Larne, O.; Hagman, Z.; Lilja, H.; Bjartell, A.; Edsjo, A.; Ceder, Y. miR-145 suppress the androgen receptor in prostate cancer cells and correlates to prostate cancer prognosis. Carcinogenesis 2015, 36, 858–866. [Google Scholar] [CrossRef]
- Guo, T.; Wang, Y.; Jia, J.; Mao, X.; Stankiewicz, E.; Scandura, G.; Burke, E.; Xu, L.; Marzec, J.; Davies, C.R.; et al. The Identification of Plasma Exosomal miR-423-3p as a Potential Predictive Biomarker for Prostate Cancer Castration-Resistance Development by Plasma Exosomal miRNA Sequencing. Front. Cell Dev. Biol. 2020, 8, 602493. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Di Wang, X. miR-150-5p represses TP53 tumor suppressor gene to promote proliferation of colon adenocarcinoma. Sci. Rep. 2019, 9, 6740. [Google Scholar] [CrossRef]
- Alekseev, B.; Knyazev, E.; Shkurnikov, M.; Mikhailenko, D.; Zotikov, A.; Nyushko, K.; Tonevitskiy, A.; Kaprin, A. Mp28-10 panel of 6 micrornas for minimally invasive diagnosis of prostate cancer. J. Urol. 2017, 197, e341. [Google Scholar] [CrossRef]
- Costa, A.C.; Santos, J.M.O.; Medeiros-Fonseca, B.; Oliveira, P.A.; Bastos, M.; Brito, H.O.; Gil da Costa, R.M.; Medeiros, R. Characterizing the Inflammatory Microenvironment in K14-HPV16 Transgenic Mice: Mast Cell Infiltration and MicroRNA Expression. Cancers 2022, 14, 2216. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Acunzo, M.; Romano, G.; Wernicke, D.; Croce, C.M. MicroRNA and cancer—A brief overview. Adv. Biol. Regul. 2015, 57, 1–9. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | aHR (95% CI) | p-Value | aHR (95% CI) | p-Value |
|---|---|---|---|---|
| Entire cohort (n = 55) | Initial model | Final model ** | ||
| Age at disease diagnosis (≥64 vs. <64 years *) | 1.59 (0.75–3.37) | 0.229 | - | - |
| Metastasis at disease diagnosis (Yes vs. no *) | 0.65 (0.30–1.38) | 0.259 | - | - |
| PSA at disease diagnosis (>20 vs. ≤20 ng/mL *) | 0.66 (0.30–1.43) | 0.292 | - | - |
| Gleason score (≥8 vs. <8 *) | 2.60 (1.14–5.95) | 0.024 | 1.75 (0.94–3.27) | 0.080 |
| Age at ARAT initiation (≥76 vs. <76 years *) | 0.75 (0.33–1.72) | 0.496 | - | - |
| ECOG at ARAT agent initiation (2 vs. 0/1 *) | 6.48 (1.94–21.62) | 0.002 | 4.09 (1.30–12.87) | 0.016 |
| Indication for ARAT agent (After vs. before docetaxel *) | 3.15 (1.59–6.25) | <0.001 | 2.93 (1.51–5.69) | 0.001 |
| ARAT agent (ENZ vs. AbA *) | 0.74 (0.36–1.54) | 0.425 | - | - |
| Patients under AbA (n = 19) | Initial model | Final model ** | ||
| Gleason score (≥8 vs. <8 *) | 1.65 (0.45–6.12) | 0.451 | - | - |
| ECOG at ARAT agent initiation (2 vs. 0/1 *) | NA | NA | - | - |
| Indication for ARAT agent (After vs. before docetaxel *) | 1.00 (0.28–3.57) | 0.995 | - | - |
| Hsa-miR-16-5p (Low vs. high levels *) | 8.99 (2.09–38.71) | 0.003 | 5.58 (1.54–20.28) | 0.009 |
| Hsa-miR-125b-5p (Low vs. high levels *) | 0.32 (0.07–1.45) | 0.140 | - | - |
| Hsa-miR-130b-3p (Low vs. high levels *) | 4.09 (0.46–36.79) | 0.208 | - | - |
| Hsa-miR-145-5p (Low vs. high levels *) | 3.36 (0.40–28.52) | 0.267 | 4.71 (1.18–18.88) | 0.029 |
| Factor | aHR (95% CI) | p-Value | aHR (95% CI) | p-Value |
|---|---|---|---|---|
| Entire cohort (n = 64) | Initial model | Final model ** | ||
| Age at disease diagnosis (≥64 vs. <64 years *) | 8.82 (3.11–25.01) | <0.001 | 7.56 (3.18–18.01) | <0.001 |
| Metastasis at disease diagnosis (Yes vs. no *) | 2.24 (0.79–6.34) | 0.128 | - | - |
| PSA at disease diagnosis (>20 vs. ≤20 ng/mL *) | 0.59 (0.25–1.37) | 0.217 | - | - |
| Gleason score (≥8 vs. <8 *) | 2.76 (1.21–6.29) | 0.016 | 2.66 (1.33–5.32) | 0.006 |
| Age at ARAT initiation (≥76 vs. <76 year s *) | 0.77 (0.34–1.77) | 0.537 | - | - |
| ECOG at ARAT agent initiation (2 vs. 0/1 *) | 5.81 (1.35–25.01) | 0.018 | 5.73 (1.47–22.28) | 0.012 |
| Indication for ARAT agent (After vs. before docetaxel *) | 4.20 (1.87–9.46) | <0.001 | 4.50 (2.03–9.98) | <0.001 |
| ARAT agent (ENZ vs. AbA *) | 0.94 (0.45–1.99) | 0.873 | - | - |
| Entire cohort (n = 70) | Initial model | Final model ** | ||
| Age at disease diagnosis (≥64 vs. <64 years *) | 5.70 (2.60–12.50) | <0.001 | NA | NA |
| Gleason score (≥8 vs. <8 *) | 2.49 (1.29–4.78) | 0.006 | NA | NA |
| ECOG at ARAT agent initiation (2 vs. 0/1 *) | 9.37 (2.35–37.43) | 0.002 | NA | NA |
| Indication for ARAT agent (After vs. before Docetaxel *) | 4.89 (2.32–10.30) | <0.001 | NA | NA |
| Hsa-miR-20a-5p (Low vs. high levels) | 2.48 (1.28–4.83) | 0.007 | NA | NA |
| MiRNA | MiRNA Genomic Location | Potential Targets | Source | References |
|---|---|---|---|---|
| Hsa-miR-16-5p | 3q25.33 13q14.2 | CCND1 | MIRT001225 | [81,82,83] |
| BCL2 | MIRT001800 | |||
| VEGFA | MIRT003890 | |||
| TP53 | MIRT005764 | |||
| Hsa-miR-20a-5p | 13q31.3 | CCND1 | MIRT000179 | [84,85] |
| BCL2 | MIRT003011 | |||
| PTEN | MIRT003369 | |||
| VEGFA | MIRT004450 | |||
| MYC | MIRT005289 | |||
| STAT3 | MIRT050559 | |||
| Hsa-miR-34a-5p | 1p36.22 | MYC | MIRT000695 | [73,86,87,88] |
| CCND1 | MIRT001013 | |||
| BCL2 | MIRT002298 | |||
| TP53 | MIRT007112 | |||
| AKT1 | MIRT733152 | |||
| Hsa-miR-125b-5p | 11q24.1 21q21.1 | TP53 | MIRT000535 | [66,67,89] |
| AKT1 | MIRT004363 | |||
| STAT3 | MIRT005006 | |||
| BCL2 | MIRT006253 | |||
| EGFR | MIRT733343 | |||
| Hsa-miR-130b-3p | 22q11.21 | STAT3 | MIRT053071 | [68,90,91,92] |
| PTEN | MIRT054460 | |||
| Hsa-miR-145-5p | 5q32 | EGFR | MIRT003325 | [48,49,93] |
| MYC | MIRT004290 | |||
| VEGFA | MIRT006215 | |||
| Hsa-miR-150-5p | 19q13.33 | VEGFA | MIRT004272 | [71,72,94,95] |
| TP53 | MIRT052652 | |||
| Hsa-miR-155-5p | 21q21.3 | CCND1 | MIRT020946 | [69,96] |
| MYC | MIRT054028 | |||
| PTEN | MIRT734893 | |||
| Hsa-miR-320a | 8p21.3 | MYC | MIRT044759 | [70,94] |
| PTEN | MIRT438486 | |||
| VEGFA | MIRT732583 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, J.; Tavares, V.; Afonso, A.; Garcia, J.; Cerqueira, F.; Medeiros, R. Plasmatic MicroRNAs and Treatment Outcomes of Patients with Metastatic Castration-Resistant Prostate Cancer: A Hospital-Based Cohort Study and In Silico Analysis. Int. J. Mol. Sci. 2023, 24, 9101. https://doi.org/10.3390/ijms24109101
Silva J, Tavares V, Afonso A, Garcia J, Cerqueira F, Medeiros R. Plasmatic MicroRNAs and Treatment Outcomes of Patients with Metastatic Castration-Resistant Prostate Cancer: A Hospital-Based Cohort Study and In Silico Analysis. International Journal of Molecular Sciences. 2023; 24(10):9101. https://doi.org/10.3390/ijms24109101
Chicago/Turabian StyleSilva, Jani, Valéria Tavares, Ana Afonso, Juliana Garcia, Fátima Cerqueira, and Rui Medeiros. 2023. "Plasmatic MicroRNAs and Treatment Outcomes of Patients with Metastatic Castration-Resistant Prostate Cancer: A Hospital-Based Cohort Study and In Silico Analysis" International Journal of Molecular Sciences 24, no. 10: 9101. https://doi.org/10.3390/ijms24109101