
Modulation of Membrane Trafficking of AQP5 in the Lens in Response to Changes in Zonular Tension Is Mediated by the Mechanosensitive Channel TRPV1

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

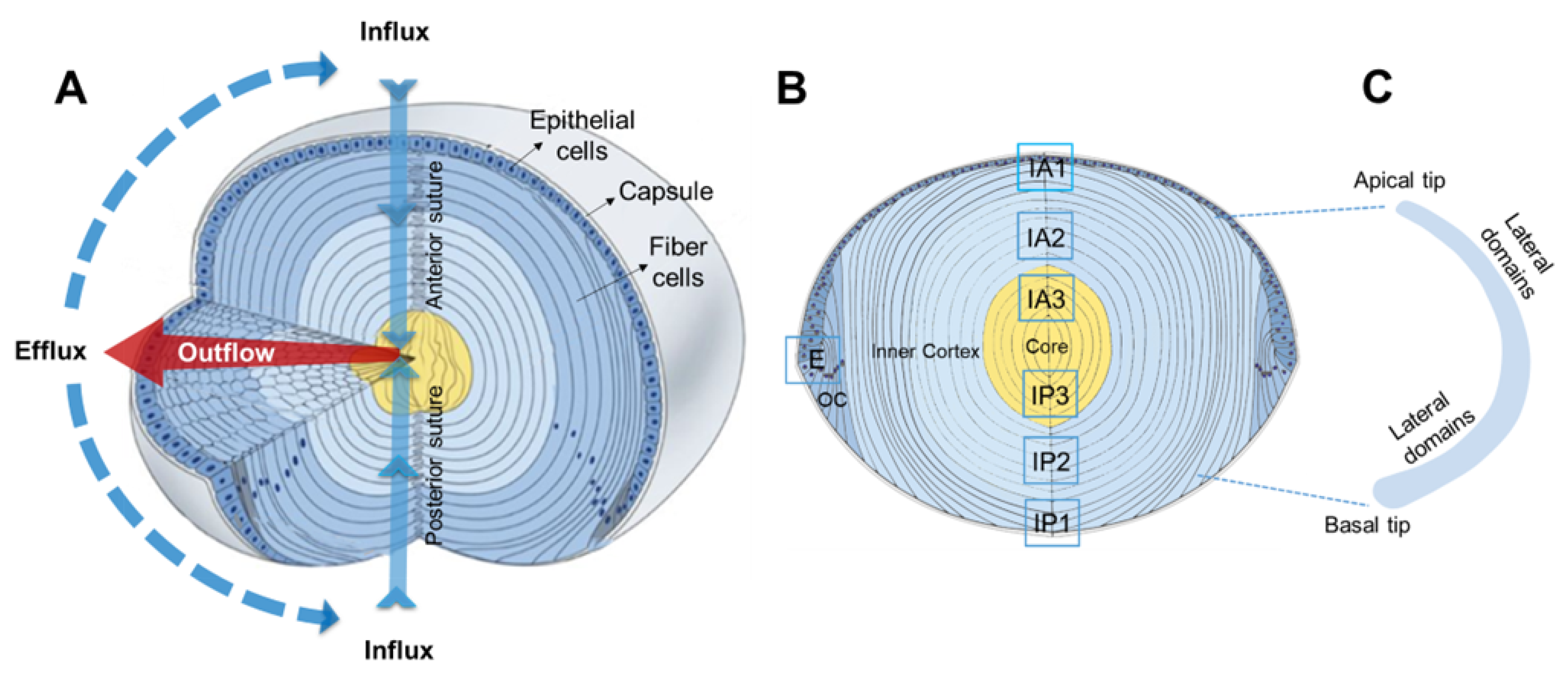
2. Results
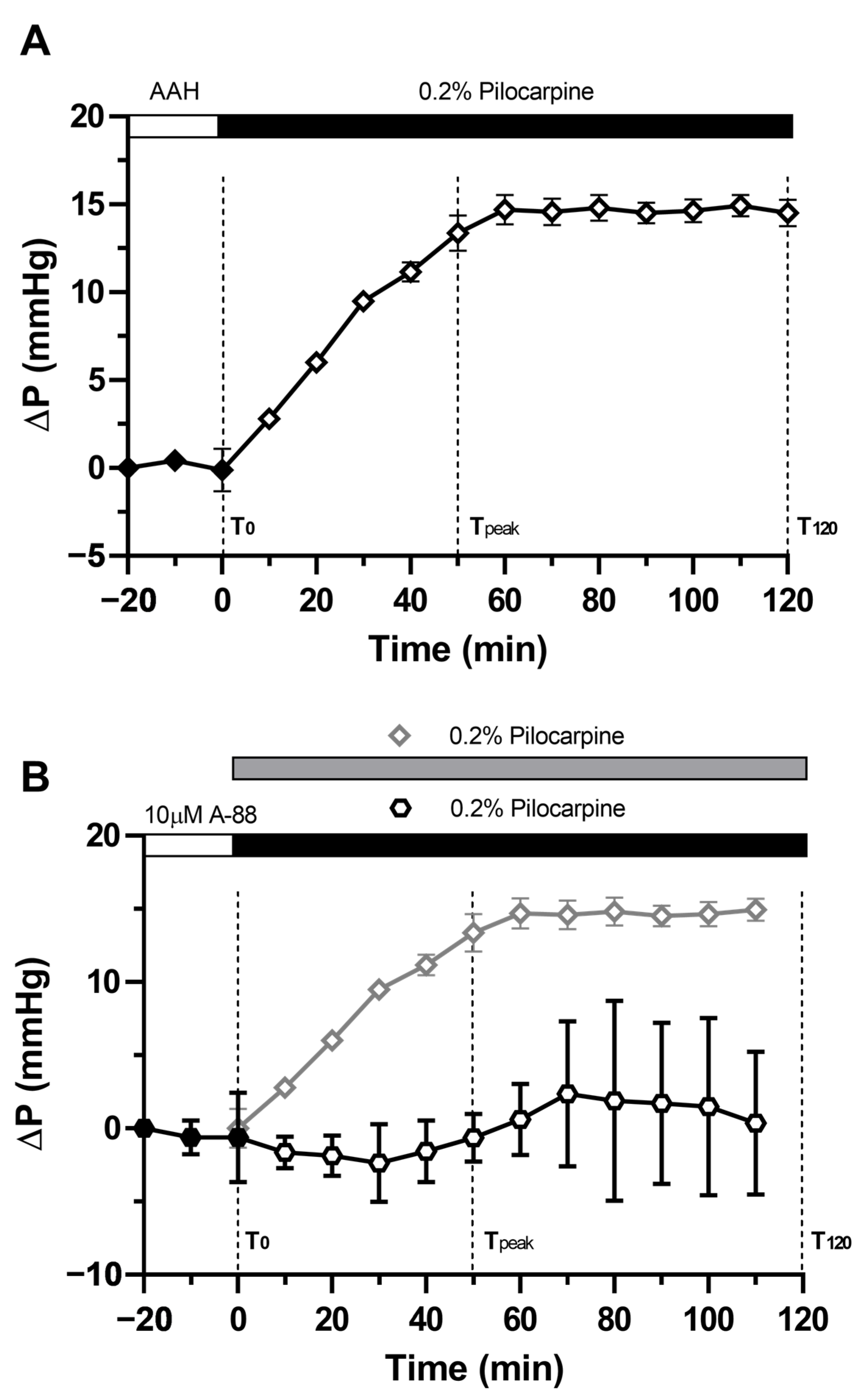
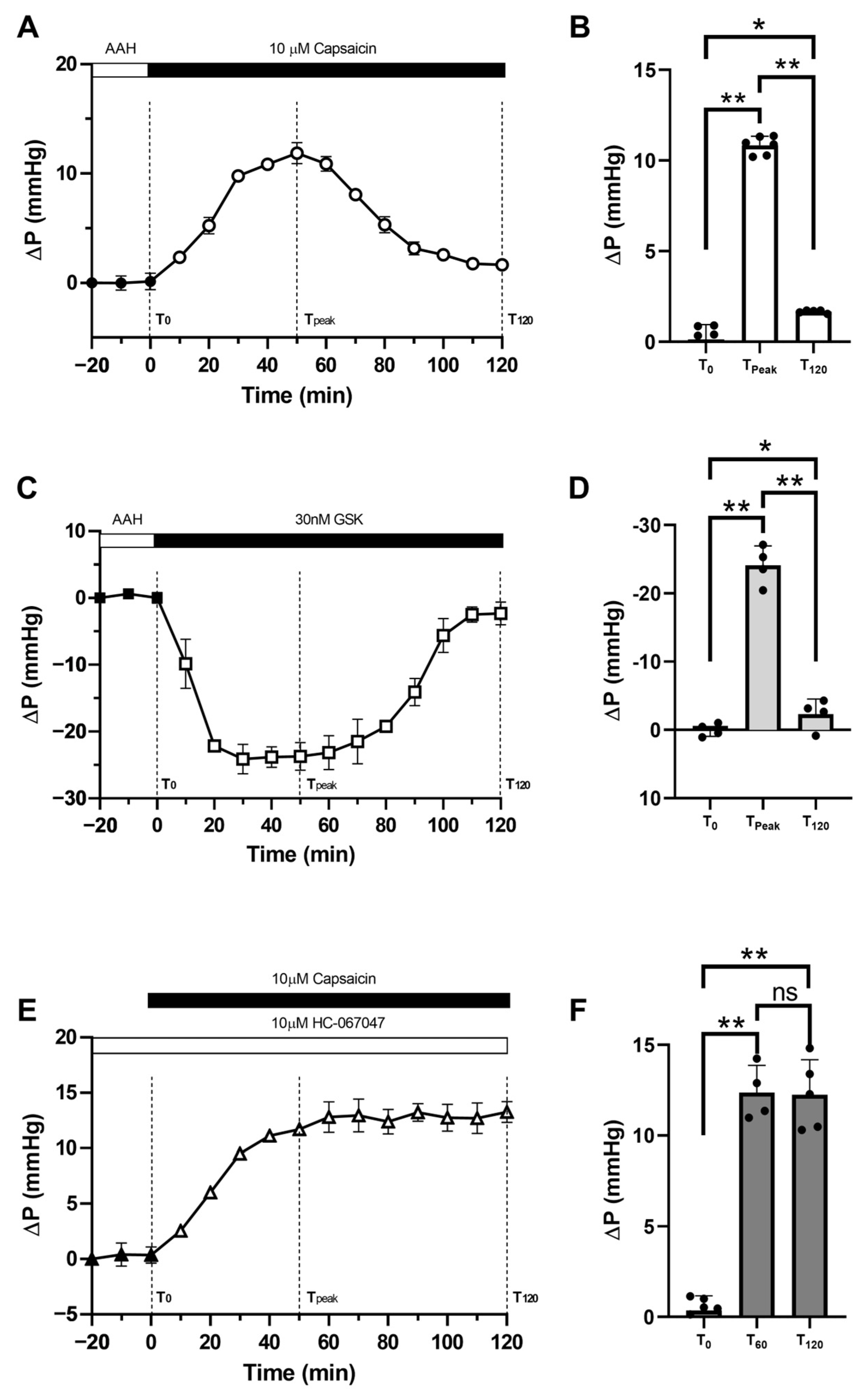
2.1. Pilocarpine Increases Hydrostatic Pressure in the Rat Lens
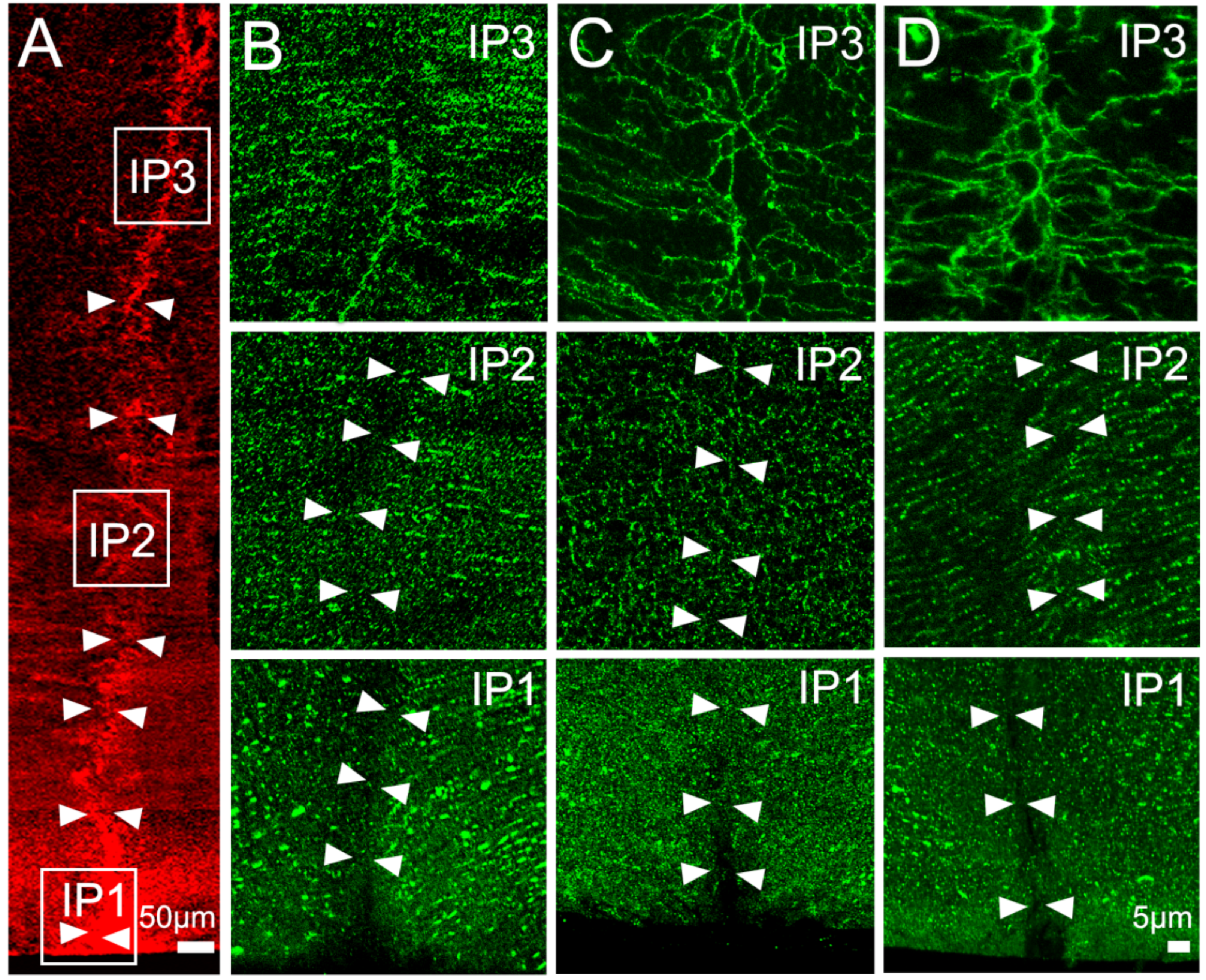
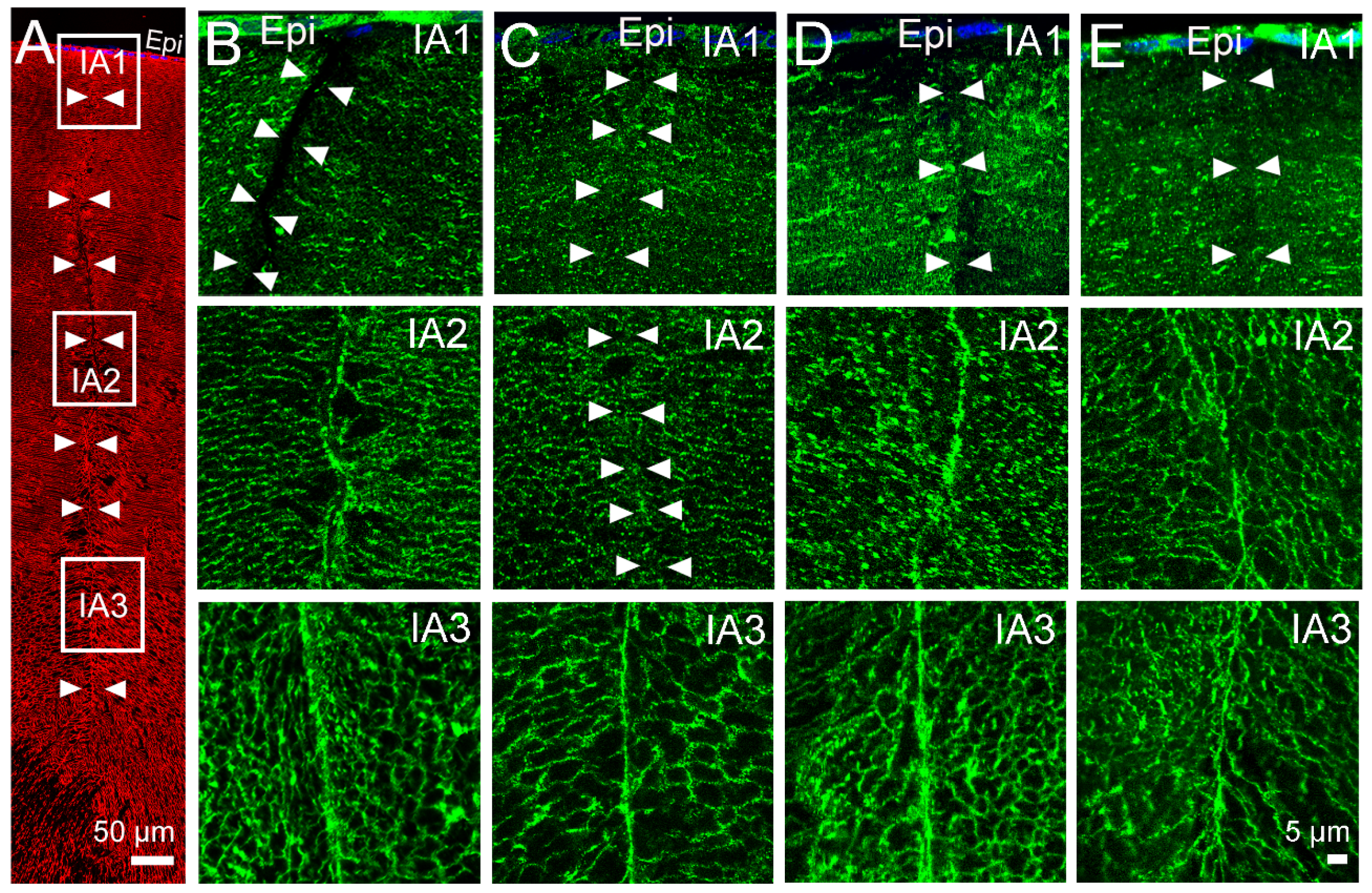
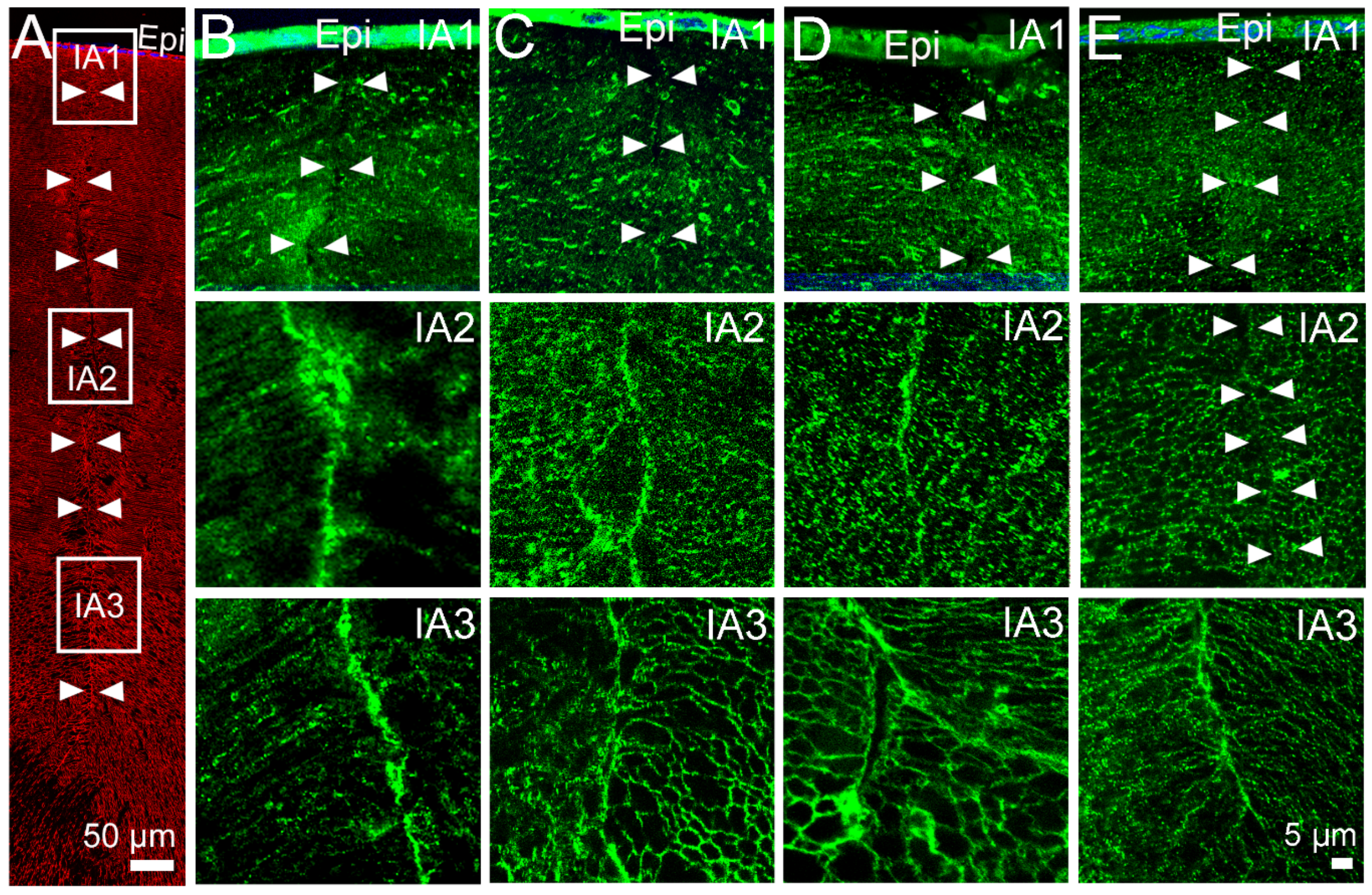
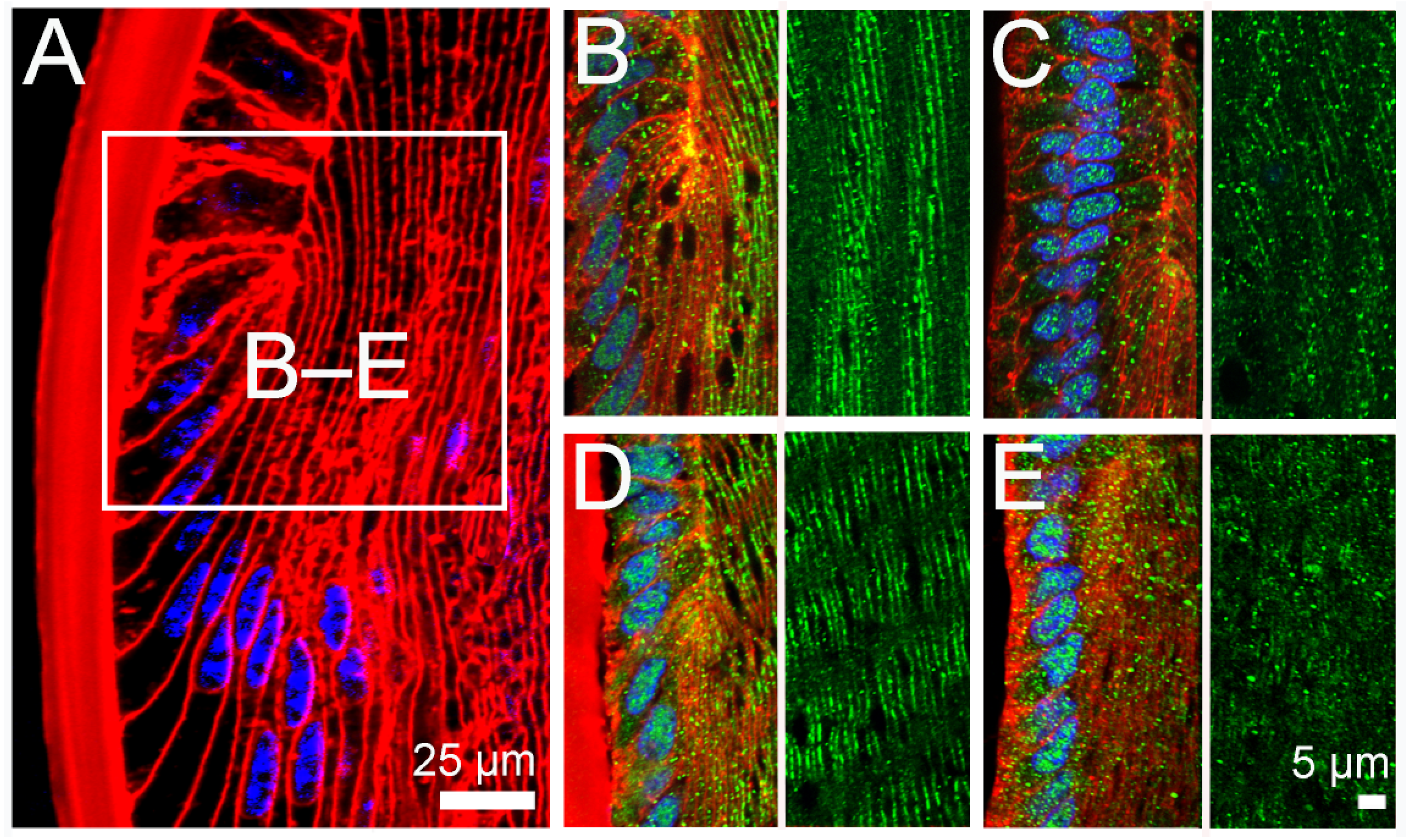
2.2. Effect of Inhibition of TRPV1 on Membrane Trafficking AQP5 in the Anterior and Posterior Influx Zones
2.3. Effect of Direct Activation of TRPV1 on Surface Pressure and Subcellular Localisation of AQP5 in the Rat Lens
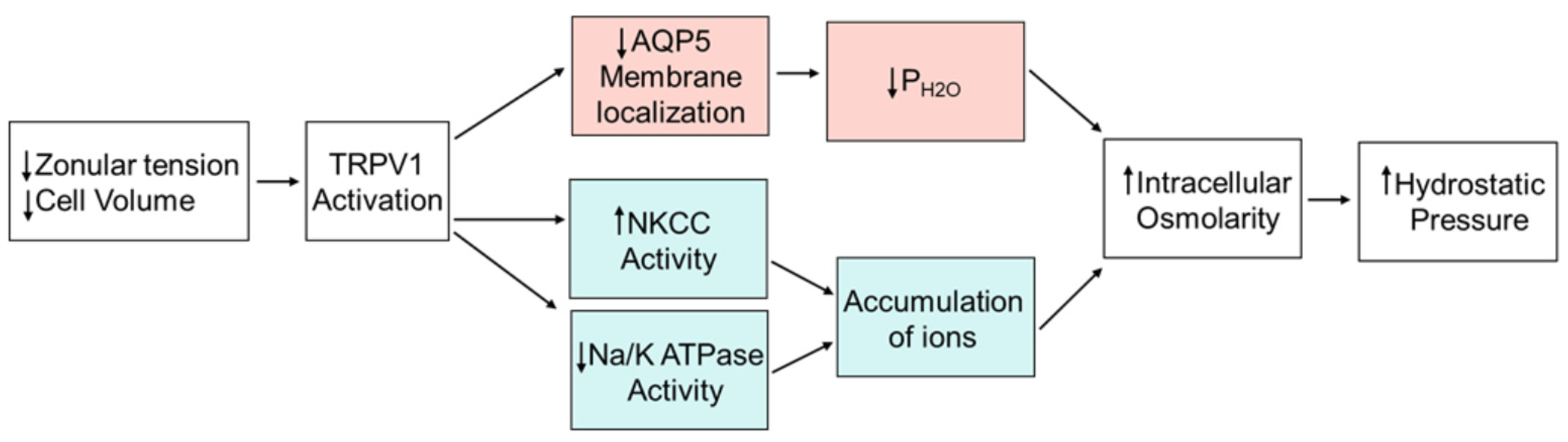
3. Discussion
4. Methods and Materials
4.1. Reagents and Buffers
4.2. Lens Preparation
4.3. Measurements of Lens Intracellular Hydrostatic Pressure
4.4. Lens Organ Culture, Fixation, and Immunolabelling
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Donaldson, P.J.; Chen, Y.; Petrova, R.S.; Grey, A.C.; Lim, J.C. Regulation of lens water content: Effects on the physiological optics of the lens. Prog. Retin. Eye Res. 2022, 101152, in press. [Google Scholar] [CrossRef]
- Donaldson, P.J.; Grey, A.C.; Maceo Heilman, B.; Lim, J.C.; Vaghefi, E. The physiological optics of the lens. Prog. Retin. Eye Res. 2017, 56, e1–e24. [Google Scholar] [CrossRef]
- Audette, D.S.; Scheiblin, D.A.; Duncan, M.K. The molecular mechanisms underlying lens fiber elongation. Exp. Eye Res. 2017, 156, 41–49. [Google Scholar] [CrossRef]
- Bassnett, S. On the mechanism of organelle degradation in the vertebrate lens. Exp. Eye Res. 2009, 88, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Bassnett, S.; Shi, Y.; Vrensen, G.F. Biological glass: Structural determinants of eye lens transparency. Philos. Trans. R Soc. Lond. B Biol. Sci. 2011, 366, 1250–1264. [Google Scholar] [CrossRef] [PubMed]
- Kuszak, J.R.; Zoltoski, R.K.; Tiedemann, C.E. Development of lens sutures. Int. J. Dev. Biol. 2004, 48, 889–902. [Google Scholar] [CrossRef]
- McAvoy, J.W.; Chamberlain, C.G.; de Iongh, R.U.; Hales, A.M.; Lovicu, F.J. Lens development. Eye 1999, 13, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, Y.; Lovicu, F.J.; McAvoy, J.W. Planar cell polarity in the mammalian eye lens. Organogenesis 2011, 7, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, P.J.; Chee, K.S.; Lim, J.C.; Webb, K.F. Regulation of lens volume: Implications for lens transparency. Exp. Eye Res. 2009, 88, 144–150. [Google Scholar] [CrossRef]
- Vaghefi, E.; Kim, A.; Donaldson, P.J. Active Maintenance of the Gradient of Refractive Index Is Required to Sustain the Optical Properties of the Lens. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7195–7208. [Google Scholar] [CrossRef]
- Vaghefi, E.; Donaldson, P.J. The lens internal microcirculation system delivers solutes to the lens core faster than would be predicted by passive diffusion. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R994–R1002. [Google Scholar] [CrossRef]
- Mathias, R.T.; White, T.W.; Gong, X. Lens gap junctions in growth, differentiation, and homeostasis. Physiol. Rev. 2010, 90, 179–206. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, M.D.; Soeller, C.; Sisley, A.M.; Cannell, M.B.; Donaldson, P.J. Gap junction processing and redistribution revealed by quantitative optical measurements of connexin46 epitopes in the lens. Investig. Ophthalmol. Vis. Sci. 2004, 45, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Baldo, G.J.; Mathias, R.T. Spatial variations in membrane properties in the intact rat lens. Biophys. J. 1992, 63, 518–529. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Sun, X.; Yatsula, V.; Wymore, R.S.; Mathias, R.T. Isoform-specific function and distribution of Na/K pumps in the frog lens epithelium. J. Membr. Biol. 2000, 178, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Muir, E.R.; Pan, X.; Donaldson, P.J.; Vaghefi, E.; Jiang, Z.; Sellitto, C.; White, T.W. Multi-parametric MRI of the physiology and optics of the in-vivo mouse lens. Magn. Reson. Imaging 2020, 70, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Barton, K.; De Maria, A.; Petrash, J.M.; Shiels, A.; Bassnett, S. The stratified syncytium of the vertebrate lens. J. Cell Sci. 2009, 122, 1607–1615. [Google Scholar] [CrossRef]
- Gao, J.; Sun, X.; Moore, L.C.; White, T.W.; Brink, P.R.; Mathias, R.T. Lens intracellular hydrostatic pressure is generated by the circulation of sodium and modulated by gap junction coupling. J. Gen. Physiol. 2011, 137, 507–520. [Google Scholar] [CrossRef]
- Gao, J.; Sun, X.; Moore, L.C.; Brink, P.R.; White, T.W.; Mathias, R.T. The effect of size and species on lens intracellular hydrostatic pressure. Investig. Ophthalmol. Vis. Sci. 2013, 54, 183–192. [Google Scholar] [CrossRef]
- Gao, J.; Sun, X.; White, T.W.; Delamere, N.A.; Mathias, R.T. Feedback Regulation of Intracellular Hydrostatic Pressure in Surface Cells of the Lens. Biophys. J. 2015, 109, 1830–1839. [Google Scholar] [CrossRef]
- Shahidullah, M.; Mandal, A.; Mathias, R.T.; Gao, J.; Križaj, D.; Redmon, S.; Delamere, N.A. TRPV1 activation stimulates NKCC1 and increases hydrostatic pressure in the mouse lens. Am. J. Physiol. Cell Physiol. 2020, 318, C969–C980. [Google Scholar] [CrossRef]
- Chen, Y.; Petrova, R.S.; Qiu, C.; Donaldson, P.J. Intracellular hydrostatic pressure regulation in the bovine lens: A role in the regulation of lens optics? Am. J. Physiol. 2022, 322, R263–R279. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gao, J.; Li, L.; Sellitto, C.; Mathias, R.T.; Donaldson, P.J.; White, T.W. The Ciliary Muscle and Zonules of Zinn Modulate Lens Intracellular Hydrostatic Pressure Through Transient Receptor Potential Vanilloid Channels. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4416–4424. [Google Scholar] [CrossRef] [PubMed]
- Verkman, A.S. Water Permeability Measurement in Living Cells and Complex Tissues. J. Membr. Biol. 2000, 173, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Schey, K.L.; Gletten, R.B.; O’Neale, C.V.T.; Wang, Z.; Petrova, R.S.; Donaldson, P.J. Lens Aquaporins in Health and Disease: Location is Everything! Front. Physiol. 2022, 13, 882550. [Google Scholar] [CrossRef]
- Schey, K.L.; Petrova, R.S.; Gletten, R.B.; Donaldson, P.J. The Role of Aquaporins in Ocular Lens Homeostasis. Int. J. Mol. Sci. 2017, 18, 2693. [Google Scholar] [CrossRef]
- Kumari, S.S.; Varadaraj, K. Intact AQP0 performs cell-to-cell adhesion. Biochem. Biophys. Res. Commun. 2009, 390, 1034–1039. [Google Scholar] [CrossRef]
- Grey, A.C.; Walker, K.L.; Petrova, R.S.; Han, J.; Wilmarth, P.A.; David, L.L.; Donaldson, P.J.; Schey, K.L. Verification and spatial localization of aquaporin-5 in the ocular lens. Exp. Eye Res. 2013, 108, 94–102. [Google Scholar] [CrossRef]
- Petrova, R.S.; Schey, K.L.; Donaldson, P.J.; Grey, A.C. Spatial distributions of AQP5 and AQP0 in embryonic and postnatal mouse lens development. Exp. Eye Res. 2015, 132, 124–135. [Google Scholar] [CrossRef]
- D’Agostino, C.; Parisis, D.; Chivasso, C.; Hajiabbas, M.; Soyfoo, M.S.; Delporte, C. Aquaporin-5 Dynamic Regulation. Int. J. Mol. Sci. 2023, 24, 1889. [Google Scholar] [CrossRef]
- Petrova, R.S.; Webb, K.F.; Vaghefi, E.; Walker, K.; Schey, K.L.; Donaldson, P.J. Dynamic functional contribution of the water channel AQP5 to the water permeability of peripheral lens fiber cells. Am. J. Physiol. Cell Physiol. 2018, 314, C191–C201. [Google Scholar] [CrossRef]
- Jacobs, M.D.; Donaldson, P.J.; Cannell, M.B.; Soeller, C. Resolving morphology and antibody labeling over large distances in tissue sections. Microsc. Res. Tech. 2003, 62, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Delamere, N.A.; Shahidullah, M.; Mathias, R.T.; Gao, J.; Sun, X.; Sellitto, C.; White, T.W. Signaling Between TRPV1/TRPV4 and Intracellular Hydrostatic Pressure in the Mouse Lens. Investig. Ophthalmol. Vis. Sci. 2020, 61, 58. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.E.; Mathias, R.T. The aquaporin zero puzzle. Biophys. J. 2014, 107, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Froger, A.; Clemens, D.; Kalman, K.; Németh-Cahalan, K.L.; Schilling, T.F.; Hall, J.E. Two distinct aquaporin 0s required for development and transparency of the zebrafish lens. Investig. Ophthalmol. Vis. Sci. 2010, 51, 6582–6592. [Google Scholar] [CrossRef] [PubMed]
- Kalman, K.; Németh-Cahalan, K.L.; Froger, A.; Hall, J.E. Phosphorylation determines the calmodulin-mediated Ca2+ response and water permeability of AQP0. J. Biol. Chem. 2008, 283, 21278–21283. [Google Scholar] [CrossRef] [PubMed]
- Mathias, R.T.; Wang, H. Local osmosis and isotonic transport. J. Membr. Biol. 2005, 208, 39–53. [Google Scholar] [CrossRef]
- Pierscionek, B.K. In vitro alteration of human lens curvatures by radial stretching. Exp. Eye Res. 1993, 57, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Pierscionek, B.K. Age-related response of human lenses to stretching forces. Exp. Eye Res. 1995, 60, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Fu, T.; Xu, Q.; Chen, W.; Chen, Z.; Guo, J.; Deng, C.; Manyande, A.; Wang, P.; Zhang, H. Quantitative analysis of internal components of the human crystalline lens during accommodation in adults. Sci. Rep. 2021, 11, 6688. [Google Scholar] [CrossRef]
- Petrova, R.S.; Bavana, N.; Zhao, R.; Schey, K.L.; Donaldson, P.J. Changes to Zonular Tension Alters the Subcellular Distribution of AQP5 in Regions of Influx and Efflux of Water in the Rat Lens. Investig. Ophthalmol. Vis. Sci. 2020, 61, 36. [Google Scholar] [CrossRef]
- Ishikawa, H.; DeSantis, L.; Patil, P.N. Selectivity of muscarinic agonists including (+/−)-aceclidine and antimuscarinics on the human intraocular muscles. J. Ocul. Pharmacol. Ther. 1998, 14, 363–373. [Google Scholar] [CrossRef]
- Shahidullah, M.; Mandal, A.; Beimgraben, C.; Delamere, N.A. Hyposmotic stress causes ATP release and stimulates Na,K-ATPase activity in porcine lens. J. Cell. Physiol. 2012, 227, 1428–1437. [Google Scholar] [CrossRef] [PubMed]
- Willette, R.N.; Bao, W.; Nerurkar, S.; Yue, T.-l.; Doe, C.P.; Stankus, G.; Turner, G.H.; Ju, H.; Thomas, H.; Fishman, C.E.; et al. Systemic Activation of the Transient Receptor Potential Vanilloid Subtype 4 Channel Causes Endothelial Failure and Circulatory Collapse: Part 2. J. Pharmacol. Exp. Ther. 2008, 326, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Julius, D. The Vanilloid Receptor: A Molecular Gateway to the Pain Pathway. Annu. Rev. Neurosci. 2001, 24, 487–517. [Google Scholar] [CrossRef]
- Holzer, P. Capsaicin: Cellular targets, mechanisms of action, and selectivity for thin sensory neurons. Pharmacol. Rev. 1991, 43, 143–201. [Google Scholar] [PubMed]
- Shahidullah, M.; Mandal, A.; Delamere, N.A. Damage to lens fiber cells causes TRPV4-dependent Src family kinase activation in the epithelium. Exp. Eye Res. 2015, 140, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.S.; Keddy, R.; Zheng, G.Z.; Schmidt, R.G.; Koenig, J.R.; McDonald, H.A.; Bianchi, B.R.; Honore, P.; Jarvis, M.F.; Surowy, C.S.; et al. Tetrahydropyridine-4-carboxamides as novel, potent transient receptor potential vanilloid 1 (TRPV1) antagonists. Bioorganic Med. Chem. 2008, 16, 8516–8525. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrova, R.S.; Nair, N.; Bavana, N.; Chen, Y.; Schey, K.L.; Donaldson, P.J. Modulation of Membrane Trafficking of AQP5 in the Lens in Response to Changes in Zonular Tension Is Mediated by the Mechanosensitive Channel TRPV1. Int. J. Mol. Sci. 2023, 24, 9080. https://doi.org/10.3390/ijms24109080
Petrova RS, Nair N, Bavana N, Chen Y, Schey KL, Donaldson PJ. Modulation of Membrane Trafficking of AQP5 in the Lens in Response to Changes in Zonular Tension Is Mediated by the Mechanosensitive Channel TRPV1. International Journal of Molecular Sciences. 2023; 24(10):9080. https://doi.org/10.3390/ijms24109080
Chicago/Turabian StylePetrova, Rosica S., Nikhil Nair, Nandini Bavana, Yadi Chen, Kevin L. Schey, and Paul J. Donaldson. 2023. "Modulation of Membrane Trafficking of AQP5 in the Lens in Response to Changes in Zonular Tension Is Mediated by the Mechanosensitive Channel TRPV1" International Journal of Molecular Sciences 24, no. 10: 9080. https://doi.org/10.3390/ijms24109080